
遗传因素在风险决策加工中的作用
2014-04-29 00:44何清华薛贵陈春辉董奇陈传升
心理科学进展 2014年2期
何清华 薛贵 陈春辉 董奇 陈传升
编者按:
“人类如何进行决策”是近几十年来行为科学、经济学、心理学和神经科学等学科极为关注的重大科学问题。继2013年8月“The 1st International sytoposium on Decision Making and Brain Research”(第一届决策与脑研究国际研讨会)在北京成功举办后,中国心理学会决策心理学专业委员会(筹)11月又成功地在南京召開的第十六届全国心理学学术会议上,组织了1场分会重点报告和2场专题报告。为了更好地促进中国行为决策的研究与发展,本刊计划2014年伊始,开设一“决策心理学”专栏,邀请这2次重要会议的特邀报告人(Keynote speakers)l,为《心理科学进展》贡献一篇结合各自研究的最新研究进展报告。冀这一系列的特邀报告能促进我们对人类决策行为的理解,改善决策质量,并将该领域的研究成果真正应用于影响国计民生的重大决策中去。
《心理科学进展》主编:隋南
摘要:风险决策是指个体对不同选项及其概率进行权衡之后做出决定的过程。它是一个复杂的加工过程,需要平衡奖赏选项的诱惑和损失选项的忧虑。风险决策能力是人脑最重要的高级功能之一。在现实生活中,风险决策能力具有非常大的个体差异,而遗传和环境各自都在其中起着关键的作用。在这篇文章中,我们首先综述近年来风险决策研究领域中探讨遗传影响风险决策加工的研究,包括双生子研究与分子遗传学研究。在介绍分子遗传学研究时,按照基因所属神经递质系统,分别介绍了多巴胺递质系统相关基因(如COMT,DAT等)、五羟色胺递质系统相关基因(如SLC6A4、TPH1等)和其他基因(如BDNF)对风险决策能力个体差异的影响。随后,我们探讨了环境对风险决策个体差异的影响,以及基因,环境交互作用对风险决策个体差异的影响。接下来,我们介绍了将脑的结构与功能作为内表型来考察基因和环境对风险决策个体差异的影响的最新进展。在文章的最后,我们指出今后对风险决策个体差异的研究应该同时考虑遗传和环境,并考察脑的结构和功能在其中的中介作用。
关键词:遗传;基因;风险决策;环境;基因,环境交互;脑
分类号:B849.C91,B845
1、引言
日常生活中,人无时无刻不在做各种决定,可以小到穿哪件衣服、吃哪种食物,也可以大到与谁结婚、买哪栋房子。这些决定与我们的生活息息相关,甚至还影响到国家的发展和政策的制定,被称为决策(Decision Making)。决策在个人生活和社会发展中具有重要作用,对人类决策的研究吸引了来自哲学、人类学、政治学、经济学、社会学、心理学、认知科学以及神经科学等众多领域的研究者(薛贵,何清华,吕忠林,&Bechara,2012;Xue,Chen,Lu,&Dong,2010)。风险决策fRisky decision making)是指在各个选项的可能结果及其概率都已知的情况下所作出的决策,该方面的研究一直是决策研究中最活跃的领域之一。
虽然决策在人的日常生活中具有普遍性,但是不同人的决策也具有很大的差异性。即便是在相同的条件下,不同人的决策也千差万别,不同人也具有不同的决策特点(He et al.,2010)。对于风险决策也如此,而这些差别都来自于做决定的主体——人的差异。人的差异来自于各个方面,包括遗传的差异、环境的差异、甚至是脑结构和功能的差异。而对这些差异的研究将有助于我们全面了解风险决策个体差异的来源,有助于我们了解其背后的基因机制和神经机制,有助于对那些具有风险决策障碍的群体(主要是成瘾群体)进行有针对性的评估和治疗,同时还有助于我们对成瘾高危群体——青少年人群进行预防性干预。本文将详细介绍当前对风险决策个体差异的基因相关研究、基因一环境交互作用研究、以及以脑结构和功能为中介的基因/环境-脑-行为的研究,探讨其中遗传、环境、脑结构和功能各自分别对风险决策个体差异的贡献。
2、遗传因素对风险决策个体差异的影响
2.1 双生子研究
研究遗传因素对风险决策个体差异影响的传统研究方法就是采用双生子研究(Twin Study)。双生子研究通过比较同卵双生子(Monozygotic Twins,MZ)和异卵双生子(Dizygotic Twins,DZ)的同种行为差别计算出遗传力(Heritability),并以此来判断遗传因素对该行为的影响大小。在决策领域,已经有少量双生子研究开始关注遗传能够在多大程度上解释个体决策行为上的变异。
