
5种红树植物的叶片结构及其抗逆性比较1)
2014-07-27 07:59刘锴栋黎海利
东北林业大学学报 2014年7期
陈 燕 刘锴栋 黎海利
许方宏 钟军弟 成夏岚 袁长春
(广东湛江红树林国家级自然保护区管理局) (湛江师范学院)
5种红树植物的叶片结构及其抗逆性比较1)
陈 燕 刘锴栋 黎海利
(湛江师范学院,湛江,524048)
许方宏 钟军弟 成夏岚 袁长春
(广东湛江红树林国家级自然保护区管理局) (湛江师范学院)
研究了采自广东徐闻海岸滩涂的5种红树植物(红海榄(Rhizophorastylosa)、角果木(Ceriopstagal)、白骨壤(Avicenniamarina)、桐花树(Aegicerascorniculatum)和榄李(Lumnitzeralittorea))的叶片解剖结构,利用模糊隶属函数值法综合评价其抗逆性。结果表明:5种红树植物的叶片形成了适应海岸潮间带生境的结构,即具下生气孔,气孔密度较小;叶片和角质层较厚;形成下皮;栅栏组织细胞多层;富含单宁。种间抗逆性综合评价结果由大到小排序为白骨壤、桐花树、红海榄、榄李、角果木。
红树植物;叶片解剖结构;抗逆性
Journal of Northeast Forestry University.-2014,42(7).-27~31,68
We used five mangrove species leaves ofRhizophorastylosa,Ceriopstagal,Avicenniamarina,AegicerascorniculatumandLumnitzeralittorea, collected from Xuwen, Guandong, to study their leaf structural features and evaluate the stress resistances by subordinate function value analysis. Five species were adapted to aquatic environment in structure with stomata distributed in lower epidermis, lower stomatal density and thicker cuticle. The tannin content was high in the leaf blade. By subordinate function value analysis, the stress resistance capabilities of the five mangrove species were in order ofAvicenniamarina>Aegicerascorniculatum>Rhizophorastylosa>Lumnitzeralittorea>Ceriopstagal.
Keywords Mangrove; Leaf structure; Stress resistance
红树林分布于热带、亚热带陆地与海洋交界的海岸滩涂,是陆地向海洋过度的特殊生态系统,在防风消浪、促淤造陆、净化水体和调控生态平衡等方面发挥着重要的生态效能[1-2]。红树植物是专指生长在红树林中的木本植物,是红树林群落的构成物种[3]。红树植物长期适应潮汐周期性的淹浸生境,能耐渍水、耐盐土,但对低温较敏感。我国红树林主要分布区的年平均温度为23.5 ℃,最冷月均温16.6 ℃,低温0~6 ℃(个别-2 ℃);林鹏根据不同红树种类的天然分布区域,从生态学角度把中国红树植物划分为7个抗寒性等级[4]。
