
闽西北地区不同龄组常绿阔叶混交林生态系统碳储量结构特征
2014-05-08 03:37张治钟全林程栋梁徐朝斌胡波常云妮
生态环境学报 2014年2期
张治,钟全林, ,程栋梁, ,徐朝斌,胡波,常云妮
1. 福建师范大学地理科学学院,福建 福州 350007;2. 湿润亚热带山地生态国家重点实验室培育基地,福建 福州 350007
近年来,CO2浓度升高引起的全球变暖等一系列生态问题,成为世界环境领域关注的焦点。森林是陆地生态系统的主体,森林生态系统是陆地碳循环的主要组成部分。据统计,森林生态系统碳储量占全球陆地生态系统碳库总量的56%,其中森林植被碳储量占全球植被碳储量总量的80%以上。森林土壤碳库也储存了占全球土壤碳库 40%的碳(Dixon等,1994;Brown等,1999;Mathi等,1999;Houghton等,2001)。可见,森林在全球碳循环中起着非常重要的作用。森林通过生长,从大气中吸存了大量的CO2,其贮存能力取决于森林类型、种类组成、林龄及其与人类活动的关系(IPCC,2000; Houghton等,2000)。
亚热带常绿阔叶林是地球上的主要植被类型之一,并集中分布于我国,是我国亚热带山地的地带性森林植被类型,不仅发育充分,种类组成繁多,而且群落结构复杂,类型多样,具有重要的经济、社会和生态效益(张新时,2007)。目前,对于亚热带地区人工林生态系统碳储量的研究较多,而在区域尺度上对常绿阔叶混交林生态系统碳储量的研究较少。本研究选取福建西北部地区多群落类型组成的常绿阔叶混交林,通过典型样地调查取样,分析了闽西北地区常绿阔叶林生态系统不同发育阶段碳储量及其格局特征,为合理估算亚热带常绿阔叶林生态系统碳储量和区域尺度上森林生态系统碳库提供参考依据和基础数据。
1 研究区自然概况
福建闽西北地区包括南平、三明、龙岩、宁德四市,样地分布于 116°01′~119°06′E,25°14′~27°49′N,海拔 233~738 m,坡度以 15°~35°居多,为复杂的低山丘陵地区。其东部为戴云山脉、玳瑁山脉,西部是武夷山脉,地理位置特殊。土地面积约8万km2,占全省的2/3,地域相对广阔,资源比较丰富。由于长期受到人类活动的影响,林相杂乱。典型的常绿阔叶林有栲树林(Castanopsis fargesii forest)、米槠林(C. carlesii forest)、甜槠林(Cadtanopsis eyrei forest)、苦槠林(Pleioblastus amarus forest)、罗浮栲林(Castanopsis fabri forest)、青冈栎林(Quercus variabilis forset)、黧蒴栲林(Castanopsis fissa forest)、木荷林(Schima superba forest)、福建青冈栎(Cyclobalanopsis chungii forest)等。该区域属亚热带季风气候,年均温在 15~20 ℃,年均降水量1600~2500 mm,无霜期250~300 d。土壤为红壤,间有黄壤和红黄壤(陈伟,1996;廖成章等,2011)。