早期的研究很大程度上集中在社会决策方面(是指在两人或者多人之间进行的决策,包括竞争与合作等1,结果表明,人类的社会决策行为具有一定的遗传性。首先,Segal和Hershberger(1999)首次采用双生子的研究思路来研究人类决策行为。他们对59对MZ和37对DZ(年龄在10-83岁之间)施测了囚徒困境游戏。结果发现遗传与该游戏中合作的频率和持续性都有关系。随后,来自美国和瑞典的联合研究小组采用多种任务在双生子样本中考察了遗传对决策的影响(cesarini et al.,2008;Cesarini,Dawes,Johannesson,Lichtenstein,&Wallace,2009;Wallace,Cesarini,Lichtenstein,&Johannesson,2007)。Wallace等人(2007)采用最后通牒游戏,试图探讨遗传与公平偏好的关系。结果发现,虽然总体上,MZ和DZ在最后通牒游戏中的接受率没有显著差异;但是MZ之间的接受率相关为0.39,而DZ的接收率相关只有-0.04。此外,研究者还估计出遗传因素能解释42%的个体变异。Cesarini等人(2008)采用信任游戏对美国和瑞典两地的双生子进行研究,探讨遗传在信任和可信度的个体差异中的作用。结果发现,MZ之间的行为相似度高于DZ:在美国样本中,信任在MZ之间的相关系数为0.13,而在DZ之间为0.07;这个结果在可信度上也类似(0.26 vs,0.06)。在瑞典样本中,结果也相似(信任:0.25 vs,0.01;可信度:0.29 vs,0.18)。对于信任,美国样本的遗传力约为10%,瑞典样本约为20%。而对于可信度,美国样本的遗传力约为17%,瑞典样本约为18%。随后,Cesarini等人(2009)又考察了遗传在利他和风险决策行为中的作用。他们采用独裁者游戏来测量利他行为,用一个简单的赌博游戏(提供两个选项,选项1是肯定得到一定数量的钱;而另一个选项是50%的可能性赢得100元,50%的可能性什么也没有)来测量风险回避行为。采用风险投资任务来测量被试的风险投资行为(让被试假设他们买彩票中了100万,现在有个投资的机会,将有50%的概率收回双倍的投资,但是也有50%的概率只收回一半的投资,问被试希望投资多少钱)。并用一个0-10的量表(0代表一点也不愿意冒险,10代表非常愿意冒险)让被试自评其风险行为,即风险态度。结果发现,总体上这四个指标(利他行为、风险回避、风险投资和风险态度)上MZ和DZ之间都没有显著差异,但是在四个指标上都显示出MZ之间的相关系数高于DZ(利他行为:0.32 vs,0.11:风险回避:0.22 vs,0.03;风险投资:0.26 vs,0.10;风险态度:0.37 vs,-0.03)。遗传对利他行为的解释率为22%,而对风险回避的解释率为14%。
Cesarini等人(2009)的研究考察了遗传对社会决策的影响,但同时也考察了遗传对风险决策的影响。跟随着该研究的脚步,近年来,研究者们开始用双生子来考察遗传对风险决策的影响。比如,近期的一项研究报告了赌博行为的遗传力:对于男性,遗传因素可以解释85%的赌博行为变异,而女性则只可以解释45%(Beaver et al.,2010)。而另一項研究则没有发现类似的性别差异,而是发现对于所有人而言,遗传因素大致能解释50%的赌博行为变异(slutske,Zhu,Meier,&Martin,2010)。最近的一项纵向研究考察了遗传对爱荷华赌博测验(Iowa Gambling Task,IGT)得分的影响fTuvblad et al.,2012)。该研究在该组被试11-13岁、14-15岁以及16M8岁时分别施测了IGT,结果发现遗传因素能分别解释35%、20%和46%的决策变异。
这些研究都表明,遗传因素是影响个体风险决策的重要因素,对风险决策行为的个体差异起到了一定的解释作用(最低14%,最高85%)。但是,双生子研究只揭示了一种总体上的遗传可能性,而没法揭示遗传在风险决策加工中的具体机制。而随着基因技术的发展,分子遗传学的研究已经能够解答这个问题。
2.2 分子遗传学研究
基因是遗传的物质基础,是决定人体功能和健康的内在因素。人类基因组计划(HumanGenome Proiect,HGP)自从1990年正式启动到2003年正式完成,历经13年,受世人瞩目,取得了很多重要的成果。人类行为上的个体差异在很大程度上是由于不同人的基因差异导致的。最常见的基因变异有两种,一种是单个碱基上的变异,即单核苷酸多态性(single-Nucleotide Polymorphism,SNP)。另一种是碱基序列上的重复次数,被称为数目可变串联重复多态性(Variable Number of Tandem Repeats,VNTR)。比如多巴胺D4受体基因(DRD4)第3外显子上48bp重复多态性就具有普遍的重复2次(2R)、重复4次(4R)和重复7次(7R)的差别(Chen,Burton,Greenberger,&Dmitrieva,1999)。
到目前为止,对特定基因与风险决策个体差异关系的探讨主要集中在多巴胺和五羟色胺等神经递质系统的相关基因上,因为风险决策的加工过程总是伴随着奖赏和情绪fBechara&Damasio,2005;Bechara,Damasio,Damasio,&Anderson,1994;Bechara,Damasio,Tranel,&Damasio,1997;Bechara,Tranel,&Damasio,2000;He et al.,2010;Li,Lu,DArgembeau,Ng,&Bechara,2009),而多巴胺与奖赏的加工密切关系、五羟色胺与情绪加工的密切关系使得这两种神经递质系统相关基因在风险决策个体差异研究中得到重视。除此之外,还有一些其他基因也影响着人类复杂的决策过程。
