
干旱胁迫对黄土高原4种蒿属植物叶形态解剖学特征的影响
2014-08-08 02:15:09梁宗锁龚春梅韩蕊莲
生态学报 2014年16期
王 勇,梁宗锁,龚春梅,韩蕊莲,于 靖
(1. 中国科学院, 教育部, 水土保持与生态环境研究中心,杨凌 712100; 2. 西北农林科技大学, 生命科学学院,杨凌 712100)
干旱胁迫对黄土高原4种蒿属植物叶形态解剖学特征的影响
王 勇1,2,梁宗锁1,2,龚春梅2,韩蕊莲1,*,于 靖2
(1. 中国科学院, 教育部, 水土保持与生态环境研究中心,杨凌 712100; 2. 西北农林科技大学, 生命科学学院,杨凌 712100)
以黄土高原演替初期阶段的猪毛蒿(ArtemisiascopariaWaldst. et Kit.)、茭蒿(ArtemisiagiraldiiPamp.)、铁杆蒿(ArtemisiagmeliniiWeb. ex Stechm.)3种优势种和1种常见种黄花蒿(ArtemisiaannuaLinn.)为供试材料,采用盆栽试验与称重控水法,将土壤含水量分别控制在田间最大持水量的75%(适宜水分对照)、55%(中度干旱)和35%(重度干旱),研究干旱胁迫对菊科蒿属植物叶片形态解剖学的影响,揭示植物叶片生态适应机制,为生态恢复过程中植被演替的规律提供理论依据。于植物营养生长末期选取代表性的叶片,分别采用电子扫描显微镜、电子透射显微镜和光学显微镜对叶表皮特征、亚细胞显微特征和组织结构进行观察。结果表明:干旱胁迫下,这4种植物表现为不同程度的叶片增厚、栅栏组织厚度、叶片紧实度和角质层厚度增加;3种旱生型植物猪毛蒿、茭蒿和铁杆蒿的叶片下表皮具有浓密的表皮毛,有利于防止水分过度蒸腾,而中生型黄花蒿叶片无此功能;铁杆蒿和黄花蒿上表皮毛稀少,不利于保水和防止强光照,干旱胁迫下依靠降低上表皮气孔密度和叶细胞叶绿体数目来适应环境;茭蒿和猪毛蒿叶绿体形态相似,具有较厚的基粒和浓密的基粒片层,强光下容易引起氧化损伤,猪毛蒿通过浓密的上表皮毛而茭蒿通过较强的抗氧化机制防止光能过剩,两者在重度干旱下均产生大量脂质体。4种蒿属植物中黄花蒿的叶片解剖学特征受到干旱的影响变化最大,铁杆蒿和猪毛蒿次之,茭蒿最弱,3种优势种的干旱适应性显著强于黄花蒿,干旱下叶片解剖学的结论与干旱耐受性指标结果一致。
菊科蒿属植物;气孔特征;组织解剖;抗旱性
黄土高原地区是典型的干旱、半干旱地区,也是我国水土流失最为严重的地区。陕北黄土丘陵区,隶属于温带大陆性季风气候,年均气温6—14℃,年均降水量为200—700,形成了该地区特有的干燥、温差大的生态环境[1]。由于人类活动造成黄土高原生态系统退化和水土流失,在黄土高原地区出现了旱化为主要特征的土壤退化现象,制约着植被恢复进程。然而在这种干旱的环境下,菊科蒿属植物以其自身独特的生理生态特征作为先锋物种,出现在黄土高原撂荒地次生演替的各个阶段[2- 4]。作为该区域自然演替序列早期的优势建群物种,猪毛蒿(ArtemisiascopariaWaldst. et Kit.)、铁杆蒿(ArtemisiagmeliniiWeb. ex Stechm.)、茭蒿(ArtemisiagiraldiiPamp.)等菊科蒿属植物适应自然植被演替序列发展前期干旱环境,对土壤养分及微环境的改造产生了重要的影响[4- 8]。因此,研究这些先锋物种与干旱的关系对于揭示植物干旱适应性及黄土高原植被恢复有重要意义。
目前众多研究表明:干旱胁迫下,植物叶片气孔密度增加,气孔阻力上升,气孔开度变小,气孔趋于关闭,气孔的长、宽明显减小,光合速率下降,并且明显地降低了叶片细胞的叶绿体数目及每个叶绿体内的淀粉粒数[9- 11];在严重干旱下,抗旱性弱的小麦品种比抗旱性强的小麦品种更容易受影响[12],比如细胞破裂,内溶物溢出,多数叶绿体溶解散乱在基质中;在对酿酒葡萄、柑橘、苹果和番茄的干旱研究中也发现类似的现象[9,13- 15]。干旱胁迫还对植物叶片组织解剖结构产生影响。随着水分减少植物栅栏组织厚度增加,海绵组织厚度相应减少,叶片厚度增大,叶片组织结构紧密度增加,疏松度减少[16- 20]。此外,植物为了适应干旱、强辐射环境,通常表现为叶片小而厚、叶表皮角质层增厚、叶肉栅栏组织/海绵组织增加、叶脉机械组织发达、进化出表皮附属物、叶片还特化成鳞片状、条状、柱状或针状[21- 28]。关于陕北撂荒地菊科蒿属植物的研究已有许多报道,但多集中在群落演替、种间竞争、水分平衡、形态解剖适应性、抗氧化特性等研究方面[29- 34],而对于该地区演替优势种叶片形态解剖学与水分关系的研究还较少。因此,本实验选取陕北撂荒地演替初期阶段的3种优势种:猪毛蒿(ArtemisiascopariaWaldst. et Kit.)、茭蒿(ArtemisiagiraldiiPamp.)和铁杆蒿(ArtemisiagmeliniiWeb. ex Stechm.))及1种黄土高原常见种黄花蒿(ArtemisiaannuaLinn.)作为研究材料,观察其不同土壤干旱条件下的叶表皮特征、亚细胞显微特征和组织结构的变化,进而从形态解剖学方面阐明其适应土壤干旱的生理机制,为黄土高原干旱半干旱条件下的生态恢复提供理论依据。
1 试验材料与方法
1.1 试验材料
猪毛蒿、铁杆蒿、茭蒿和黄花蒿的种子于2009年11月份采自陕北黄土丘陵沟壑区安塞县高桥乡的自然植物群落中,室内自然风干后,选取籽粒饱满的种子密封置于室外自然过冬,2010年4月初播种于中国科学院水土保持研究所干旱防雨棚内。
1.2 试验设计
盆栽试验所用塑料桶规格为22 cm(高)×24 cm(内径),盆栽土壤为关中土娄土,田间最大持水量为28%。试验设适宜水分(对照),中度、重度干旱胁迫3个水分处理,其土壤含水量分别占田间最大持水量的75%(75%θ∫),55%(55%θ∫),35%(35%θ∫),每处理4个重复,同时设立3个水分梯度的空白盆栽桶(未播种)。所有盆栽桶内土壤干重一致,均为14.5kg,水分充分饱和,播种发芽后,全部浇水至适宜水分,待所有苗子长到三叶期时,每桶留取生长健壮一致幼苗8株。从5月份下旬开始选择性浇水,使各个处理的土壤含水量达设定的水平,之后每隔2 d用电子秤(精度0.005kg)称量并记录各桶质量1次,再补充水分至各个处理水平的土壤水分含量。植物在3个水分梯度下经过3个月的营养生长后分别选取有代表性的功能叶进行解剖学研究和生物量测定计算。
1.3 研究方法