2 研究方法
2.1 样地设置
依据福建省2008年的二类森林资源调查资料中的各龄组常绿阔叶林面积与资源数据,结合其小班分布图,于2011年 8月至 10月,采用典型样地调查法在闽西北地区共设置了10常绿阔叶林调查样点,其中幼龄林2个、中龄林3个、近熟林2个、成熟林3个共10个样点,每个样点设置3个50 m×20 m的样地,样地间距不少于 100 m,测定并记录其经纬度与林分年龄、郁闭度等。将每个样地分成10个10 m×10 m的样方,对各样方的乔木层(胸径≥5 cm起测)树种进行逐一鉴别,测定胸径、树高。
样点所选设的区域都是远离居民点,人类活动干扰影响较小。各样点经纬度、林分年龄及其所包含的主要群落类型见表1。
2.2 乔木生物量测算
树高(H)与胸径(D)等测树因子与植株生物量有密切关系,因此本文采用“相对生长法”来估算植株的生物量。通过查阅文献,可以得到大量不同地区乔木生长关系式,根据各空间要素及气候指标等尽可能相同的原则,从中筛选出如下关系式,用于计算本区乔木生物量(李海奎和雷渊才,2010):
W干=0.044(D2H)0.9169
W皮=0.023(D2H)0.7115
W枝=0.0104(D2H)0.9994
W叶=0.0188(D2H)0.8024
W根=0.0197(D2H)0.8963
通过此关系式,得到样方内单株乔木生物量,加和得到样方内乔木总生物量,除以样方面积得到乔木层单位面积总生物量。
2.3 林下植被与枯落物生物量测算
灌木层(DBH﹤5 cm,高度﹥50 cm)的调查,通过在样地中随机设置3个2 m×2 m的样方,记录主要种类及盖度后,将样方内所有植物全部收获后,分根、枝干、叶称质量,分别取约300 g代表性样本称质量后,带回实验室烘干后测定其干质量。
在灌木调查样方(2 m×2 m)内,围取1个1 m×1 m的代表性小方框进行草本层调查,记录主要种类及其盖度后,将方框内所有植物全部收获,分地上部分和地下部分称质量,并分别取约 300 g代表性样本称鲜质量后,带回实验室,烘干后测定其干质量。
对于高度﹤50 cm的小灌木,归为草本层。
地表枯落物的调查,与灌草层调查同步。即,在测定灌草层生物量的样方内,选择代表性地段,布置1个1 m×1 m的代表性小方框,将样方框里的所有枯落物收集并称质量。同时,取 1个有代表性的子样品称质量后,带回实验室测定其干质量。
2.4 土壤调查
土壤调查采用剖面法+土钻法,其中土壤剖面主要用于测量土壤容重,土钻法主要用于测定土壤有机碳。
在50 m × 20 m的乔木样方内,按对角线方向,平均设置 3个土壤调查样点,在每 1个采土点上,按照 0~10、10~20、20~30、30~50、50~100 cm,5个层次采集样品,若土壤层无法达到100 cm,则采至基岩为止,记录实际深度。每层随机钻取 3钻土,混合成一个混合样。同一样地 3个土壤调查样点的同层次土样组成该层次混合样品。取样时尽量保持每个小土体的完整性,尽早进行风干处理。之后,沿剖面按0~10、10~20、20~30、30~50、50~100 cm 分层,用环刀采集各层土壤,带回实验室测定土壤容重。