1980年代以来,国内外学者对红树植物生态学、生理学和群落学进行了广泛深入的研究[5-6],近年来在红树植物化学成分方面的研究成果也较多[7-9]。而红树植物生态解剖学和抗逆性的相关研究则较少。在生态解剖结构方面,Reimold[10]、林鹏[11]和邓传远[12]等研究了红树植物的木材解剖结构;Stace[13]研究了4种红树植物的叶片结构和生态适应;李元跃[3]、吴钿等[14]研究了红树、红海榄、秋茄、木榄和海莲5种红树的生态解剖结构,指出红树科植物具有很强的耐旱能力。在抗逆性研究方面,池伟等[15]观察了12种北移红树植物的越冬表现;郑松发等[16]探讨了5种红树植物适应缺氧生境的能力强弱;刘锴栋等[17]、陈燕等[18]分析和比较了角果木(Ceropstagal)和红海榄(Rhizophorastylosa)在低温胁迫下的抗逆生理变化,发现两种红树植物之间的生理响应存在一些差异。
叶片是植物进化过程中对环境变化较敏感且可塑性较大的器官,在不同选择压力下已经形成各种适应类型,其结构特征最能体现环境因子的影响或植物对环境的适应[19]。通过叶片的解剖构造与环境适应性的相关性研究来评判植物的抗逆性是一种简单易行又相对可靠的方法[20],该方法在陆生植物中已得到广泛应用[21-23]。目前,红树植物叶片解剖结构的研究多停留于对叶片横切结构的简单描述[3,24],至今未见利用叶片结构特征的数据统计分析进行红树植物抗逆性综合评价的相关报道。本试验以红海榄(Rhizophorastylosa)、角果木(Ceriopstagal)、白骨壤(Avicenniamarina)、桐花树(Aegicerascorniculatum)和榄李(Lumnitzeralittorea)5种红树植物的叶片为研究对象,观测叶片的生态适应结构,并选取相对稳定的结构特征结合模糊数学隶属函数法对5种红树植物的抗逆性进行综合评价,试图从叶的结构上寻求与抗逆性的关系,为红树植物引种造林的选育种提供科学依据与应用参考。
1 材料与方法
试验材料取自广东省徐闻县迈陈海岸滩涂红树林。该红树林带宽120~340 m,优势种为白骨壤和红海榄,由外滩(低潮带)至内滩(堤岸),5种红树植物的生态序列依次为:白骨壤、红海榄、桐花树、角果木、榄李。从林带中选取成熟、健康、长势一致的植株作样木,每一树种选择5株。从每一样木的东南面树冠上部剪取1个小枝(顶枝),每小枝带3对叶片,带回实验室用自来水冲洗干净叶片表面污物,晾干水分后备用。
叶横切制片:参照潘叶等[25]的方法进行。剪取枝条上第2对成熟叶片中脉两侧约5 mm×5 mm的小块,用FAA固定液固定后抽气,系列酒精脱水(65%、75%、85%、90%、95%、100%),然后用V(纯酒精)∶V(二甲苯)=1∶1混合液进行脱色,二甲苯透明,间隔时间为1 h,TB-718型生物组织自动包埋机以及TB-718型生物组织包埋机(冷台)进行石蜡包埋,Taiva R136轮转式切片机切片,厚度为10~12 μm,1%番红染色,中性树胶封片,制成永久切片。
叶表皮制片:选取各树种的第3对成熟叶片,用刮片法刮取新鲜叶片的上下表皮,剪取5 mm×5 mm规格,按照石蜡切片的制作方法,经过脱水、透明、染色、中性树脂封片等步骤制做成永久装片。
在NikonYS100光学显微镜下观察,并用易创YM310显微成像系统和MiE软件进行摄影及显微测量,平行测定100个值,取平均数。观测的指标包括:叶片厚度、上下角质层厚度、上下表皮厚度、栅栏组织厚度、海绵组织厚度、气孔密度等指标。同时计算叶片组织结构紧密度CT,R(%)、叶片组织结构疏松度SR(%)、以及栅栏组织/海绵组织值。