2.2.1 多巴胺相关基因对风险决策个体差异的影响
多巴胺(Dopamine,DA)是大脑内一种重要的神经递质,属于儿茶酚胺(Catecholamine)的一种。多巴胺能神经元主要集中在中脑的黑质致密区(Substantia Nigrazona Compacta,SNpc)、腹侧被盖区(Ventral Tegmental Area,VTA)、下丘脑及其脑室周围。多巴胺能神经元由中脑的VTA投射到边缘系统,包括中隔(septum)伏隔核(NucleusAccumbens)、嗅结节(Olfactory Tubercle)、杏仁核(Amygdala)以及海马(Hippocampus),主要调控奖赏行为、短时记忆、自主活动、动机情绪以及适应性行为等(Mereu et al.,1995)。对多巴胺基因与风险决策个体差异的研究主要涉及三个与多巴胺相关的基因,它们是COMT,DAT和DRD4。
在多巴胺相关的基因中,研究得最多的是儿茶酚胺甲基转移酶(Catechol-O-Methyl Transsferase,COMT)基因,该基因负责编码COMT。COMT是最重要的DA降解酶,尤其是在前额叶皮层,60%以上的DA由它降解。人类COMT基因位于第22号染色体上,研究得最为广泛的位点是编号为rs4680的SNP也有研究涉及编号为rs4868的SNP(e.g.,Roussos,Giakoumaki,Pavlakis,&Bitsios,2008)。以rs4680位点(也被称为Vall58Met)为例,如果缬氨酸(Val)被甲硫氨酸(Met)取代,可引起编码出来的COMT热不稳定性的改变,从而导致COMT活性减低3.4倍(Lachman et al.,1996)。即是说,Val/Val基因型的酶具有高活性,Val/Met基因型的酶具有中度活性,而MetlMet基因型的酶活性最低。而因为COMT的作用是降解DA的,因此与Val/Val相比,Met/Met基因型的前额叶DA水平更高。
Roussos等人(2008)首次报告了COMT基因与风险决策任务的相关。他们采用IGT任务,并分型了被试的COMT rs4818基因多态性fG/G。G/C和c/c)。结果发现G/G基因型个体的IGT成绩比C/G和C/C基因型个体好;同时发现这个趋势在一项执行功能测验(stoekings of Cambridge)上正好相反。基因型在IGT上的作用主要体现在后期选择(风险决策)上,而在前20个选择(模糊决策)时没有差别。作者于是推论说COMT高表达活动(G等位基因)导致的前额叶较低的多巴胺水平有利于依赖情绪的决策加工,但是却不利于不依赖情绪的执行功能加工。van den Bos,Homberg,Gijsbers,den Heijer和Cuppen(2009)同样采用IGT任务,结果发现COMT MetlMet基因型被试的表现差于Val/Val,而这个差别也仅仅出现在IGT后期。他们还发现COMT与5-HTTLPR基因之间存在交互作用。Kang等人(2010)采用相同的任务和位点,却没有发现任何的COMT基因型效应(无论是模糊决策还是风险决策),也没有发现COMT与另一基因BDNF的交互作用。我们的研究也没有发现COMT基因对IGT任务得分的主效应,而是发现了基因一环境的交互作用(详细介绍见遗传环境交互对风险决策个体差异的影响部分)
另一个研究较多的多巴胺相关的基因是多巴胺转运体基因(Dopamine Transporter,DAT)。人类DAT基因位于第5号染色体上,研究得比较多的是一段40bp的VNTR,主要重复次数为3,13,表现出11种等位基因(3R到13R),其中9R和10B最为常见。研究已经发现,与IOR纯合子相比,9R等位基因携带者的中脑激活相对较高。在风险决策研究中,Zhong等人(2009)已經发现9R携带者在赢钱上更能容忍风险(较小的风险回避)。DreherKohn,Kolachana,Weinberger和Berman (2009)的研究既发现了DAT基因的主效应,还发现了DAT与COMT基因的交互作用。
第三个研究较多的多巴胺相关的基因是多巴胺D4受体基因(Dopamine Receptor D4,DRD4)。人类DRD4基因位于第11号染色体上,研究得比较多的是第3外显子上的一段48bp的VNTR,主要重复次数为2-11,其中2R、4R和7R最为常见。不同长度的基因编码出来的蛋白使得受体具有不同的活性,具有较长内环的D4受体与G蛋白的结合能力较低。Kuhnen和Chiao(2009)发现7R携带者在一项赌博任务上比非7R携带者冒更多的风险。而Ha,Namkoong,Kang,Kim和Kim(2009)采用IGT任务却没发现DRD4基因的主效应,但是他们发现DRD4与5-HTTLPR基因的交互作用影响IGT表现,尤其是风险决策阶段。