选取不同干旱处理的4种蒿属盆栽植物中部成熟叶片作为研究材料,每种植物每个处理选取10株共20片叶片,分为两组,一组切取1mm×4mm方片用于制备半薄和超薄切片,另一组切取5mm×5 mm方片用于制备电子扫描样品,所有叶片切取部位均位于叶片中部。将两组切取好的样品立即投入预冷的4%戊二醛溶液中,4℃下固定4h;之后用0.1mol/L的PBS漂洗3次,10min/次;再将样品投入1%锇酸溶液中,固定2h。取出后,用0.1mol/L的PBS漂洗3次,10min/次;再将材料依次放入50%乙醇、60%乙醇、70%乙醇、80%乙醇、90%乙醇、100%乙醇各1次,20min/次,纯丙酮2次,30min/次。
第一组用于制备半薄和超薄切片的样品在室温下,依次放入1∶3、1∶1、3∶1的环氧树脂Epon812包埋剂丙酮溶液中,每级4h,最后纯包埋剂过夜。再在包埋模具内滴1滴包埋剂,把样品置入并使之定位于合适位置,缓缓注入包埋剂,然后放入烤箱中进行聚合。控温时间及温度依次为:37℃12h;45℃12h;60℃24h。之后修块儿、切片半薄切片(0.5—1m),甲苯胺蓝染色用于光学显微镜观察组织结构;超薄切片(50—70nm),醋酸铀染色10—15min,用于电子透射显微观察亚细胞结构。
1.4 观察及测定项目
1.4.1 蒸腾耗水量、干物质生物量和水分利用效率
整个生长季植物蒸腾耗水量为45次(3个月)称重控水计算所得蒸腾耗水量之和减去空白土壤盆栽水分总蒸散量,每次各个盆栽蒸腾耗水量则为各自设定质量减去浇水前称重质量,空白土壤蒸散量同上。为了排除植物生长带来的水分补充误差,每隔15d拔取植株3棵称量鲜重,换算每盆增加质量,然后调整设定盆栽质量。处理结束后小心取出完整植株,90℃杀青,60℃烘干至恒重。水分利用效率= 干物质量(g)/ 生长季耗水量(kg)。
1.4.2 电子扫描显微镜(日本电子,JSM- 6360LV,日本)下观察4种蒿属植物叶片上下表皮及其附属物的形态观察,电子透射显微镜(日立, HT7700, 日本)观察亚细胞结构叶绿体的形态变化。
1.4.3 光学显微镜(OLYMPUS BX51+DP70, 日本)下观察测定4种蒿属植物在3种土壤水分条件下叶片上下表皮的气孔密度(个/mm2)、表皮细胞密度(个/mm2)、气孔长宽,计算气孔指数。
气孔指数=100×气孔密度/(气孔密度+表皮细胞密度)。
1.4.4 光学显微镜(OLYMPUS BX51+DP70, 日本)下观察测定4种蒿属植物在3种土壤水分条件下叶片组织解剖学特征,包括叶片厚度、表皮厚度、角质层厚度、栅栏组织厚度、海绵组织厚度。
叶片结构紧密度=100%×栅栏组织厚度 / 叶片厚度。
叶片结构疏松度=100%×海绵组织厚度 / 叶片厚度。
1.5 数据处理
用Excel 2003整理试验数据,SPASS 17.0软件进行统计分析,字母法标记显著水平。
2 结果与分析
2.1 蒸腾耗水量、干物质生物量和水分利用效率
由图1可见,中度和重度水分胁迫条件下,4种蒿属植物的耗水量相对于适宜水分条件均减少,减少的幅度分别为:铁杆蒿,20.3%和58.3%;猪毛蒿,26.3%和54.5%;茭蒿,24.7%和68.4%;黄花蒿,17.8%和44.5%。总生物量均在水分胁迫下降低(猪毛蒿在中度水分胁迫下例外)。与重度水分胁迫相比,中度水分胁迫下, 4个种总生物量下降的幅度:茭蒿(0.4%)<铁杆蒿(32.4%)和黄花蒿(32.8%);严重干旱下4个种总生物量下降幅度为:猪毛蒿(49.6%)<茭蒿(53.5%)<铁杆蒿(68.2%)和黄花蒿(67.8%)。在中度和重度水分胁迫条件下,4种蒿属植物的水分利用效率相对于适宜水分条件变化率表现为:铁杆蒿,-13.6%和-20.4%;猪毛蒿,38.3%和9.6%;茭蒿,30.9%和51.5%;黄花蒿,-20.0%和-46.9%。

图1 3种土壤水分条件下4种蒿属植物生长季内蒸腾耗水量、干物质量和水分利用效率变化Fig.1 Effect of different soil water content on total water consumption, vegetative dry weight and WUE of 4 Artemisia
2.2 4种蒿属植物的叶片表面特征
2.2.1 4种蒿属植物表皮附属物
由图2 可见,铁杆蒿和茭蒿的上表皮表皮毛呈卷曲状,下表皮的表皮毛较浓密,这可以有效地防止水分蒸腾。而猪毛蒿的上下表皮都布满了横向平行排列的表皮毛,其表皮毛由表皮细胞向上突出后分成两根向两侧弯曲平行排列。而黄花蒿的上下表皮都只零星分布极个别的表皮毛。黄花蒿和铁杆蒿上下表皮也都有囊泡状的附属物。

图2 4种蒿属植物表皮及其附属物Fig.2 Leaf epidermis and attachment of 4 Artemisia species1: 铁杆蒿上表皮Upper epidermis of A.gmelinii;2: 铁杆蒿下表皮Lower epidermis of A.gmelinii;3: 猪毛蒿上表皮Upper epidermis of A.scoparia;4: 猪毛蒿下表皮Lower epidermis of A.scoparia;5: 茭蒿上表皮Upper epidermis of A.giraldii;6: 茭蒿下表皮Lower epidermis of A.giraldii;7: 黄花蒿上表皮Upper epidermis of A.annua;8: 黄花蒿下表皮Lower epidermis of A.annua.
2.2.2 3种土壤水分条件下4种蒿属植物叶气孔特征比较
由图3所示,气孔密度受干旱影响最大的是黄花蒿,与适宜水分条件下的相比,在重度干旱条件下黄花蒿上表皮气孔密度增加近20倍,下表皮则增加了近0.7倍;猪毛蒿的上下气孔密度分别在重度干旱是对照的近1.6倍和1.5倍; 铁杆蒿则增加近1.5倍左右。较特殊的是铁杆蒿上表皮气孔密度在干旱下呈现下降趋势,同时茭蒿的上、下表皮气孔密度表现出相反的变化。
4种植物的表皮密度受干旱影响表现出不同的变化。下表皮密度在干旱下表现为:茭蒿降低,其余3种植物重度干旱下增加。铁杆蒿和猪毛蒿上表皮密度均在中度干旱下降低,重度干旱下与对照无显著性差异;黄花蒿只有在重度干旱下有所下降;例外的是,茭蒿的上表皮密度在干旱下呈上升趋势。
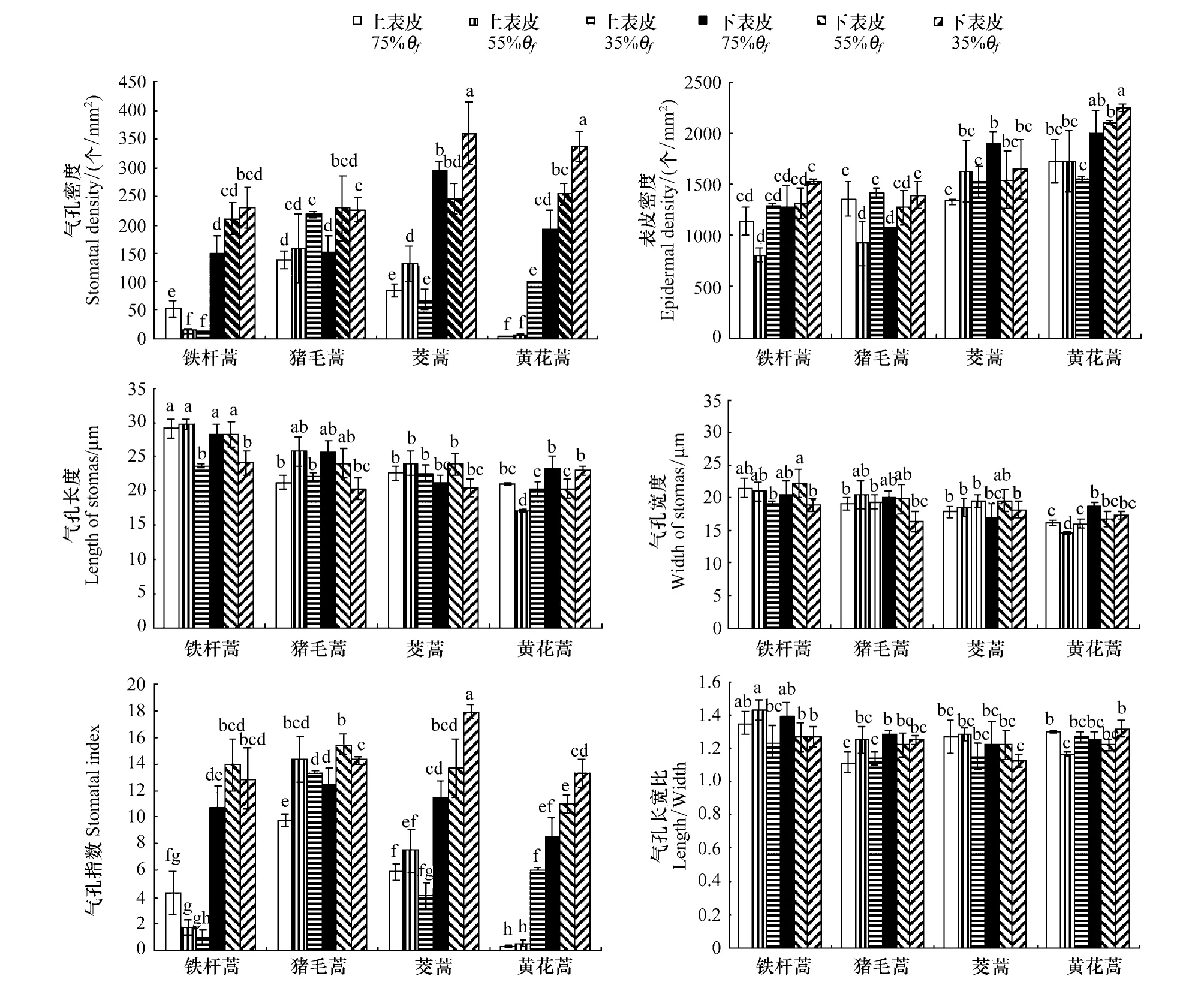