叶片组织结构紧密度CT,R=((下皮+内皮+栅栏组织厚度)/叶片厚度)×100%;
叶片组织结构疏松度SR=(海绵组织厚度/叶片厚度)×100%。
抗寒性综合评价:选取叶片紧密度、叶片疏松度和栅栏组织/海绵组织3个相对稳定的结构特征,利用模糊隶属函数法[26]综合评价5种红树植物抗寒性的强弱,隶属函数值计算公式为:
Ui,j=(Xi,j-Xjmin)/(Xjmax-Xjmin)。
式中:Xi,j为i植物种j性状值;Xjmin为j性状中最小值;Xjmax为j性状中最大值;Ui,j为i植物种j性状的抗寒隶属函数值。
如某一指标与抗寒性为负相关,可用反隶属函数计算其抗寒隶属函数值:
Ui,j=1-(Xi,j-Xjmin)/(Xjmax-Xjmin)。
数据分析:采用Excel软件对数据进行处理,应用DPS数据系统软件进行主成分分析和方差分析。
2 结果与分析
2.1 叶表皮形态特征
5种红树表皮细胞都呈不规则多边形,垂周壁式样多为平直,桐花树、角果木和榄李可见弓形壁(图1)。

A.白骨壤(下表皮内表面);B.红海榄;C.角果木;D.桐花树;E.榄李。
白骨壤、红海榄、桐花树和角果木的气孔只分布于下表皮(下生气孔),仅榄李的上、下表皮都有气孔分布。各树种的气孔密集排列于叶脉两侧,沿中脉横向减少;白骨壤和榄李的气孔属于无规则型,桐花树、角果木和红海榄的气孔为平列型。5个树种的气孔器皆呈长椭圆形(表1),保卫细胞长宽比为1.28~1.72,保卫细胞最大的是红海榄,其次是角果木,最小的是白骨壤;单位面积的气孔密度(个·mm-2)由大到小排序为白骨壤、榄李、桐花树、红海榄、角果木;而气孔由小到大的排序为白骨壤、榄李、角果木、桐花树、红海榄。
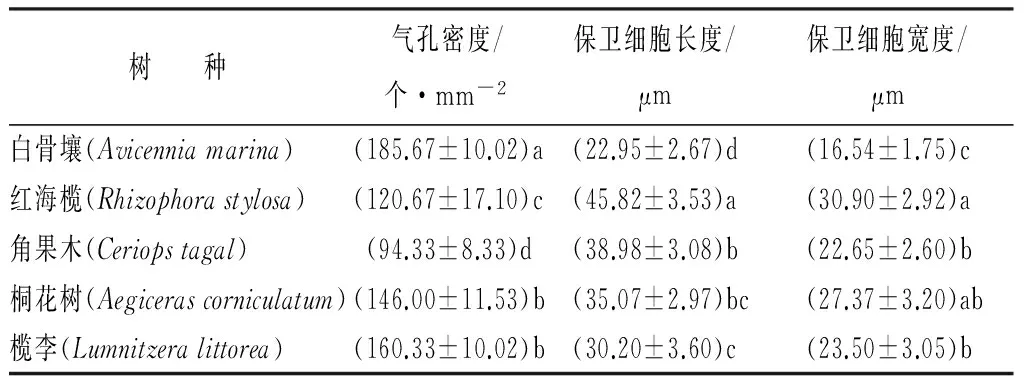
表1 5种红树植物的气孔特征
注:表中数据为平均值±标准差;同列相同字母表示差异不显著(P>0.05);不同字母表示差异显著(P<0.05)。
白骨壤下表皮密被腺毛,难于观察到气孔,但从下表皮的内表面(图1A)可观测到小而密度较大的气孔;其余4个树种的叶表皮均光滑无毛。桐花树和白骨壤的上下表皮均有盐腺分布,具有直接排盐作用,红海榄、角果木和榄李叶表面无盐腺结构。
2.2 叶横切面结构特征
5种红树的叶片横切面结构见图2,各项观测指标的统计结果列于表2。叶片最厚的红海榄达663.12 μm,最薄的桐花树只有360.17 μm,5个树种叶片厚度由厚到薄的排序为红海榄、榄李、角果木、白骨壤、桐花树。

A.白骨壤;B.红海榄;C.角果木;D.桐花树;E.榄李。

植物种叶片厚度/μm上角质层厚度/μm下角质层厚度/μm上表皮厚度/μm下表皮厚度/μm上皮层数目/层上皮层厚度/μm白骨壤(Avicenniamarina)(415.32±22.06)c(2.60±0.29)c(2.35±0.29)bc(20.09±0.55)a(17.56±1.46)bc7~9(198.25±19.45)b红海榄(Rhizophorastylosa)(663.12±33.12)a(4.51±0.37)b(3.34±0.35)a(15.22±1.85)c(14.52±1.68)c5~7(287.35±26.87)a角果木(Ceriopstagal)(440.83±24.14)c(5.23±0.47)ab(2.19±0.35)c(24.00±1.01)a(24.80±1.51)a1~2(52.77±4.80)d桐花树(Aegicerascorniculatum)(360.17±23.98)d(5.56±0.51)a(2.27±0.36)bc(16.94±0.90)c(22.64±2.51)a3~5(95.07±6.73)c榄李(Lumnitzeralittorea)(610.68±27.34)b(3.15±0.38)c(2.85±0.27)ab(22.01±0.27)b(19.28±1.02)b——