2.5 样品采集与碳含量测定
对样地生物量贡献率占前 5位的 5个优势乔木树种取样,每种按大、中、小径级共选择3~5株样木,按叶、枝、干、根(2~5 cm)取样,同器官混合形成一个样品。每个样品鲜质量300 g左右,带回实验室烘干磨碎后,用于碳质量分数测定。用于碳质量分数测定的灌木、草本及枯落物样品,在样地外围采集,每个样品约300 g。将土钻取到的土壤样品装入自封袋,带回实验室,自然风干后,粉碎。所有样品过 100目筛,采用重铬酸钾-外加热容量法测定样品有机碳含量。

表1 样地位置及其包含的群落类型Table 1 The location and main community types of these samples
2.6 碳储量计算
生物碳储量为生态系统各层片生物量与相应碳素含量的乘积。
土壤碳储量的计算公式如下:
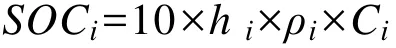
式中:SOCi为第 i层土壤有机碳含量(t·hm-2),hi为第 i 层土层厚度(m),ρi为第 i层土壤容重(g·cm-3),Ci为第 i层有机碳质量分数(g·kg-1)。
2.7 数据处理
对不同发育阶段各器官碳含量及生态系统个层次的碳储量进行方差分析及多重比较,使用软件为SPSS17.0。
3 结果与分析
3.1 植被及土壤碳含量
所调查的 4个龄组不同群落各组份的碳含量,随着发育阶段、器官及所在层片的不同而变化(图 1)。在每个发育阶段,各器官之间变化不大,碳含量的最大值51.4%(成过熟林树干)与最小值 46.4%(中龄林根)相差不足 10%。如图2,随着群落的发育,乔木各器官的碳含量,成过熟林树叶的碳含量极显著大于其余龄组(P<0.01),中龄林根的碳含量显著小于其余龄组(P﹤0.05),其余均无显著差异(P﹥0.05)。林下灌木在每个发育阶段,各器官(叶、枝、根)的碳含量均无显著差异(P﹥0.05),而各器官在不同阶段的碳含量,随着林分发育而变化,各器官碳含量的大小排序均为近熟林﹥中龄林﹥成过熟林﹥幼龄林。其中叶的碳含量幼龄林显著小于其余龄组(P﹤0.05),枝的碳含量幼龄林显著小于近熟林(P﹤0.05),根的碳储量近熟林显著大于成过熟林及幼龄林(P﹤0.05)。草本层地上部分的碳含量均大于地下。乔木层、灌木层、草本层的平均碳质量分数分别为48.5%、46.9%和41.2%,随层片高度不同而变化。凋落物碳含量在不同发育阶段无显著差异(P﹥0.05)。在每个发育阶段,土壤碳含量随土层深度变化而变化,由表层到底层,碳含量逐渐减小。在各个发育阶段,0~10、10~20 cm土层碳含量均显著大于其余3个土层(P﹤0.05),而0~10、10~20 cm 之间差异亦达到显著水平(P<0.01)。
3.2 植被及土壤碳储量