图3 3种不同土壤水分条件下4种蒿属植物叶表皮细胞和气孔器的比较Fig.3 Comparison of epedermal cells and stomas features of leaves of 4 Artemisia species under 3 different soil water content小写字母a—g表示个处理间有无显著差异(P<0.05),字母相同表示差异性不显著; 图中数据:均值±SE,n=20
干旱对气孔指数的影响在铁杆蒿和黄花蒿的上表皮表现最为显著。与适宜水分条件下相比,重度干旱下铁杆蒿上表皮气孔指数下降近4.3倍,黄花蒿增加近20倍;猪毛蒿在中度和重度干旱下均增加近1.4倍;茭蒿仅在中度干旱下增加,重度干旱下有所下降;重度干旱下黄花蒿和茭蒿下表皮气孔指数在增加近1.6倍,铁杆蒿和猪毛蒿有所增加,但差异不显著。
铁杆蒿的上下表皮气孔长度均在重度干旱下显著下降;猪毛蒿上表皮气孔长度却在中度水分条件下显著上升,下表皮呈下降趋势;与之相反,黄花蒿的上表皮气孔长度在中度下显著下降。黄花蒿的上表皮气孔宽度在中度水分条件下显著下降;铁杆蒿和黄花蒿的上表皮气孔的长宽比分别在中度和中度干旱下显著下降。
2.3 3种土壤水分条件下4种蒿属植物叶片亚细胞结构
铁杆蒿的叶绿体贴紧细胞壁周围一圈,并且有较大的淀粉颗粒,充满叶绿体大部分位置,有的叶绿体甚至多达3个淀粉粒(图版1,1—9);茭蒿叶肉细胞内的叶绿体也有较多的淀粉粒(图版1,22;图版2,25);而猪毛蒿和黄花蒿叶绿体的淀粉粒较少或者较小(图版1,10,13—16;图版2,28、32、33)。铁杆蒿和黄花蒿的叶绿体较大,在每个细胞内分布的个数较少(图版1,4;图版2,34),而猪毛蒿和茭蒿正好相反,其叶绿体较小,但每个细胞分布的数目较多(图版1,13、16;图版2,25)。铁杆蒿在适宜水分条件下叶绿体内基粒分布稀疏,但成规则的形状(图版1,2、3);中度干旱下基粒数目增多,但基质片层增厚(图版1,5、6);在重度干旱下叶绿体内基粒密度增加,但基质片层杂乱(图版1,8)。铁杆蒿在3种水分处理下均可见叶绿体内相当数量的脂质体,其大小不一。猪毛蒿在适宜水分下叶绿体内基粒较厚,基质片层清晰可见,并且排列整体(图版1,11、12);在中度干旱下,与适宜水分下相似(图版1,15),但细胞内叶绿体数目有所增加(图版1,13);在重度干旱下基质片层增粗,基粒数目增多,但排列凌乱,脂质体数目明显增多(图版1,17、18)。茭蒿在干旱下,与适宜水分相比叶绿体内淀粉粒有增多的迹象(图版1,19、22;图版2,25);茭蒿的基粒类囊体和基质片层与其它物种相比非常浓密(图版1,21、23、24),在重度干旱下基粒减少,脂质体增多的现象(图版2,26、27)。黄花蒿的基粒相对于其它物种厚度小一些,基质片层更稀疏,但清晰可见(图版2,30、32、33)。在重度干旱下脂质体增多(图版2,35)。
2.4 4种蒿属植物在3种土壤水分条件下叶片解剖学特征比较
由表1可见,与适宜水分下相比,干旱处理下铁杆蒿和黄花蒿的叶片厚度随着干旱胁迫的加剧而增厚,其增幅分别达13.1%和21.4%(P<0.05);猪毛蒿在中度干旱下达19%,茭蒿变化差异不显著。分别与对照相比较,猪毛蒿在重度干旱下上表皮厚度下降21.9%,中度干旱下下表皮厚度下降16.2%;其余3种植物的上下表皮厚度变化差异不显著。上、下表皮栅栏组织受干旱影响变化与对照相比增幅分别为:铁杆蒿达33.9%—38%(上)和27.7%(下);猪毛蒿在中度干旱下达35.5%(上)和45.6%(下);黄花蒿,30.9%(上)。较特殊的是在重度干旱下茭蒿的上表皮栅栏组织厚度下降20.4%;黄花蒿叶片的下表皮栅栏组织分化出现。叶片海绵组织厚度仅在重度干旱下显著下降,其下降幅度分别为:铁杆蒿22.1%、茭蒿22.4%、黄花蒿26%;猪毛蒿无显著变化。上、下表皮角质层厚度的变化最显著的是猪毛蒿在重度干旱下增幅达38.9%(上)和48%(下);其次是茭蒿在中度干旱下达41.4%(下)和铁杆蒿在重度干旱下达19%(上);特殊的是黄花蒿在重度干旱下上、下表皮角质层厚度分别降低幅度达36%和15.2%。干旱胁迫下,叶片栅栏组织/海绵组织厚度比例与对照相比在铁杆蒿和黄花蒿中增幅达到显著差异,分别高达75.7%和174%;其余变化差异不显著。叶片结构紧实程度上对干旱的响应黄花蒿增幅达69.8%、猪毛蒿18.9%、铁杆蒿10.4%;铁杆蒿和黄花蒿的叶片疏松度在重度干旱下降幅分别达36.7%和38.8%;其余变化无显著差异。
2.5 综合分析
植物的干旱适应性是一个综合的特征,这不仅包括对水分的利用与保持,光合作用的维持,自身生理特征对逆境伤害的抵御,及自身遗传上的可塑性等。利用主成分分析对图2、表1及上下表皮毛(数据录入有表皮毛:1; 无表皮毛:0)有无等各个解剖学指标进行降维(图4),分为两类,CP1主要与植物的保水相关,包括:上表皮角质层、表皮毛、下表皮气孔密度及上下表皮厚度等;CP2主要与植物的光合组织相关,包括:栅栏组织厚度、叶片紧实度等等;累计贡献率达到68%。由图4可以看出铁杆蒿和茭蒿综合表现出对干旱响应较小,而猪毛蒿和黄花蒿的响应较大,或者可塑性大,这或许与铁杆蒿和茭蒿有较强大的根系,而猪毛蒿和黄花蒿根系不发达有关,而根系发达是干旱区植物的主要特征。4种植物在重度干旱下均向植物保水节水发展的趋势,只是猪毛蒿和黄花蒿在干旱条件下表现了较强的适应性变化,具有一定的可塑性,其组织形态、结构和比例的变化体现了对不同干旱强度响应策略的差异。
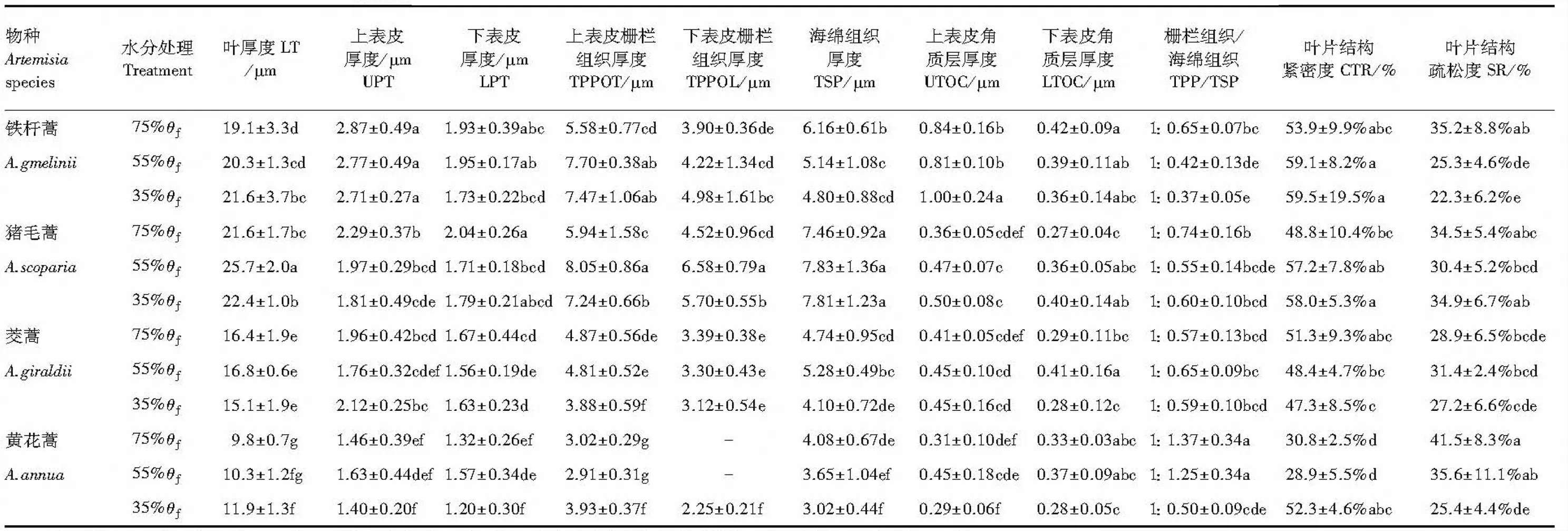
表1 4种蒿属植物在3种土壤水分条件下叶片解剖学特征比较
小写字母a—g表示个处理间有无显著差异(P<0.05),同一列中字母相同表示差异性不显著; 表中数据为:均值±SE,n=20
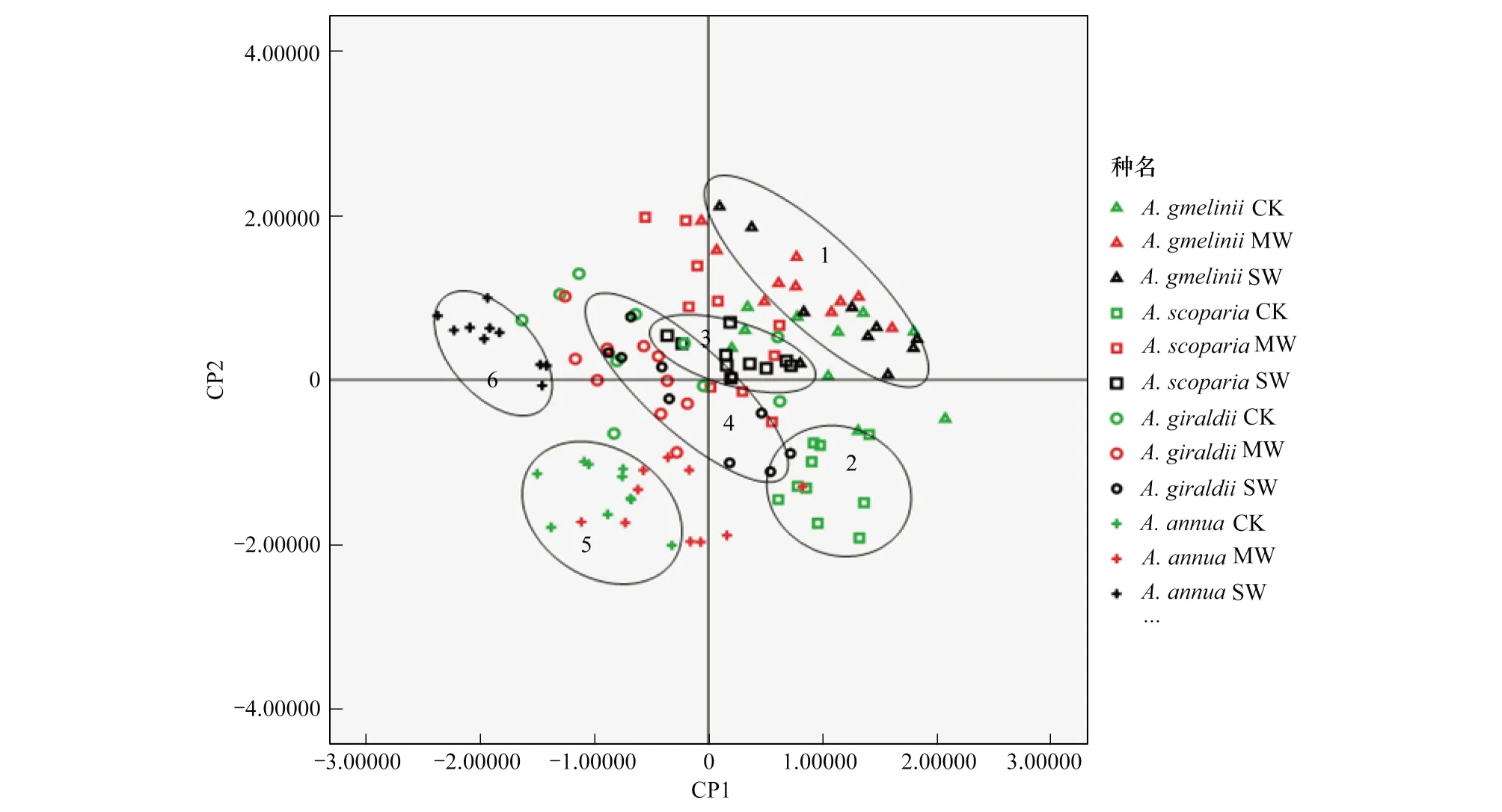
图4 3水分梯度下4种蒿属种叶片形态结构特征主成分分析Fig.4 Principal component analysis of leaf morphological characters of 4 Artemisia species under 3 kinds of water irrigation strategy贡献率:CP1,57.5%; CP2,10.5%, 累积贡献率68%;主要分布示意:1: 铁杆蒿重度干旱;2: 猪毛蒿适宜水分;3: 猪毛蒿重度干旱;4: 茭蒿重度干旱;5: 黄花蒿适宜水分;6: 黄花蒿重度干旱; CK: 适宜水分;MW: 中度干旱;SW: 中度干旱
3 结论与讨论