植物种下皮层数目/层下皮层厚度/μm栅栏组织厚度/μm海绵组织厚度/μm叶片紧密度/%叶片疏松度/%栅栏组织/海绵组织白骨壤(Avicenniamarina)——(95.03±7.37)bc(79.44±8.38)d70.6219.131.20红海榄(Rhizophorastylosa)1(33.25±4.07)b(105.79±6.89)b(199.14±12.13)c64.3030.030.53角果木(Ceriopstagal)1~2(46.25±5.49)a(51.48±8.16)d(235.11±18.23)b34.1453.340.22桐花树(Aegicerascorniculatum)2(48.02±7.91)a(82.05±11.79)c(86.62±10.89)d62.5124.050.95榄李(Lumnitzeralittorea)——(205.69±16.93)a(357.70±15.48)a33.6958.580.58
注:表中“—”表示该树种无此结构;表中部分数据为平均值±标准差;同列不同小写字母表示品种间差异显著(P<0.05)。
5个树种的叶表皮细胞外平周壁上有厚的连续分布的角质层,角质层厚度为2.19~6.51 μm,上角质层均厚于下角质层。桐花树的上角质层最厚,其次是角果木,最薄的是白骨壤。
白骨壤、红海榄、角果木和桐花树具复表皮,复表皮由表皮细胞与下皮(层)细胞共同构成,下皮细胞为大型的薄壁细胞,排列紧密,无明显胞间隙(为便于区别,文中将上表皮下方的皮层简称下皮,下表皮内侧的皮层简称内皮)。白骨壤下皮细胞层数最多,达7~9层,厚度达叶厚的47.74%;下表皮由单层细胞构成,无内皮,表皮外覆盖40~50 μm厚的腺毛层,每一腺毛由2~3个细胞组成,呈铁砧状,头部细胞多破裂状。红海榄叶片下皮由5~7层细胞构成,厚度也超过叶厚的43.34%,下皮细胞由小到大,嵌入栅栏组织;内皮只有1层,细胞较小,占叶厚的5.02%。桐花树的下皮3~4层,内皮多为2层,各占叶厚的26.40%、13.34%。角果木的下皮和内皮均为1~2层,分别占叶厚的11.97%和10.50%。榄李上下表皮均为单表皮,无下皮(内皮)结构。
5种植物的叶肉组织都有栅栏组织和海绵组织的分化,但栅栏组织的层数1~4层不等。角果木只有1层(异面叶);榄李靠上下表皮各有2层(等面叶);桐花树、红海榄和白骨壤为3~4层(异面叶)。栅栏组织细胞相对较小,排列紧密而较整齐,仅红海榄的栅栏组织因上皮组织细胞嵌入而呈山峰状不整齐排列。海绵组织厚度和疏松程度(细胞间隙)依树种不同而异,榄李、角果木、红海榄、桐花树和白骨壤的海绵组织厚度依次为357.70、235.11、199.14、86.62、79.44 μm,在高倍镜(400 x)下观察海绵组织,角果木和红海榄的细胞排列较疏松,胞间隙较大;白骨壤的胞间隙较小;桐花树的胞间隙不明显;而榄李海绵组织细胞排列相对紧密,难以观察到胞间隙,且细胞不含叶绿体,特化为贮水组织。
叶片细胞富含单宁也是红树植物的一个明显特征,不同树种中单宁的分布有一定差异。红海榄、角果木和桐花树的单宁主要分布于下皮、栅栏组织和海绵组织;榄李主要分布于栅栏组织及叶脉周围的海绵组织;白骨壤叶片细胞基本不含单宁。