随群落的不断发育,各龄组乔木层碳储量大小排序为近熟林﹥成过熟林﹥中龄林﹥幼龄林(图3)。其中各组份在整个乔木层碳储量中的贡献率基本稳定,树干所占比例最大,为50%~52%,枝次之,为21.77%~26.45%,根为16.78%~17.58%,叶贡献率最小,为0.07%~0.08%。灌木层与草本层在不同发育阶段的碳储量,均表现出以下规律,从幼龄林到中龄林不断增长,在中龄林达到最大值后,又随发育的进行显现出不断下降的趋势。随着地表凋落物现存量的不断增加,其碳储量也表现出幼龄林﹥中龄林﹥近熟林﹥成过熟林的趋势。土壤碳储量随着林分发育,表现为幼龄林﹤中龄林﹤近熟林﹤成过熟林。表明随着林龄的增加,土壤碳储量不断增加。垂直空间上,在各发育阶段,0~30 cm土层在土壤碳储量的贡献率分别为 59.14%、54.05%、56.49%、60.19%,是土壤碳库的主体。
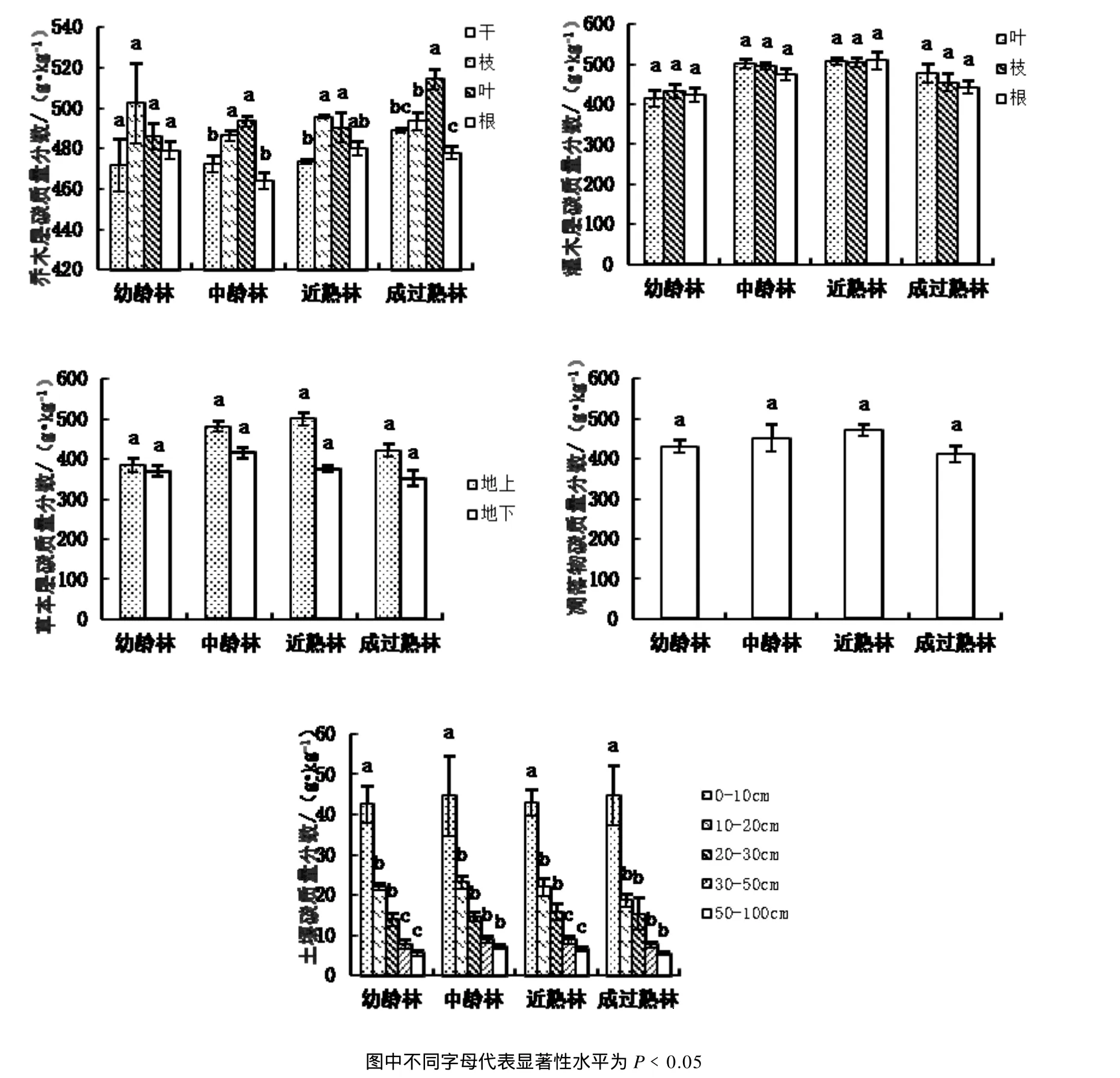
图1 不同龄组各组分碳含量Fig.1 Carbon concentration in each componenty of different age of stand
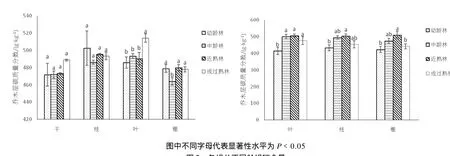
图2 各组分不同龄组碳含量Fig.2 Carbon concentration in different age of stand of each componenty
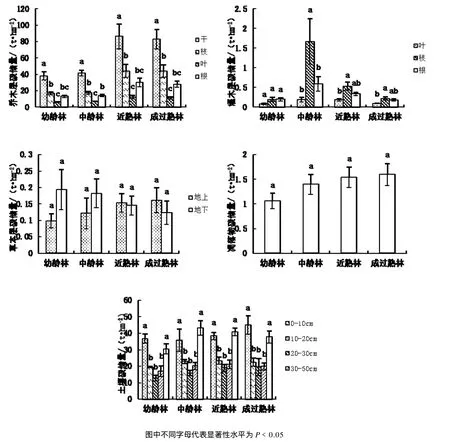
图3 不同龄组各组分碳储量Fig.3 Carbon storage in each componenty of different age of stand

图4 生态系统碳储量Fig.4 Carbon storage of the ecosystem
3.3 生态系统碳储量
幼龄林、中龄林、近熟林和成过熟林生态系统的碳储量分别为192.14、221.15、317.11和312.12 t·hm-2(图4)。凋落物与土壤碳储量也表现出随林龄增加而增加的趋势,乔木层在4个龄组对整个生态系统碳储量的贡献率为38.36%、35.97%、54.26%和53.04%。土壤层的贡献率为46.21%~62.16%,林下植被与凋落物碳储量贡献率微弱,仅占到整个生态系统碳储量的0.79%~1.87%。在每个发育阶段,各层片对整个生态系统碳储量的贡献率相对稳定,空间分布格局特征相似。
4 结论与讨论
4.1 讨论
本研究采用了碳含量的实测值估算闽西北天然常绿阔叶林碳储量。闽西北天然常绿阔叶林乔木层、灌木层和草本层各器官的碳质量分数变化范围分别为 46.4%~51.4%、41.4%~50.9%和 35.1%~49.9%,其平均碳质量分数分别为 48.5%、46.9%、41.2%。国际上在计算森林碳储量时,通常采用的转换系数为0.45~0.5,IPCC在 2003年出版的《土地利用、土地利用变化和林业优良做法指南》中,将全球森林植被碳质量分数的缺省值定为0.5(IPCC,2003)。