陕北黄土高原大气湿度低、光辐射强[1],这些因素造成强烈的大气干旱,植物的生存就得首先得保证低的蒸腾作用,再次保证足够的光合作用,并且防止过剩光能的损伤作用。菊科的蒿属植物往往能够凭借自身的生态适应性成为黄土高原的先锋物种,并在该地区的生境改造中扮演着重要的角色[2,4,29,33]。
水分胁迫对植物的影响综合表现在生长及生物量的积累上。耐旱的植物通过长期进化后具有相适应的器官结构及生理生化过程来避免或抵御逆境伤害,最大化的维持自身的生长和繁殖。通过对4种蒿属植物在干旱下的蒸腾耗水量、生物量积累和水分利用效率的测定,表明3种旱生型植物水分调控和利用的能力、维持生长的能力均高于中生型植物;虽然铁杆蒿干旱下的生长和水分利用效率较对照有大幅下降,但其干旱下的数值显示与猪毛蒿和茭蒿的相当,均大于黄花蒿。叶片是植物光合和蒸腾器官,其形态结构直接影响到植物干旱下的水分利用与调整策略,对植物的生长至关重要。因此4种蒿属植物叶片之间的形态结构差异,及其对干旱的响应变化是这些植物对干旱环境重要的适应策略之一,其与根系结构和生理生化过程特征共同决定了植物的适应能力。
表皮毛是植物长期进化适应环境的结果,短期内的水分处理对其影响变化不大,但表皮毛的作用直接或间接的影响植物一系列的生理过程,尤其是蒸腾作用、光合作用;表皮附属物能够反射阳光, 减少叶片表面空气的流动, 降低蒸腾作用, 防止水分过渡丧失[35],进而影响到植物的形态建成。从4种蒿属植物叶表皮形态来看,3种旱生型的下表皮具有浓密的表皮毛,这有利于防止水分过度蒸腾,而中生型黄花蒿就无此功能;铁杆蒿和黄花蒿上表皮毛极其稀少,不利于保水和防止强光照,因此依靠降低上表皮气孔密度和叶细胞叶绿体数目来适应干旱胁迫。本结果进一步补充了杨超等[33]调查了黄土高原撂荒地自然演替群落中3种蒿类具有表皮毛和腺毛等旱生适应结构,揭示了黄花蒿为中生型植物与同类旱生型表皮上的差异。因此从表皮毛生长适应性方面,在防止水分蒸发和强光辐射猪毛蒿最有利,其次茭蒿,再次铁杆蒿,最后黄花蒿,这与干旱下4植物生物量的积累变化相一致。
高度发达的栅栏组织既可避免干旱地区强烈光照对叶肉细胞的灼伤,又可以有效的利用衍射光进行光合作用[16],栅栏组织越厚,栅栏组织与叶厚度的比值越大,栅栏组织细胞越小,且排列越紧密,则植物利用光能的效率越高[17];此外,叶肉栅栏组织发达, 细胞层数增加而体积减小, 海绵组织相对减少, 细胞间隙减小等变化是植物水分短缺响应, 该特征有助于CO2等气体从气孔下室到光合作用场所的传导, 又可抵消因气孔关闭和叶肉结构的变化所引起的CO2传导率的降低, 从而提高植物对水分的利用率, 表现出植物的抗旱适应性[18]。遗传上铁杆蒿属过渡型叶片,猪毛蒿和茭蒿是等面叶,上下表皮均存在紧密的栅栏组织,这均有利于其减少蒸腾,增加光合能力;而黄花蒿是异面叶对干旱的适应能力较差,这也造成其对干旱胁迫的响应变化最大,其叶片在重度干旱下下表皮栅栏组织由无到有,紧实度大大增加,有向等面叶变化的趋势;而这一变化在茭蒿叶片中未有显著变化。由于栅栏组织与海绵组织的分化程度与水分状态密切相关,可见55%或35%θ土壤水分条件对黄花蒿的影响远远大于其余3种植物。叶厚度增加也有利于防止水分的过分蒸腾[16]。同样黄花蒿的叶片厚度在干旱下是4种植物种增幅最大者,茭蒿未有显著变化。
气孔是植物控制水分和进行气体交换的通道, 而气孔调节是植物抵御干旱和适应环境的机制之一,且气孔的分布特征、密度和面积等受环境水分状况的影响[36]。气孔密度随着环境中水分和湿度减少而增加, 但气孔气孔面积则向小型化发展, 气孔多下陷形成气孔窝或其上有突出的角质膜[36- 37]。但也有研究结果显示, 气孔密度并不随干旱程度的加剧而一直升高, 而是先升高后下降[38]。4种植物的气孔密度部分表现出增加趋势;尤其是黄花蒿上表皮气孔分布在重度干旱下由无到有的变化与其相等面叶发生变化一致;而铁杆蒿和茭蒿的上表皮气孔密度在干旱下下降,这有利于干旱下的节水,但同时也限制了CO2的扩散,不利于光合,但是其变化与铁杆蒿的上表皮无表皮毛的现象和干旱下上表皮角质层增厚综合来看这更有利于节水调节功能。由于干旱对叶面积扩展的限制而面积则向小型化发展导致气孔密度增加, 所以气孔指数常作为气孔分化强度的指标[39]。早期研究认为,气孔指数受湿度而不是光强的影响[40],但这种环境条件对气孔分化的调节机理至今仍不清楚。通过气孔指数表明干旱通过影响表皮细胞及气孔发育来改变气孔密度,从而对水分利用进行调控。不同水分梯度下4种植物的气孔指数显示均与气孔密度变化一致,可见气孔密度的增加主要由于干旱对气孔分化过程造成的影响,而非主要由叶片变小而导致气孔密度增加。但小而密的气孔同时也具有较高的灵活性[36], 因此有利于植物保持体内水分和保证有效的光合作用, 是植物适应旱生环境的表现。4种植物叶片气孔器长、宽干旱下均有所下降,向小气孔发展,并且铁杆蒿和茭蒿的气孔在重度干旱下变宽。
叶绿体是植物光合作用、能量流动的重要细胞器。茭蒿的基粒和基质片层较其它3种植物浓密,又加上茭蒿上表皮毛稀疏,这对于强辐射地区的植物来说,会导致光能过程引起氧化损伤,这一结果与王勇等[34]研究发现茭蒿的脂质过氧化程度和抗氧化酶、抗氧化剂,特别是抗坏血酸含量在4种蒿类中最高相适应,说明茭蒿的抗氧化保护机制。猪毛蒿也具有与茭蒿相似多而小、基粒较厚的的叶绿体,这与两者都是等面叶相关;由于猪毛蒿上表皮毛浓密,可防止过剩光能,这与茭蒿依靠生理过程防止过剩光能同为适应策略。但两者在重度干旱下也均引起大量脂质体的出现(图版1,17、18;图版2,26)。铁杆蒿和黄花蒿无上表皮毛,这导致其叶绿体随时处于光能过剩状态,因此两者的叶绿体基粒都较薄,基粒片层较稀疏,且细胞内叶绿体数目较少。
综上所述,由于各自的叶片遗传结构特征存在差异,导致4种植物干旱下的适应机制不同。茭蒿的上表皮毛稀疏,不利于保水和抵抗强光辐射,再加上叶绿体基粒片层浓密,很容易导致光损伤。因此茭蒿具有较低上表皮气孔密度和较厚的上表皮角质层来减少蒸腾,并且形成较强的抗氧化保护机制[34]。而下表皮有浓密的表皮毛,有利于保持水分,从而有4种植物中最大的气孔密度,以保证足够的CO2的扩散。干旱下通过减少上表皮栅栏组织厚度来降低光吸收;同时降低海绵组织来保证叶紧实度和减少上表皮气孔分化来保水。除此之外,茭蒿的叶片厚度,上下表皮厚度,紧实度等都在4种植物中居中,对干旱的响应最弱,表现出较高的水分利用效率,应具有最佳的适应策略。