2.2.2 五羟色胺相关基因对风险决策个体差异的影响
与多巴胺一样,五羟色胺(serotonin/5-hVdroxytryptamine,5-HT)也是一种重要的中枢神经递质。大脑内部的5-HT能神经元主要分布于低位脑干的中线上,被称为中缝核群(Raphe Nuclei),,5-HT能神经元的纤维投射到广泛的大脑区域。这些脑区都与决策能力相关,包括前额叶(腹内侧、背外侧和眶额皮层)、杏仁核、纹状体和脑岛(Baumgarten&Grozdanovic,1999;Wai et al.,2008)。该类神经元在电生理方面有一个特点,即放电缓慢而规律。中枢神经系统5-HT含量及功能异常可能与多种情绪相关疾病(如抑郁症、双向障碍等)的发病有关。在风险决策的研究中,研究者们主要关注的是SLC6A基因的两个位点(5-HTTLPR和STin2),也有少量的研究者关注TPH1,TPH2和MAOA基因。
在5-HT相关基因中研究得最多的是五羟色胺转运体(5-HTT)基因SLC6A4[Solute CarrierFamily 6(Neurotransmitter Transporter,Serotonin),Member 4],且主要集中在该基因启动区一段44bp的片段长度多态性,被称作5-HTTLPRrSerotonin-Transporter-Linked Promoter Region),该位点的多态性(Short或s vs,Long或)影响转录的活性和合成的5-HTT活性。5-HTT多位于突触前膜,其特异性表现在调节突触前膜上钠离子依赖的神经递质回收,从而控制突触间隙中5-HT的浓度。5-HTTLPR,等位基因的转录活性比s高2.5倍,它能增加5-HTT基因的转录和5-HTT蛋白的表达,从而使5-HT的回吸收率增加。
在行为研究上,已经有一些研究开始探讨5-HTTLPR多态性与经济决策的关系(综述见He etal.,2010)。Homberg,van den Bos,den Heiier,Suer和Cuppen(2008)和van den Bos等人(2009)采用同一批被试发现总体上s/s基因型被试比,携带者在IGT任务上做得差(选择更多的坏牌),这个差异在后期(风险决策)更明显。而后者还发现了COMT基因与5-HTTLPR基因的交互作用,基因型为Met~Met和s/s的被试在IGT上的表现最差。而Ha等人(2009)和Stoltenberg和Vandever(2010)采用同样的IGT任务却在前后期都没发现5-HTTLPR的主效应。Ha等人(2009)发现了5-HTTLPR与DRD4基因的交互作用影响IGL尤其是风险决策阶段。Stoltenberg和Vandever(2010)则发现了性别与5-HTTLPR基因的交互作用,而且这个交互作用只在模糊决策时期出现。具体来讲,s携带者比l/l基因型被试选择了更多的好牌,而这个差异仅在男性被试身上出现。在风险决策方面,除了前面提到的IGT任务后半段的研究(Homberg et al.,2008;van den Bos et al.,2009)之外,还有一些研究者采用了其他任务来研究5-HTTLPR对风险决策的影响,比如仿真气球冒险任务fBalloon Analogue Risk Task,BARTl(crisan et al.,2009),投资任务(Kuhnen&Chiao,2009),框架效应(Crisan et al.,2009;Roiser et al.,2009)和多重价格列表任务(zhong et al.,2009)等。有的研究发现s与更多的风险回避有关,比如BART任务中较少的气球吹动次数(较少的冒险),投资任务中较少的投资(Kuhnen&Chiao,2009),更强的框架效应(Crisan et al.,2009;Roiser et al.,2009);但是Zhong等人(2009)的研究未发现显著的结果,只发现了类似的趋势。在病理上,研究者发现5-HTTLPR基因的,等位基因能显著增加自杀倾向者的IGT任务表现(Jollant et al.,2007),携带该等位基因的重度抑郁症患者(MaiorDepressive Disorder,MDD)在IGT上选择更多的好牌(Must et al.,2007)。