根据叶片结构中的紧密组织(栅栏组织+下皮+内皮)与疏松组织(海绵组织)求得各树种叶片紧密度由大到小的排序为白骨壤(70.62%)、红海榄(64.30%)、桐花树(62.51)、角果木(34.14%)、榄李(33.69%)。5种植物叶片疏松度由大到小的顺序为榄李(58.58%)、角果木(53.34%)、红海榄(30.03%)、桐花树(24.05%)、白骨壤(19.13%)。从紧密度和疏松度可见,白骨壤、红海榄和桐花树的叶片结构偏于紧密,紧密组织比率超过62%;而榄李和角果木叶片结构相对较疏松,疏松度大于53%。5个树种的栅栏组织/海绵组织值由大到小依次为白骨壤、桐花树、榄李、红海榄、角果木,其中榄李和红海榄的栅栏组织/海绵组织值近于相等。
2.3 5种红树植物的抗逆性评价
植物对环境的适应结构可产生稳定的遗传,但一般情况下,植物单一物质或组织的数量如叶片厚度、栅栏组织和海绵组织厚度等往往会随着样品所处生态条件和生理状态的不同而发生变化[20,27];而叶片组织结构的紧密度、疏松度和栅栏组织/海绵组织等比值指标则反映了各组织之间在遗传上的相互制约关系,如紧密度反映栅栏组织、海绵组织和下皮紧密组织三者之间的相互制约关系。因此,紧密度等比值指标比单一组织的数量指标的遗传稳定性更好,常被用作抗逆性鉴定指标[20]。由于采用单一的抗逆性指标很难反映植物的抗逆性实质,而多指标的综合评价可以比较客观地反映物种的抗逆性。因此,本研究选取叶片紧密度、疏松度和栅栏组织/海绵组织3个相对稳定的遗传指标作为植物抗逆性的鉴定指标,利用模糊数学隶属函数法综合评价5种红树的抗逆能力。
多指标综合评价结果(表3)表明,白骨壤的抗逆隶属值的平均值最大,达0.666 7;其次是桐花树,为0.550 0;第三是红海榄,第四是榄李,二者的隶属值十分接近,最小的是角果木,仅有0.293 1。因此,5种红树植物的抗逆性排序由大到小依次为:白骨壤、桐花树、红海榄、榄李、角果木。
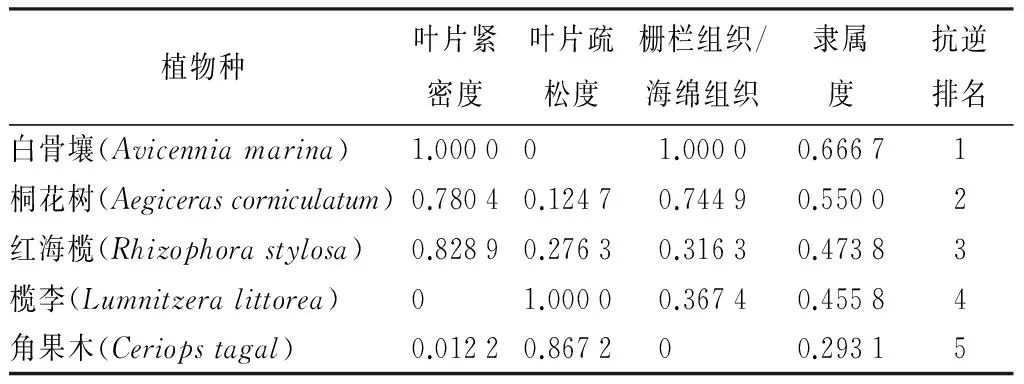
表3 5种红树的抗逆性指标的综合评价
3 结论与讨论
植物性状是由遗传因子和环境因子共同作用形成的。即遗传的多样性决定了植物的多样性,环境的多样性又深刻地影响着植物的多样性[19]。红树植物由不同科、属的树种组成,生长于潮汐浸淹、土壤盐渍化和通气不良的生境,经过长期的自然选择,使其演化形成了一系列趋同性的生态适应结构。
多具下生气孔。气孔密度较小(94.33~185.67个·mm-2),气孔面积较大(382.07~1 415.22 μm2)。5种红树植物的叶片形态接近中生植物,其与叶片体积缩小、气孔密度增大、气孔面积向小型化发展的典型旱生植物形态有较大差异[28]。
叶片和角质层较厚。叶片厚而坚硬,可以提供机械支持作用,利于抵抗风浪;角质层由脂肪性物质组成,加厚的角质能抑制水分散失,对避免水体污染入侵也具有意义。
常形成下皮组织。下皮由表皮细胞通过平周分裂向内侧产生的薄壁细胞组成,细胞大型,内贮大量水分[3]。研究表明,下皮在调节水分平衡控制蒸腾速率和防止细胞失水造成叶片皱缩等方面具有重要意义[29-30]。本研究发现,白骨壤、红海榄、桐花树、角果木、榄李的下皮细胞层数表现为递减趋势,这一排列顺序正好与5种植物的生态序列相一致,即生长分布越靠外滩前缘的种类,其下皮的层数越多;越靠内滩(近岸)的树种,其下皮的层数越少,甚至不形成下皮(榄李无下皮组织)。说明下皮的细胞层数与每日潮汐淹浸时间长短有明显的相关性,这种相关性是由盐度引起还是因淹水时间造成,有待进一步研究。Powman[30]认为美洲红树叶片下皮的数目与土壤水分中的盐度有关,但相关的研究文献甚为缺乏。