与之相对的黄花蒿叶片异面叶,上下表皮均无表皮毛;在4种蒿中具有最薄的叶片厚度、角质层厚度和叶片紧实度,这些均不利于保持水分。所以在进化上表现出气孔全部分布于下表皮,胞质内叶绿体稀疏,基粒厚度小而松散来防止光能过剩,但为了保证足够光合作用叶绿体又发育的较大。干旱下其叶片结构响应变化也最大:叶片增厚,下表皮栅栏组织的出现,以及叶片紧实度的增加等,但这些变化后的值也均小于3种旱生型未变化的值;再有干旱下其生长减弱,叶片上下表皮气孔分化大大加强来增加CO2气体交换,对于无表皮毛的黄花蒿叶片来说是得不偿失;更甚者是生长受到抑制,叶片上下表皮角质层厚度也减小,从而叶片水分保持更低。
与黄花蒿叶片形态相似的是铁杆蒿叶片,但下表皮毛浓密。其上表皮无表皮毛,同样不利保水和防止强光辐射;但是在4种植物中其有最厚的上表皮角质层,栅栏组织,上表皮厚度和叶片紧实度;且上表皮有低水平的气孔分布;叶绿体大而少但基粒稀疏;这些特征足够补偿无上表皮毛带来的缺陷。铁杆蒿叶片结构对干旱响应较黄花蒿小,茭蒿大,表现为:上表皮气孔分化降低,气孔变小,增加上表皮的角质层厚度来保持水分;下表皮毛浓密,有利保水,同时增加下表皮气孔分化来保证光合CO2交换。
猪毛蒿叶片的上下表皮均有浓密的表皮毛,因此上下均有气孔分布,且密度相当,有利于保水,防止强辐射,且具有和茭蒿相似的多而小、基粒较厚的叶绿体。干旱下,其叶片变厚,上下栅栏组织加厚,海面组织减少,紧实度增加,上下表皮角质层加厚保水,气孔分化增加。但是这些指标响应大部分在中度干旱下达到最佳值,重度度干旱下可能由于生长限制严重而降低,结果与水分利用效率在重度干旱下上升一致。
由此可见:4种蒿属植物中黄花蒿的叶片解剖学特征受到干旱的影响变化最大,铁杆蒿和猪毛蒿次之,茭蒿叶片解剖学特征对干旱的响应最弱;其种之间响应的强弱差异反应了4种植物叶片对干旱的适应性和适应机制不同,包括水分的利用与保持,光合作用的维持与保护;3种优势种的干旱适应性显著强于黄花蒿;干旱下叶片解剖学的结论与干旱耐受性指标结果一致。
[1] Wang Y F. Vegetation Resources in the Loess Plateau and Their Reasonable Use. Beijing: China Press of Science and Technology, 1991:1- 20.
[2] Chen Y S, Zhang C L, Ding S Y, Zhang Z W. The taxonomy and analysis ofArtemisiaLinn. in the Loess Plateau of Northern Shaanxi. Acta Botanica Boreali-occidentalia Sinica, 1993, 13(3): 238- 245.
[3] Liu J H, Li D W, Liu G B, Li X L, Hou X L. Traits of water ecotypes and life-form spectrum of vegetation under canopy ofRobiniapseudoacaciaforests. Science of Soil and Water Conservation, 2008, 6(2): 95- 99.
[4] Wen Z M, Jiao F, Po Y J, Jiao J Y. Natural restoration of vegetation and dynamic changes of species diversity in gully regions on Loess Plateau—case study in Wuqi county. Research of Soil and Water Conservation, 2005, 12(1): 1- 3.
[5] Chen Y M, Liang Y M, Cheng J M. The zonal character of vegetation construction on Loess Plateau. Acta Phytoecologica Sinica, 2002, 26(3): 339- 345.
[6] Cheng J M, Wan H E, Du F. Recovery and reconstruction of degraded shrub-grass vegetation in semi-arid region of Loess Plateau. Scientia Silvae Sinicae, 2001, 37(4): 50- 57.
[7] Hou F J, Xiao J Y, Nan Z B. Eco-restoration of abandoned farmland in the Loess Plateau. Chinese Journal of Applied Ecology, 2002, 13(8): 923- 929.
[8] Zhu Z C, Huang K, Li J Z. Herbosa type and characteristics of succession on woodland of Loess Plateau in north Shaanxi province. Grassland of China, 1989, (3): 18- 24.
[9] Wang S H, Zhang F M. Effect of different water treatments on photosynthetic characteristics of cucumber plants growing in solar greenhouse. Chinese Bulletin of Botany, 2002, 19(6): 727- 733.
[10] Meng L, Li L X, Chen W F, Xu Z J, Liu L X. Effect of water stress on stomatal density, length, width and net photosynthetic rate in rice leaves. Journal of Shenyang Agricultural University, 1999, 30(5): 477- 480.
[11] Wu Y Y, Li D Q. Effects of soil water stress on osmotic adjustment and chloroplast ultrastructure of winter wheat leaves. Acta Agriculturae Boreali-Sinica, 2001, 16(2): 87- 93.