前人的这些研究的发现很不一致,究其原因可能有以下几个方面:1)首先,前人的研究采用了不同的任务。不同的任务可能具有不同的认知和情绪的加工机制,并且不同任务所依赖的具体脑功能也不尽相同,这些不同的脑区还可能分属不同的神经递质系统,以至于具有不同的基因机制(Stoltenberg&Vandever,2010)。因此,采用认知和神经机制都比较清楚的任务就显得尤为重要。2)其次,前人研究的样本量都相对较小。而小样本的研究其统计检验力较低,并会导致结果的不稳定性增加(Munafo,Brown,&Hariri,2008)。3)一些研究只考察了女性被试(Homberg et al.,2008;vanden Bos et al.,2009),,这样的设计使得我们无法观察决策中已经广泛存在的性别差异(Bolla,Eldreth,Matochik,&Cadet,2004;Overman et al.,2006;Reavis&Overman,2001),也无法考察性别与基因型的交互作用(Stoltenberg&Vandever,2010)。4)最后,这些研究大多数没有控制个体在基本认知能力(如智力和记忆等)上的个体差异,而基本认知能力是影响个体决策个体差异的重要因素(Gupta et al.,2009;Morsanyi&Handley,2008;Yechiam&Busemeyer,2005;Yechiam,Hayden,Bodkins,ODonnell,&Hetrick,2008)。
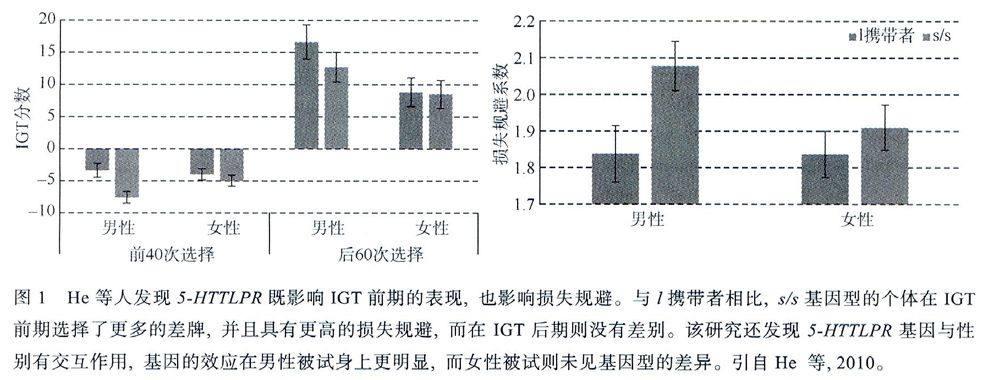
基于以上几个原因的考量,我们研究组(Heet al.,2010)在大样本(N=572)的中国汉族被试群体中考察了5-HTTLPR对决策的影响。该研究采用IGT和损失规避测验(LOSS Aversion Task,LAT)测查了模糊决策、风险决策和损失规避(风险决策的一种特殊形式,见Tom,Fox,Trepel,&Poldrack,2007)。该研究采用了认知和神经机制都较为清楚的两个风险决策任务在大样本的中国被试中考察了风险决策个体差异的基因机制。对于所有被试,该研究还采用韦氏智力测验和韦氏记忆测验测查了其基本认知能力,以排除基本认知能力对本研究所考察的基因机制的干扰。结果发现5-HTTLPR既影响IGT前期的表现,也影响损失规避,与,携带者相比,s/s基因型的个体在IGT前期选择了更多的差牌,并且具有更高的损失规避,而在IGT后期则没有差别(图1)。进一步的分析发现,5-HTTLPR基因还与性别有交互作用,基因的效应在男性被试身上更明显,而女性被试则未见基因型的差异(图1)。根据前人的脑损伤和fMRI研究,作者推论5-HTTLPR基因可能影响与决策密切相关的腹内侧前额叶(Ventromedial PrefrontalCortex,VMPFC)、杏仁核和脑岛等区域的激活,而女性则由于荷尔蒙的不规则,使得这种影响的变异较大,不容易做出差异來。
之前的研究已经发现5-HTTLPR基因多态性对风险决策个体差异的影响,但是这些研究主要基于正常被试群体。对那些具有决策障碍的临床样本的考察有助于我们深入理解基因影响决策行为背后的机制。同时,有研究表明rs2553,(A/G)是5-HTTLPR,等位基因的多态性位点,而七等位基因与s等位基因的功能相似(Hu et al.,2006)。这个三等位基因多态才是真正调节了五羟色胺转运体基因表达的功能性基因。突触间隙的五羟色胺除了受到五羟色胺转运体的调节外,还受到5-HTlA受体的调节(Bayliss,Li,&Talley,1997;Penington,Kelly,&Fox,1993)。编码5-HTlA受体蛋白的基因是5-HTRlA,在这个基因中rs6295是最经常被研究的单核苷酸多态。一些动物实验表明,五羟色胺转运体和5-HTlA受体这两个蛋白之间有一种类似于协同的生物联系。在5-HTlA受体被敲除的小鼠中5-HT转运体的浓度亦降低(Fabre et al.,2000;Li,Wichems,Heils,Lesch,&Murphy,2000;Li et al.,1999),同时在5-HT转运子被敲除的小鼠中5-HT的活性亦会减小fAse,Readeg Hen,Riad,&Descarries,2001)。最近,我们与李君的研究组(Gu et al.,2013)在正常被试和精神分裂症病人中考察了5-HTTLPR和5-HTRlA基因对风险决策的影响。结果发现在精神分裂症病人身上两个基因都显著影响IGT任务前40次选择的分数,表现为突触间隙五羟色胺水平越低,其IGT成绩越差。控制组的结果类似但不显著(Guet al.,2013)。