可见,如果采用 0.45或 0.5的转换系数,势必会在一定程度上低估或高估本区域常绿阔叶林的碳储量。本文中,在每个龄组,乔木各器官碳含量变化不大,这与李铭红和于明坚(1996)及莫江明等(2003)之前的研究结果相似,反映了植物组织中碳水化合物含量的相似性。同时,随群落发育,乔木各器官在不同龄组间差异不明显,这也与刘恩等(2012)对南亚热带不同林龄红锥人工林的研究结果相似,说明林龄对乔木器官碳含量无显著影响。乔木层、灌木层、草本层的平均碳含量表现出随层片所在高度降低而减小的趋势,这可能与上层植物能够更多地吸收光照,进行充分的光合作用,积累更多的有机质有关。说明随植株高度及植物组织木质化程度降低,碳含量相应减少。同时,处于群落最下层的草本层地上部分碳含量普遍大于地下部分,从而显示出叶片作为光合作用的主要器官,在碳素同化中的重要性。林龄对凋落物碳含量无显著影响而分解状况对其影响显著(刘恩等,2012),本研究中凋落物的碳含量也与此规律相吻合。各龄组凋落物的碳含量无显著差异,所选取样地中乔木均以樟科、壳斗科为优势树种,凋落物性状相近,且调查对象为凋落物现存量,分解状况相似。
森林土壤是森林生态系统的一个重要碳库,在整个生态系统碳循环中发挥重要作用。本研究中,土壤的平均碳储量为134.986 t·hm-2,大于邻近地区杉木林土壤碳储量(68.581 t·hm-2)、福建柏人工林(102.818 t·hm-2)以及杉木观光木混交林(80.281 t·hm-2)(杨玉盛等,2003),这表明随着亚热带森林群落演替,土壤碳储量表现出针叶林﹤针阔混交林﹤常绿阔叶林的规律,这一结论也与宫超等(2011)对湖南鹰嘴界自然保护区内三种处于不同演替阶段森林土壤碳库的研究结果相吻合。同时,此结果与同地区甜槠林群落土壤碳储量(130.67 t·hm-2)(郎飞等,2012)相近,但远小于中国森林土壤平均碳储量(193.55 t·hm-2)和世界平均碳储量(189.00 t·hm-2)(周玉荣等,2000),这与森林土壤碳储量与植被类型、当地气候条件等因素有十分密切的关系。亚热带地区降水丰富,气温高,生物地球化学循环旺盛,有利于有机物质的分解,但不利于土壤碳素的积累,同时,闽西北地区雨量丰富且集中,山高坡陡,土壤抗侵蚀性差且易发生水土流失,土壤碳储量远低于高纬度地区以及我国森林土壤碳储量的平均水平。本研究中,4个龄组0~30 cm土层的碳储量占 0~100 cm总碳储量的比例分别达到59.14%、54.05%、56.49%、60.19%,表层土为森林土壤碳库的主体。因此,应加强本地区天然林植被的保护,较少人为干扰,保持地表水土,以增加土壤碳储量,对整个生态系统碳储量的积累有重要意义。