[12] Zhang Y P, Wang Z M, Wu Y C, Zhang X. Stomatal characteristics of different green organs in wheat under different irrigation regimes. Acta Agronomica Sinica, 2006, 32(1): 70- 75.
[13] Zhu S H, Yu S L. Changes of cell and tissue ultrastructure of wine grape leaves under water stress. Transactions of the Chinese Society of Agricultural Engineering, 2004, 20(Z1): 73- 77.
[14] Xie S X, Zhang Q M, Xiong X Y, Deng Z N, Lovatt C J. Effect of water stress on leaf and root cell ultra-structure of Citrus. Journal of Hunan Agricultural University: Natural Sciences, 2008, 34(2): 168- 172.
[15] Qi H Y, Liu Y, Liu H T. Effect of water deficit on stomatal characteristics and ultrastructure of chloroplast in tomato leaves. Acta Botanica Boreali-Occidentalia Sinica, 2009, 29(1): 9- 15.
[16] Hu Y, Chu H J, Li J Q. Response of leaf anatomy characteristics and its plasticity to different soil water conditions ofMedicagoruthenicain four populations. Plant Science Journal, 2011, 29(2): 218- 225.
[17] Li Y H, Lu Qi, Wu B, Zhu Y J, Liu D J, Zhang D J, Zhang J X, Jin Z H. A review of leaf morphology plasticity linked to plant response and adaptation characteristics in arid ecosystems. Chinese Journal of Plant Ecology, 2012, 36(1): 88- 98.
[18] Chartzoulakis K, Patskas A, Kofidis G, Bosabalidis A, Nastou A. Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivers. Scientia Horticulturae, 2002, 95(1/2): 39- 50.
[19] Chen X, Xu Y F, Zhang Z Y. Leaf anatomical structure and photosynthetic physiology responses ofSorbusfolgneriseedlings under drought stress. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(1): 111- 116.
[20] Gong W C, Zhuang L, Zhao W Q, Tian Z P. Anatomical structure and ecological adaptability of two kinds of halophytes (HaloxylonammondendronChenopodiaceae andTamarixramosissimaAmaranthaceae). Acta Ecologica Sinica, 2009, 29(12): 6764- 6771.
[21] Sam O, Jeréz E, Dell′Amico J, Ruiz-Sanchez M C. Water stress induced changes in anatomy of tomato leaf epidermes. Biologia Plantarum, 2000, 43(2): 275- 277.
[22] Mendes M M, Gazarini L C, Rodrigues M L. Acclimation ofMyrtuscommunisto contrasting Mediterranean light environments effects on structure and chemical composition of foliage and plant water relations. Environmental and Experimental Botany, 2001, 45(2): 165- 178.
[23] Zhao Q F, Ma R J, Du G Z, Su N K, Wang G. Ecological signification of the leaf structure of three species of Ligularia in differential attitudes. Journal of Lanzhou University: Natural Science Edition, 2006, 42(1): 33- 39.
[24] Yang J Y, Yang J, Yang M B, Liang N, Kong W X, Qing H. Mechanisms of ecological adaptation ofCaraganastenophyllato drought stress in different habitats. Acta Botanica Boreali-Occidentalia Sinica, 2009, 29(12): 2476- 2482.