STin2(serotonin Transporter intron 2)位点与5-HTTLPR一样,都位于SLC6A4基因上,不同的是5-HTTLPR位于启动区,而STin2是该基因第2内含子上的17bp的VNTR,通常有10R和12R两种。目前只有一项研究发现STin2基因10R在输钱条件下的损失规避比12R更低(zhong et al.。2009)。色氨酸羟化酶(Tryptophan Hydroxylase,TPH)亚型基因TPHl和TPH2负责编码血清素生物合成的限速酶,在脑内TPHl和TPH2在几个脑区表达相当,如额叶皮层、丘脑、海马、下丘脑和杏仁核。在正常被试群体,有一项研究考察了TPH2基因编号为rsl386438的SNP,结果发现该位点的多态性与IGT表现无关(Stoltenberg&Vandever,2010)。而在病理人群中,研究者发现TPHl A218C多态性以及TPH2编号为rslll8997的SNP多态性都与自杀倾向者的IGT表现有关(Jollant et al.,2007)。也有研究者发现边缘人格障碍患者(Borderline Personality Disorder,BPD)在IGT中表现较差,而且在患者人群中,TPHl一种单体型(ACGCCG)个体多数是IGT成绩较差的个体(Maurex et al.,2009)。单胺氧化酶A基因(Monoamine Oxidase A,MAOA)位于人类x染色体上,MAOA的活性在不同的脑区有差异,在大脑各个脑区均有广泛分布,主要参与5-HT和DA的降解。MAOA基因研究最多的是启动区一段30bp的VNTR,可出现2-5次的重复,3.5R或4R所编码的酶活性较高(H等位基因);3R或5R等位基因所编码的酶活性较低(L等位基因)。有一项病理研究发现MAOA多态性与自杀倾向者的IGT表现有关,H/H基因型的被试表现较好(Jollant et al.,2007)。
2.2.3 其他基因位点对风险决策个体差异的影响
其他与风险决策个体差异相关的基因(不属于DA或5-HT神经递质系统)主要是脑源性神经营养因子基因(Brain-Derived Neurotrophic Factor,BDNF)。BDNF是一种在神经元损伤后再生修复和防止神经细胞退行性变等多方面发挥重要作用的细胞因子,它主要分布在大脑皮层、髓质、小脑、海马等中枢神经系统。人类BDNF基因位于第11号染色体上,研究得最为广泛的位点是编号为rs6265的SNP(也被称为val66met),有两种等位基因Val和Met,影响BDNF蛋白在细胞内的加工及分泌过程。Kang等人(20101考察了BDNF多态性对IGT的影响,结果发现Met/Met和Val/Met基因型都与IGT后期(风险决策)较低的得分(即他们选择了更多的坏牌)相关,并且这些被试的学习效应也较小;但是BDNF基因型与IGT前期(模糊决策)的得分无关。
2.3 多基因系统分析研究
虽然上述众多研究逐渐从基因水平揭示了风险决策的遗传机制,但是目前的研究还存在两个突出的问题:一是研究结果不一致,二是单个基因往往只有1%左右的效应量,难以解释行为的高遗传性(Plomin,Haworth,&Davis,2009)。对于第一个问题,前面提到被试特征、样本量、行为指标等各个方面都是可能引起研究间差异的因素,要进行严格控制;对于第二个问题,从基因系统的水平进行分析可能是一种解决之道。由于DA、5-HT系统的功能显然是合成、代谢、转运、受体等众多相关基因协同作用的结果,逐个研究这些基因的作用显然无法揭示其全貌。因此我们认为,风险决策可能是由很多基因位点共同影响的,由于每个位点的效应量都很小,导致研究结果难以重复,但是多个位点的效应累积起来就导致了高遗传率,从基因系统的水平能得到更准确可靠的基因效应。为了得到这种基因系统水平上的效应量,首先我们采用方差分析的方法筛查出基因系统里具有显著主效应的位点,然后采用多元回归的方法得到这些位点以及他们之间交互作用的整体贡献率,最后采用蒙特卡罗的方法进行验证。采用这种方法,我们发现饮酒(chen et al.,2012)、人格(Chen et al.,2011)、面孔表情识别(Zhuet al.,2012)、体型指标(chen et al.,2013)的个体差异大约有10~20%可以由DA系統或5-HT系统的基因变异来解释。
由于风险决策跟奖赏系统密切相关,而DA系统负责调控奖赏行为,因此我们考察了DA系统对决策行为的整体贡献。采用IGT,我们计算出每个被试对奖赏的敏感程度,结果发现合成多个DA相关的基因(DDC、DBH和TH)跟被试的奖赏敏感程度密切相关,其中DDC的rsl817074,DBH的rsl611123、rs732833和rs2519154,TH的rs4929966和rs2070762以及他们之间的交互作用解释了7%的奖赏敏感程度变异,进一步的蒙特卡罗检验表明该结果非常显著(p=0.009)。这个结果初步表明DA系统相关的基因在一定程度上决定了人们的奖赏依赖水平,从而进一步影响人的决策。
3、遗传环境交互对风险决策个体差异的影响
前文从双生子研究和分子遗传学研究两个方面介绍了遗传因素对风险决策个体差异的影响,但遗传并不是影响风险决策个体差异的唯一因素。环境也是影响风险决策个体差异的一个重要因素。缺乏对环境因素的考量以及遗传与环境交互作用的考量可能是前人基因相关研究所得结果不一致的一个根本原因。