栅栏组织层数增加。除角果木的栅栏组织只有1层外,榄李、桐花树、红海榄和白骨壤栅栏组织都在2~4层。多层、小型的栅栏细胞大大增加了叶片的内自由表面,增强了光合作用的活动,并提高了蒸腾效率[28]。
叶片细胞富含单宁。研究认为,单宁或树脂等物质的主要作用是阻碍水分的流动[28]。多数红树植物的茎、叶都含大量的单宁[32],这可能是红树植物避免过度蒸腾引致生理干旱的重要防御机制。
5种红树植物来源于不同的科、属,受遗传基因控制,各树种除了上述的共同特征应对生境外,也形成了多样性的适应对策,例如,桐花树和白骨壤形成盐腺排出多余的盐分,避免盐胁迫引致生理干旱;白骨壤缺少单宁细胞,但形成大量的表皮毛覆盖下表皮,毛状体可能有隔离气流作用,避免叶肉组织过热或过冷能较好地调节水分和控制蒸腾;而角果木则以气孔器下陷来抑制叶的水分蒸腾[31]。榄李不形成下皮,但其海绵组织排列紧密、不含叶绿体,特化成贮水组织,发挥类似下皮的水分调节作用。
环境逆境是对植物生存生长不利的各种因素的总称,包括低温、高温、干旱、淹水、盐害等很多类型。红树植物的分布和生长发育受潮汐、盐度、极端低温等多种环境因素的综合影响。郑松发等[16]研究了海南清澜港的红树植物对淹水缺氧环境的适应,发现抗淹水能力排序与红树植物在潮滩上的天然分布带(生态序列)相一致,即适应能力越强者,分布位置越靠近前缘。本试验的5种红树植物由外滩至近岸的排序为白骨壤、红海榄、桐花树、角果木、榄李,这一排序与下皮组织细胞层数由多到少的排序高度一致,与叶片紧密度由大到小的排序(白骨壤、红海榄、桐花树、角果木、榄李)呈正相关,与叶片疏松度由大到小的排序(榄李、角果木、红海榄、桐花树、白骨壤)呈负相关,与栅栏组织/海绵组织值由大到小的排序(白骨壤、桐花树、榄李、红海榄、角果木)也有一定相关性。而叶片结构中的叶厚、角质层厚、栅栏组织和海绵组织厚度等单一组织的厚度排序规律与生态序列没有明显的相关性。因此,评价红树植物耐潮汐淹浸的能力,适宜选择下皮细胞层数、紧密度、疏松度和栅栏组织/海绵组织等结构指标。
极端低温是限制红树植物向高纬度分布的重要因子。林鹏等[33]根据红树植物天然分布北界划分出中国红树植物耐寒性等级序列,其中5种红树植物的抗寒能力表现为白骨壤、桐花树、红海榄、榄李、角果木。从本试验看,与抗寒力相关性较大的解剖结构指标有叶片疏松度、叶片紧密度和栅栏组织/海绵组织。一般是疏松度越大,抗寒力越弱;紧密度和栅栏组织/海绵组织较高者,抗寒力相对较强。叶片解剖结构中其他指标的排序与抗寒性排序规律差异较大,可能与其对抗寒性的贡献率较小有关。
综上所述,红树植物对淹水缺氧和对低温的抗逆性都与叶片结构中多种因素相关,总体上每一个因素与抗逆性之间都存在着一定的联系,即抗逆性由多种因素相互共同作用综合形成。在表现上,有的因素相关性较高,对抗逆的贡献率较大,而另一些因素的相关性较小,贡献率也较低。综合而言,红树植物叶片结构的疏松度、紧密度和栅栏组织/海绵组织3个指标与抗逆性相关性较大,但如果单纯选用某个单一的指标来进行抗逆性分析可能会存在偏差。因此,选取3个指标统计隶属函数,求算抗逆隶属值的平均值进行排序,所得的抗逆性强弱结果较为合理。本研究中5种红树植物的抗性由强到弱的顺序为白骨壤、桐花树、红海榄、榄李、角果木。
[1] 李庆芳,章家恩,刘金苓,等.红树林生态系统服务功能研究综述[J].生态科学,2006,25(5):472-475.