[25] Su D, Zhang J Z, Sun G F, Xing Q, Gu D F, Dong R. Studies on anatomical structures of leaves and their drought tolerance features ofSedumaizoonL. andSedumspectabilisBoreau. Bulletin of Botanical Research, 2007, 27(4): 428- 433.
[26] Li F L, Bao W K, Liu J H, Wu N. Eco-anatomical characteristics ofSophoradavidiileaves along an elevation gradient in upper Minjiang River dry valley. Chinese Journal of Applied Ecology, 2006, 17(1): 5- 10.
[27] Zhao J H, Li Q F. Drought resistance analysis based on anatomical structures of three Wild Allium in Inner Mongolia arid grassland. Journal of Inner Mongolia University, 2010, 41(2): 201- 205.
[28] Zhang X Q, Zeng J J, Chen J W, Luo Z W, Sun M. The saxicolous moss′s features of absorbing water and its structural adaptability in the heterogeneous environment with rock desertification. Acta Ecologica Sinica, 2012, 32(12): 3902- 3911.
[29] Du H D, Xu C H, Liu P, Jiao J Y, Miao F. Foliar anatomical structures and ecological adaptabilities of dominant plants in the north Shaanxi Loess Plateau. Acta Botanica Boreali-Occidentalia Sinica, 2010, 30(2): 293- 300.
[30] Xu H M, Gao Q, Huang Y M, Jia H K. Photosynthetic characteristics of six plant species in a forest steppe of the Loess Plateau, China. Acta Phytoecologica Sinica, 2004, 28(2): 157- 163.
[31] Ma X H, Jiao J Y. Characteristics of vegetation with natural restoration in removal lands in loess hilly-gully region and the relationship between the characteristics and soil environment. Science of Soil and Water Conservation, 2005, 3(2): 15- 22.
[32] Wang J, Yang C, Yin J, Wang T J, Liu P T. Changes of the genetic diversity ofArtemisiafrigidapopulation under the disturbance of grazing. Acta Ecologica Sinica, 2004, 24(11): 2465- 2471.
[33] Yang C, Liang Z S. Foliar anatomical structures and ecological adaptabilities of dominantArtemisiaspecies of early sere of succession on arable old land after being abandoned in Loess Hilly Region. Acta Ecologica Sinica, 2008, 28(10): 4732- 4738.
[34] Wang Y, Han R L, Liang Z S. Anti-oxidative responses to soil water stress for fourArtemisiaspecies. Journal of Northwest A & F University: Natural Science Edition, 2010, 38(10): 178- 186.
[35] He J S, Chen W L, Wang X L. Morphological and anatomical features ofQuercussection suber and its adaptation to the ecological environment. Acta Phytoecologica Sinica, 1994, 18(3): 219- 227.
[36] Li F L, Bao W K. Responses of the morphological and anatomical structure of the plant leaf to environmental change. Chinese Bulletin of Botany, 2005, 22(S1): 118- 127.
[37] Bosabalidis A M, Kofidis G. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Science, 2002, 163(2): 375- 379.
[38] Zhang X Y, Yang H M, Hou Z D, Wang G X. Stomatal densities and distributions of spring wheat leaves under different planting densities and soil moisture levels. Acta Phytoecologica Sinica, 2003, 27(1): 133- 136.
[39] Masle J, Gilmore S R, Farquhar G D. The ERECTA gene regulates plant transpiration efficiency inArabidopsis. Nature, 2005, 436(7052): 866- 870.
[40] Tichá I. Photosynthetic characteristics during ontogenesis of leaves. 7. Stomatal density and sizes. Photosynthetica, 1982, 16(3): 375- 471.
参考文献:
[1] 王义凤. 黄土高原地区植被资源及其合理利用. 北京: 中国科学技术出版社, 1991:1- 20.
[2] 陈彦生, 张春林, 丁士友, 张振万. 陕北黄土高原蒿属植物的分类与分析. 西北植物学报, 1993, 13(3): 238- 245.
[3] 刘江华, 李登武, 刘国彬, 李小利, 侯禧禄. 刺槐林下植被的水分生态型和生活型谱特征. 中国水土保持科学, 2008, 6(2): 95- 99.
[4] 温仲明,焦峰,卜耀军, 焦菊英. 黄土沟壑区植被自我修复与物种多样性变化——以吴旗县为例. 水土保持研究, 2005, 12(1): 1- 3.
[5] 陈云明, 梁一民, 程积民. 黄土高原林草植被建设的地带性特征. 植物生态学报, 2002, 26(3): 339- 345.
[6] 程积民, 万惠娥, 杜峰. 黄土高原半干旱区退化灌草植被的恢复与重建. 林业科学, 2001, 37(4): 50- 57.
[7] 侯扶江, 肖金玉, 南志标. 黄土高原退耕地的生态恢复. 应用生态学报, 2002, 13(8): 923- 929.
[8] 朱志诚, 黄可, 李继瓒. 陕北黄土高原森林地带草本植物群落类型及其动态特征. 中国草地, 1989, (3): 18- 24.
[9] 王绍辉, 张福墁. 不同水分处理对日光温室黄瓜叶片光合特性的影响. 植物学通报, 2002, 19(6): 727- 733.
[10] 孟雷, 李磊鑫, 陈温福, 徐正进, 刘丽霞. 水分胁迫对水稻叶片气孔密度、大小及净光合速率的影响. 沈阳农业大学学报, 1999, 30(5): 477- 480.
[11] 武玉叶, 李德全. 土壤水分胁迫对冬小麦叶片渗透调节及叶绿体超微结构的影响. 华北农学报, 2001, 16(2): 87- 93.
[12] 张永平, 王志敏, 吴永成, 张霞. 不同供水条件下小麦不同绿色器官的气孔特性研究. 作物学报, 2006, 32(1): 70- 75.
[14] 谢深喜,张秋明, 熊兴耀, 邓子牛, Lovatt C J. 水分胁迫对柑橘叶片和根系细胞超微结构的影响. 湖南农业大学学报: 自然科学版, 2008, 34(2): 168- 172.
[15] 齐红岩, 刘洋, 刘海涛. 水分亏缺对番茄叶片气孔特性及叶绿体超微结构的影响. 西北植物学报, 2009, 29(1): 9- 15.
[16] 胡营, 楚家海, 李建强. 4个花苜蓿居群叶片解剖结构特征及其可塑性对不同水分处理的影响. 植物科学学报, 2011, 29(2): 218- 225.
[17] 李永华, 卢琦, 吴波, 朱雅娟, 刘殿君, 张金鑫, 靳占虎. 干旱区叶片形态特征与植物响应和适应的关系. 植物生态学报, 2012, 36(1): 88- 98.
[19] 陈昕, 徐宜凤, 张振英. 干旱胁迫下石灰花楸幼苗叶片的解剖结构和光合生理响应. 西北植物学报, 2012, 32(1): 111- 116.
[20] 公维昌, 庄丽, 赵文勤, 田中平. 两种盐生植物解剖结构的生态适应性. 生态学报, 2009, 29(12): 6764- 6771.
[24] 杨九艳, 杨劼, 杨明博, 梁娜, 孔维雪, 清华. 不同生境狭叶锦鸡儿对干旱胁迫的生态适应机理研究. 西北植物学报, 2009, 29(12): 2476- 2482.
[25] 苏丹, 张金政, 孙国峰, 邢全, 顾德锋, 董燃. 费菜和长药八宝叶形态结构及其与耐旱性关系的研究. 植物研究, 2007, 27(4): 428- 433.
[26] 李芳兰, 包维楷, 刘俊华, 吴宁. 岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究. 应用生态学报, 2006, 17(1): 5- 10.
[27] 赵金花, 李青丰. 内蒙古荒漠草原三种野生葱属植物解剖结构的抗旱性分析. 内蒙古大学学报: 自然科学版, 2010, 41(2): 201- 205.
[28] 张显强, 曾建军, 谌金吾, 罗正伟, 孙敏. 石漠化干旱环境中石生藓类水分吸收特征及其结构适应性. 生态学报, 2012, 32(12): 3902- 3911.
[29] 杜华栋, 徐翠红, 刘萍, 焦菊英, 苗芳. 陕北黄土高原优势植物叶片解剖结构的生态适应性. 西北植物学报, 2010, 30(2): 293- 300.