环境因素又分为风险环境因素和保护环境因素。比如,压力事件就属于风险环境因素之一,是众多精神疾病的风险因子(Van Winkel。Stefanis,&Myin-Germeys,2008),并且与多种成瘾行为相关(如吸烟、酗酒、或者不健康饮食等等)(Starcke&Brand,2012)。采用横断(Johns et al.,2004)和纵向研究(Wiles et al.,2006)的思路,研究者们还发现压力事件与自评的精神症状体验相关。在决策方面也有类似的研究,比如研究发现压力事件与正常被试的奖赏敏感性成正比,而与其惩罚敏感性呈反比(starcke&Brand,20 12);也有研究发现压力事件与成瘾者的药物渴求和戒断后的复吸有关(Sinha,2001,2008;Sinha,Catapano,&OMalley1999)。
而另一方面,亲子关系一般被认为是多种行为的保护环境因素的代表。比如,研究发现亲子关系能有效预测学业成就和后期的社会交往(Chen,Liu,&Li,2000),并且良好的亲子关系与较少的青少年问题行为相关,也与较低的抑郁水平相关(Beam,Gil-Rivas,Greenberger,&Chen,2002)。决策研究中,亲子关系也是一个重要的话题。比如Xiao等人(2011)采用纵向追踪发现儿童期亲子关系较好的儿童,其1年后的IGT测验的得分增长越多。
要探讨遗传和环境各自对行为的影响,基因环境的交互作用研究将是一个突破口。基因与环境交互是指环境变量的作用在不同的基因型被试上不同。基因一环境交互的两项先驱研究当属于Caspi等人在Science上发表的两篇论文(caspi etal.,2002;Caspi et al.,2003)。他们的第一篇研究论文报告了儿童虐待和MAOA基因的交互作用,与幼时受虐待且携带编码高水平MAOA基因型的儿童相比,那些幼时受到虐待并且携带编码低水平MAOA基因型的儿童的反社会行为几乎是前者的两倍(caspi et al.,2002)。他们的第二篇研究论文报告了5-HTTLPR基因多态性和应激刺激的交互作用对抑郁形成的影响(caspi et al.,2003),结果发现与l/l基因型的被试相比,在相同数量生活压力事件中,s携带者贻患抑郁症的概率是前者的2.5倍。
在风险决策研究领域Stoltenberg等人率先在一项研究中同时报告了基因与环境对风险决策能力的影响(stoltenberg,Lehmann,Anderson,Nag,&Anagnopoulos,2011)。该研究对391名健康高加索裔被试施测了IGT,发现5-HTTLPR l/l基因型被试在IGT前20个选择时倾向于选更多的风险牌。同时,该研究还发现那些报告了较高儿童期创伤的被试,其IGT后期的得分较低(stoltenberg et al.,2011)。虽然这项研究同时报告了基因与环境各自对风险决策个体差异的影响,但却并没有报告基因与环境的交互作用。我们最近的一篇研究首次考察了风险决策中的基因环境交互作用(He et al.,2012)。该研究施测了超过500名大学生,对被试施测了IGT任务,分析了其COMT基因型。在环境变量方面,该研究不仅评估了风险性环境因素——压力事件,还评估了保护性环境因素——亲子关系。研究结果发现COMT基因与两项环境指标都存在交互作用:压力事件倾向于只影响Met携带者,使其对奖赏的敏感程度升高,从而得到较差的IGT得分;而亲子关系则倾向于只影响Val/Val基因型被试,使得其IGT得分更高(图3)。这个结果说明,遗传与环境将以一种交互作用的方式来影响风险决策行为。
4、脑结构和功能对基因影响风险决策个体差异的中介作用
脑是思维的器官,是人类众多行为的神经基础。新近发展起来的磁共振脑成像(functionalMagnetic Resonance Imaging,fMRI)技术可以使我们直接观察大脑的活动,获得实时动态的大脑活动数据。该项技术的进展和逐渐使用使得研究者可以考察大脑的结构和功能——做为决策能力的内表型(endophenotype)——是如何受基因和环境影响的。
首先,研究者观察到不同COMT基因型的个体在风险决策任务中大脑激活的差别。比如。Boettiger等人(2007)的研究发现COMT Vall58Met多态性Fal/Fal基因型的被试更倾向于选择即时奖赏,也与更强的额一顶叶区域激活有关。Yacubian等人(2007)则发现Met/Met基因型与较强的前额叶皮层(Prefrontal Cortex,PFC)和腹侧纹状皮层(Ventral Striatum)激活有关。Dreher等人(2009)发现COMT基因型与奖赏预期阶段的腹侧纹状皮层和外侧前额叶皮层(lateral PFC)激活相关,与奖赏体验阶段的眶额皮层(Orbitofrontal Cortex,OFC)激活相关。Met~Met基因型的被试表现出最高的激活。Camara等人(2010)采用一个赌博任务,虽然在行为上沒有发现COMT的效应,但是发现COMT基因型能够调节奖赏正负(但不调节奖赏的大小)引起的腹侧纹状皮层、扣带回前部(Anterior Cingulate Cortex,ACC)和顶下皮层的激活。其中最大的激活差异在Val/Val基因型被试。该研究还发现DRD4启动区,521 C/T多态性能够调节奖赏大小(但不调节奖赏的正负)引起的脑岛前部和ACC的激活。