本研究中,闽西北地区常绿阔叶林生态系统平均碳储量为260.63 t·hm-2,略高于我国森林生态系统平均碳储量(258.83 t·hm-2)(周玉荣等,2000)。与同地区相比,大于 33年生杉木人工林(225.880 t·hm-2)(杨玉盛等,2003)与福建柏人工林(236.317 t·hm-2)(何宗明等,2003)以及中亚热带马尾松林(76.85 t·hm-2)(方晰等,2003),远大于中国热带、亚热带针叶林平均值(63.7 t·hm-2)(何斌等,2009)。同时,大于杉木观光木混交林(22.508 t·hm-2)。但与同地区常绿阔叶林生态系统相比,小于武夷山55年生甜槠林群落(315.13 t·hm-2)(郎飞等,2012)及 33年生格氏栲人工林(325.9 t·hm-2)(杨玉盛等,2006),高于30年生米槠次生林群落(178.67 t·hm-2)(李丽红,2012)。说明常绿阔叶混交林的碳储量要大于针叶纯林及针阔混交林,同时,常绿阔叶林之间也存在差异,可能是由于碳含量在不同树种之间的差异以及植株自身生物学特性导致的生物量的差异造成的。福建省森林碳储量以阔叶树为主,且在林龄构成上,以幼龄林和中龄林为主(王义祥,2004),说明闽西北地区随着阔叶树成熟和发展,将是一个巨大的潜在的碳汇。同时,在森林经营时,选用优良树种,采用合适的经营方式,对提高森林的经济效益与生态效益上有重要作用。
4.2 结论
(1)闽西北地区常绿阔叶林生态系统平均碳储量为260.63 t·hm-2。在每个发育阶段,各层片对整个生态系统碳储量的贡献率相对稳定,空间分布格局特征相似。幼龄林、中龄林、近熟林、成过熟林生态系统的碳储量分别为192.14、221.15、317.11 和 312.12 t·hm-2,基本表现出随林龄增加而逐渐增大的趋势。
(2)乔木层、灌木层、草本层、凋落物层的平均碳质量分数分别为48.5%、46.9%、41.2%、44.0%,每个层片中,各器官的碳含量差异不大,乔木层、灌木层及草本层的碳含量表现出随层片高度降低而减小的趋势。土壤碳含量由表层到底层逐渐减小。0~10、10~20 cm土层碳含量均显著大于其余3个土层。
(3)生物量碳储量在每个层片随着龄组不同,表现出不同的变化趋势。乔木层碳储量大小排序为近熟林﹥成过熟林﹥中龄林﹥幼龄林,灌木层与草本层在不同发育阶段的碳储量,均表现出以下规律:从幼龄林到中龄林不断增长,在中龄林达到最大值后,又随发育的进行显现出不断下降的趋势。随着地表凋落物现存量的不断增加,其碳储量也表现出幼龄林﹥中龄林﹥近熟林﹥成过熟林的趋势。土壤的平均碳储量为134.986 t·hm-2,随着林分发育,表现为成过熟林﹥近熟林﹥中龄林﹥幼龄林。
BROWN S L,SCHROEDER P,KERN J S.1999. Spatial distribution of biomass in forests of the eastern USA[J]. Forest Ecology and Management,123: 81-90.
DIXON R K,BROWN S,HOUGHTON R A,et al. 1994. Carbon pools and flux of global forest ecosystems[J]. Science,263: 185-190.