[30] 徐红梅, 高琼, 黄永梅, 贾海坤. 黄土高原森林草原区6种植物光合特性研究. 植物生态学报, 2004, 28(2): 157- 163.
[31] 马祥华, 焦菊英. 黄土丘陵沟壑区退耕地自然恢复植被特征及其与土壤环境的关系. 中国水土保持科学, 2005, 3(2): 15- 22.
[32] 王静, 杨持, 尹俊, 王铁娟, 刘朋涛. 冷蒿种群在放牧干扰下遗传多样性的变化. 生态学报, 2004, 24(11): 2465- 2471.
[33] 杨超, 梁宗锁. 陕北撂荒地上优势蒿类叶片解剖结构及其生态适应性. 生态学报, 2008, 28(10): 4732- 4738.
[34] 王勇, 韩蕊莲, 梁宗锁. 水分胁迫对4种菊科蒿属植物抗氧化特性的影响. 西北农林科技大学学报: 自然科学版, 2010, 38(10): 178- 186.
[35] 贺金生, 陈伟烈, 王勋陵. 高山栎叶的形态结构及其与生态环境的关系. 植物生态学报, 1994, 18(3): 219- 227.
[36] 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应. 植物学通报, 2005, 22(增刊): 118- 127.
[38] 张晓艳, 杨惠敏, 侯宗东, 王根轩. 土壤水分和种植密度对春小麦叶片气孔的影响. 植物生态学报, 2003, 27(1): 133- 136.
Effect of drought on leaf anatomical characteristics of fourArtemisiaspecies in the Loess Plateau
WANG Yong1,2,LIANG Zongsuo1,2,GONG Chunmei2,HAN Ruilian1,*, YU Jing2
1InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistry,Yangling712100,China2TheCollegeofLifeScience,TheNorthWestAgricultureandForestUniversity,Yangling712100,China
Leaf anatomical characteristics of 4Artemisiaspecies under drought conditions were investigated to reveal the drought-resistant strategies of vegetation at early stage of ecological succession in the Loess Plateau. Three dominant species asArtemisiascopariaWaldst. et Kit.,ArtemisiagiraldiiPamp. andArtemisiagmeliniiWeb. ex Stechm. and one common specie,ArtemisiaannuaLinn. during early succession in the Loess Plateau were selected to investigate the leaf anatomical adaptation ofArtemisiato soil drought stresses. Pot experiment was adopted to control the soil water content. And 75%, 55% and 35% of field moisture capacity were set as control, moderate water stress and severe water stress, respectively. The latest mature leaves were freshly collected and fixed at the end of their vegetation growth period for SEM, TEM and the optical microscope. We analyzed the difference among leaf epidermis characteristics of the fourArtemisiaspecies according to the sub-cellular micro characteristic and the organizational anatomical structure. The results show that: the four Artemisia leaves have some kind of anatomy characteristics, as blade thickness, palisade tissue thickness, cutin layer thickness, ratio of palisade tissue thickness to sponge tissue thickness, tightness of leaf tissue structure were increased under drought conditions. Leaf epidermises ofA.scoparia,A.giraldiiandA.gmeliniihave dense trichomes to prevent excessive water loss from evaporation; howeverA.annuahas sparse trichomes. The upper epidermises ofA.gmeliniiandA.annuaare sparse, which are not good for water conservation and to prevent highlight, the decrease of the stoma density and the number of chloroplast were conducted to adapt environment change. Similar chloroplast anatomy characteristics of thick grana and dense grana lamellae were observed in the leaf cells ofA.scopariaandA.giraldii, which are easier to result in oxidative damage during photosynthetic process. To prevent the oxidative damage, the dense trichomes on upper epidermises ofA.scopariaand outstanding antioxidant mechanisms ofA.giraldiiwere conducted, and a large number of liposomes were also produced in the two species. The leaf anatomical characteristics influence of drought affected on theA.annuawas the greatest between the fourArtemisiaspecies, andA.gmeliniiandA.scoparawere followed, the least affect was onArtemisiagiraldii. The three dominant species had stronger ability to adapt to drought stress than the common species-Artemisiaannua. The leaf anatomical characteristics and drought tolerance indexes of the 4 species represented similar comments of drought tolerance ability.
Artemisia; characteristic of stomata; foliar anatomical structures; drought resistance
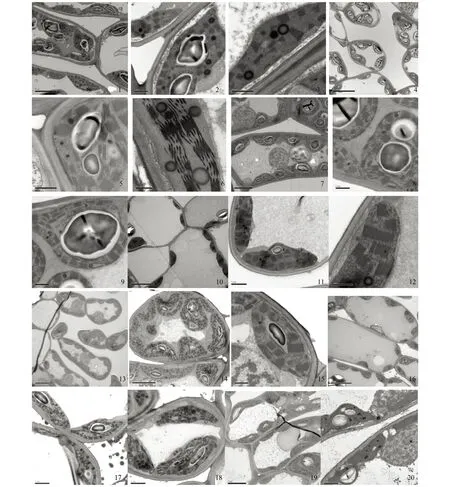
图版1 蒿属植物叶片亚细胞结构(1)1—3:铁杆蒿适宜水分A.gmelinii under CK;4—6:铁杆蒿中度干旱A.gmelinii under Medial drought stress;7—9:铁杆蒿重度干旱A.gmelinii under Severe drought stress;10—12:猪毛蒿适宜水分A.scoparia under CK;13—15:猪毛蒿中度干旱A.scoparia under Medial drought stress;16—18:猪毛蒿重度干旱A.scoparia under Severe drought stress;19—20:茭蒿适宜水分A.giraldii under CK;

图版2 蒿属植物叶片亚细胞结构(2)1—3:铁杆蒿适宜水分A.gmelinii under CK;4—6:铁杆蒿中度干旱A.gmelinii under Medial drought stress;7—9:铁杆蒿重度干旱A.gmelinii under Severe drought stress;10—12:猪毛蒿适宜水分A.scoparia under CK;13—15:猪毛蒿中度干旱A.scoparia under Medial drought stress;16—18:猪毛蒿重度干旱A.scoparia under Severe drought stress;19—20:茭蒿适宜水分A.giraldii under CK;21: 茭蒿适宜水分A.giraldii under CK; 22—24:茭蒿中度干旱A.giraldii under Medial drought stress;25—27:茭蒿重度干旱A.giraldiii under Severe drought stress;28—29:黄花蒿适宜水分A.annua under CK;30—33:黄花蒿中度干旱A.annua under Medial drought stress;34—36:黄花蒿重度干旱A.annua under Severe drought stress
中国科学院重要方向项目(KZCX2-YW- 443); 中国科学院知识创新项目(KZCX2-XB2- 05- 01)
2013- 06- 30;
2014- 06- 12
10.5846/stxb201306301802
*通讯作者Corresponding author.E-mail: liangzs@ms.iswc.ac.cn
王勇,梁宗锁,龚春梅,韩蕊莲,于靖.干旱胁迫对黄土高原4种蒿属植物叶形态解剖学特征的影响.生态学报,2014,34(16):4535- 4548.
Wang Y,Liang Z S,Gong C M,Han R L, Yu J.Effect of drought on leaf anatomical characteristics of fourArtemisiaspecies in the Loess Plateau.Acta Ecologica Sinica,2014,34(16):4535- 4548.
猜你喜欢
《学习方法报》读写拍档新高考版(2023年8期)2023-04-29 00:44:03
儿童时代·快乐苗苗(2022年7期)2022-10-18 01:17:18
今日农业(2022年14期)2022-09-15 01:44:50
今日农业(2022年3期)2022-06-05 07:12:00
黄河之声(2020年16期)2020-11-05 01:57:46
今日农业(2020年16期)2020-09-25 03:04:32
河南农业(2019年12期)2019-12-06 08:19:12
作文大王·笑话大王(2018年11期)2018-12-13 22:40:51
意林·少年版(2018年16期)2018-09-12 18:54:12
作文评点报·作文素材小学版(2016年2期)2016-03-16 16:56:29