其次,Yacubian等人(2007)除了考察了前文所述的COMT与风险决策大脑激活的关系之外,还考察了DAT基因与风险决策大脑激活的关系。结果没有发现DAT基因型对前额叶激活的影响,但是他们发现DAT基因与COMT有交互作用,该交互作用主要体现在左侧纹状皮层的激活上。而前面也提到的Dreher等人(2009)的研究既发现了DAT的主效应,还发现了DAT与COMT基因的交互作用。他们发现DAT基因的主效应主要体现在奖赏预期阶段的尾状核和腹侧纹状皮层的激活上,也体现在奖赏体验阶段外侧PFC的激活上,9R携带者表现出较高的激活强度。而COMT与DAT的交互作用则主要体现在奖赏预期阶段的腹侧纹状皮层和外侧PFC的激活上,以及奖赏体验阶段的外侧PFC和OFC的激活上。
最后,研究者对五羟色胺系统相关基因也进行了类似的研究。前人的研究已经发现5-HTTLPR多态性与杏仁核(Hariri et al.,2005;Hariri&Holmes,2006;Hariri et al.,2002;Hariri,Tessitore,Malay,Fera,&Weinberger,2002;Labus et al.,2008;Munafo et al.,2008)、腹内侧前额叶(Heinz etal.,2005;Labus et al.,2008)以及脑岛的激活相关(Labus et al.,2008)。5-HTTLPR基因多态性不仅影响这些决策相关脑区的激活强度,还影响他们之间的功能联结。比如,Roiser等人(2009)采用fMRI技术发现,5-HTTLPR基因多态性s/s被试表现出较高的杏仁核和ACC激活,同时他们还发现s/s被试的前额一杏仁核通路的联结较弱。这个结果支持了情绪在风险决策加工中的重要作用,而这个作用很可能是受杏仁核调节的。
5、总结与展望
心理学家试图分离遗传与环境分别对风险决策行为个体差异的影响。一方面,一些研究者通过双生子或者分子遗传学的研究来考察遗传对风险决策行为个体差异的影响;另一方面,另一些研究者则通过考察环境或者基因,环境交互作用来考察环境和遗传各自对风险决策行为个体差异的贡献。而随着脑成像技术的兴起,更多的研究者开始着眼于考察遗传与环境各自对风险决策行为的内表型——大脑结构和功能的影响。
总体上讲,无论是双生子研究还是分子遗传学的研究,也无论是研究得较多的基因(如COMT和SLC6A),还是研究得相对较少的基因(如DAT、TPHl、BDNF等),这些研究都表明遗传因素(基因)在风险决策加工中有重要作用,可以在一定程度上解释风险决策的个体差异。多巴胺和五羟色胺相关的基因在个体风险决策中尤为重要,不仅仅表现在不同基因型(或者基因型组合)的被试对模糊和风险的偏好不同,并且这种基因型的差别可能还伴随着大脑激活模式的差别,主要涉及的脑区有纹状体、杏仁核、ACC和广大的前额叶皮层(包括VMPFC,外侧PFC和OFC等)。虽然这些研究已经揭示了遗传对风险决策的影响,但对其具体机制的了解还相对较少。一方面,人类有2万多个基因,而每个基因都可能有众多变异位点(SNP或者VNTR或其它结构变异),目前所发现的基因与风险决策的关系仅仅是其中很小的一部分,后续研究需要继续探讨究竟有多少基因与风险决策相关,而又有多少基因与风险决策无关。另一方面,虽然目前也找出了诸如DA和5-HT相关的数个基因与风险决策能力相关,但对这些基因如何影响风险决策能力的问题还没有明确的答案,后续的研究应该着眼于揭开基因对风险决策影响的机制。
最后,环境对风险决策能力的影响常常采用基因一环境交互的方式来进行研究,这是因为,基因之间以及基因与环境之间常常具有相互的影响,表现为基因,基因交互作用或基因,环境交互作用。比如,前面提到的一部分研究都没有发现基因的主效应。这都可能是因为该基因与其他基因或环境之间产生了交互作用,进而影响风险决策行为。后续的研究应该更深入探讨基因-基因、基因-环境之间的交互作用对风险决策的影响。最后,基因和环境都更有可能是通过影响风险决策行为的内表型——决策相关脑区的结构和功能来起作用,因此,今后的研究更应该结合fMRI来考察基因和环境对风险决策加工相关脑区的结构和功能的影响。这样的研究不仅有助于深入揭示其背后的生理机制,更有助于开发出预防和治疗具有决策障碍的精神疾病的新方法。
猜你喜欢
区域治理(2022年40期)2022-11-27
疯狂英语·新读写(2021年10期)2021-12-07
中老年保健(2021年12期)2021-08-24
中国传媒大学学报(自然科学版)(2021年1期)2021-06-09
中国生殖健康(2020年6期)2020-02-01
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
奥秘(2019年8期)2019-08-28
中国生殖健康(2019年11期)2019-01-07