[2] 伍卡兰,彭逸生,郑康振,等.五种红树植物通气组织对人工非潮汐生境的响应[J].生态学报,2010,30(24):6927-6934.
[3] 李元跃,林鹏.三种红树植物叶片的比较解剖学研究[J].热带亚热带植物学报,2006,14(4):301-306.
[4] 林鹏.中国东南部海岸红树林的类群及其分布[J].生态学报,1981,1(3):283-290.
[5] 林鹏.红树林研究论文集[M].厦门:厦门大学出版社,1990:1-98.
[6] 黄玉山,谭凤仪.广东红树林研究论文选集[M].广州:华南理工大学出版社,1997:1-212.
[7] 赵亚,郭跃伟.真红树林植物化学成分及生物活性研究概况[J].中国天然药物,2004,2(3):135-140.
[8] 王继栋,董美玲,张文,等.红树林植物桐花树的化学成分[J].中国天然药物,2006,4(4):275-277.
[9] 易湘茜,高程海,何碧娟,等.红树植物木榄胚轴中苯丙素类化学成分研究[J].广西植物,2013,33(2):191-194,257.
[10] Reimold R J, Queen W H. Ecology of Halophytes[M]. New York: Acad Press,1974:51-74.
[11] 林鹏,林益明,林建辉.红树植物次生木质部的结构与进化[J].海洋学报:中文版,1998,20(4):97-102.
Leaf Stuctures and Stress Resistance in Five Mangrove Species/
Chen Yan, Liu Kaidong, Li Haili(Zhanjiang Normal University, Zhanjiang 524048, P. R. China); Xu Fanghong(The Administration of Zhanjiang Mangrove National Nature Reserve); Zhong Jundi, Cheng Xialan, Yuan Changchun(Zhanjiang Normal University)//
1) 广东省林业科技创新项目(2013KJCX011-03)、国家星火计划项目(2013GA780081,2013GA780093)、广东省科技创新项目(2013KJCX0124)、湛江市科技攻关计划项目(2012C0203,2013A30210)、湛江师范学院科研创新团队资助项目(2013CXTD05)、湛江师范学院博士启动项目(ZL09011)。
陈燕,女,1962年1月生,湛江师范学院生命科学与技术学院,副教授。
刘锴栋,湛江师范学院生命科学与技术学院,副研究员。E-mail:liukaidong2001@126.com。
2014年1月26日。
S718.47
责任编辑:任 俐。
猜你喜欢
小猕猴学习画刊(2022年4期)2022-05-05
老年博览·上半月(2021年10期)2021-11-02
海洋通报(2020年5期)2021-01-14
特区文学(2020年2期)2020-09-10
延河·绿色文学(2020年6期)2020-09-10
药学研究(2019年4期)2019-05-15
滇池(2019年1期)2019-02-14
中成药(2018年11期)2018-11-24
天然产物研究与开发(2016年6期)2016-06-05
红豆(2016年6期)2016-05-14