HOUGHTON J T,DING Y,GRIGGS D J. 2001a. Climate change 2001:The scientific basis.tribution of working groupⅠ to the third assessment report of the Intergovernmental Panel on Climate Change[M]. UK: Cambridge University Press.
HOUGHTON R A, SKOLE D L, NOBRE C A, et a1. 2000. Annual fluxes of carbon from deforestation and regrowth in the Brazilian Amazon[J].Nature,403: 301-304.
IPCC.2000. Land use.1and—use change,and forestry,a special report of the lntergovernmental Panel on Climate Change[M]. Cambridge University Press,U K.
IPCC.2003. Good Practice Guidance for Land Use,Land-use Change and Forestry[M].Hayama,Japan: IPCC/IGES.
Mathi Y,Baldocchi D D,Jarvis P G.1999.The carbon balance of tropical,temperate and boreal forests[J].Plant,Cell and Environment,22:715-740.
陈伟. 1996. 福建省森林生物多样性及其保护对策初探[J]. 福建林业科技, 23(4): 77-80.
方晰, 田大伦, 胥灿辉. 2003. 马尾松人工林生产与碳素动态[J]. 中南林学院学报, 23(2): 11-15.
宫超, 汪思龙, 曾掌权. 2011. 中亚热带常绿阔叶林不同演替阶段碳储量与格局特征[J]. 生态学杂志, 30(9): 1935-1941.
何斌, 黄寿先, 招礼军, 等. 2009. 秃杉人工林生态系统碳素积累的动态特征[J]. 林业科学, 45(9): 151-157.
何宗明, 李丽红, 王义祥. 2003. 33年生福建柏人工林碳库与碳吸存 [J].山地学报, 21(3): 298-303.
郎飞, 叶功富, 黄义雄, 等. 2012. 武夷山甜槠天然林含碳率与碳贮量研究[J]. 亚热带资源与环境学报, 7(4): 71-77.
李海奎,雷渊才.2010.中国森林植被生物量和碳储量评估[M].北京:中国林业出版社:58.
李丽红. 2012. 人促天然更新米槠次生林与杉木人工林乔木层碳贮量分配特征比较[J]. 亚热带资源与环境学报, 7(2): 63-69.
李铭红, 于明坚. 1996. 青冈常绿阔叶林的碳素动态[J]. 生态学报, 16(6):645-651.
廖成章, 洪伟, 吴承祯, 等. 2011. 福建中亚热带常绿阔叶林物种多样性的空间格局[J]. 广西植物, 23(6): 517-522.
刘恩, 王晖, 刘世荣. 2012. 南亚热带不同林龄红锥人工林碳贮量与碳固定特征[J]. Chinese Journal of Applied Ecology, 23(2): 335-340.
莫江明, 方运霆, 彭少麟. 2003. 鼎湖山南亚热带常绿阔叶林碳素积累和分配特征[J]. 生态学报, 23(10): 1970-1976.
王义祥. 2004. 福建省主要森林类型碳库与杉木林碳吸存[D]. 福州: 福建农林大学.
杨玉盛, 陈光水, 王义祥, 等. 2006. 格氏栲人工林和杉木人工林碳库及分配[J]. 林业科学, 42(10): 43-47.
杨玉盛, 谢锦升, 王义祥, 等. 2003. 杉木观光木混交林C库与C吸存[J].北京林业大学学报, 25(5): 10-14.
张新时. 2007. 中国植被及其地理格局[M]. 北京: 地质出版社: 226-235.
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522.
猜你喜欢
作物学报(2022年7期)2022-05-11
林业勘查设计(2021年2期)2021-04-08
土壤学报(2020年6期)2021-01-05
农民致富之友(2020年8期)2020-05-11
广东农业科学(2019年2期)2019-04-09
农业与技术(2018年16期)2018-11-28
科学与财富(2017年26期)2017-09-23
现代农业科技(2017年10期)2017-07-12
中国农业信息(2016年17期)2016-02-06
江苏农业科学(2014年9期)2014-11-15
