
光周期与植物开花
2014-04-26 09:41:44白变霞
长治学院学报 2014年5期
王 玺,白变霞
(长治学院 生物科学与技术系,山西 长治 046011)
在千百万年的进化过程当中,植物形成了自己独特的机制从而适应周围的环境。植物能够整合分析各种外部环境和季节信息,进而调节自身的基因表达和外部形态,从而达到适应环境的目的。在这些外部环境中,光环境信号显得尤为重要。没有光,植物将无法正常生长。植物可以感知光谱质量、强度、方向以及光照时间,进而根据不同的光照环境来对自身进行调节[1]。
1 植物对光的感受
每个光受体的光谱敏感性,依赖于其发色团吸收不同波长的能力,即依赖发色团的吸收谱(absorption spectrum)。吸收光之后,光受体蛋白可以调控下游的信号传递[2]。
光敏色素最早是从光可逆性(photoreversibility)现象中发现的[3]。在所有的绿色植物和至少某些蓝细菌中都有光敏色素。光敏色素为可溶性蛋白,主要以同源二聚体的形式存在,每个亚基与其自身的生色团结合。光敏色素具有两种能够相互转变的构象,即接收红光的非活性形式(Pr)和接收远红光的活性形式(Pfr)。Pr形式的光敏色素接受到红光照射后就转变成Pfr形式[4]。拟南芥有五个光敏色素蛋白基因,分别为PHYA、PHYB、PHYC、PHYD和PHYE。这些基因编码的光敏色素可以分为两组。光敏色素A 为光不稳定型光敏色素(light-labile phytochrome),是经过长时间的黑暗处理后或者浸湿的种子和幼苗当中的主要的光敏色素。光敏色素B-E为光稳定型光敏色素(light-stable phytochrome),是光照条件下生长的植物中主要的光敏色素类型[5]。
隐花色素,是专为蓝光和UV光受体而起的,既反映出在鉴定这两个受体的过程中的困难度,又反映出这种受体对“隐花植物”(非种子植物的旧称)的重要性。近几年的研究表明,多数对蓝光和UV-A波长的应答是由两个相关的光受体调控的。并将其称为:隐花色素1(CRY1)和隐花色素2(CRY2)。CRY1和CRY2分别由CRY1和CRY2基因所编码。隐花色素为可溶性蛋白,位于核中,在所有的植物器官中都有。每种隐花色素都包含黄素和蝶呤两个生色团[6]。
拟南芥中有一个蓝光/UV-A光受体,参与到了植物的向光性反应。也就是对单向光有应答,产生弯曲。这个光受体由NONPHOTOTROPICHYPOCOTYL 1(NPH1)基因所编码,该基因的缺失将会导致植物丧失向光性[7、8]。NPH1编码一个质膜相关的蛋白激酶,是一类N端具有生色团结合结构域的丝/苏氨酸蛋白激酶(Ser/Thr protein kinase),能够结合两个黄素单核苷酸(Flavin mononuleotide,FMN)分子[9]。向光色素介导的光信号传递主要是由级联反应(Cascade)或Ca2+第二信使途径逐级向下传递的[10]。
2 光周期与植物开花
控制植物开花时间的信号途径至少有4条,分别是:光周期途径、春化途径、自主途径和赤霉素途径[11、12]。而光周期途径是影响植物开花的一个重要因素,是目前相关研究中的热点。
Tournois(1914)and Klebs(1918)第一次提出了光照时间的长短可以影响植物的开花,光周期诱导的植物开花的概念是在1920年首次被Ganner和Allardi提出的[13]。比如,短日照植物(short-dayplants)在夜晚时长超过一定临界值时就会导致开花。长日照植物(long-day plants)在白天时间较长的时候会导致植物开花。还有一种是介于长日照和短日照之间的植物。
1934年,Knott提出了一个非常重要的观点,那就是,日照时间的长短是被叶子所感知的。但是因为开花是发生在植物的茎顶端分生组织(SAM,shoot apical meristem),因此,必然有一种信号可以从叶子传递到SAM,这种信号即是被人们称为“成花素”的蛋白[14]。
在二十世纪30年代中期,科学家们做了大量的工作去分离成花素,但是当时大部分的研究集中在对叶子的提取分析,试图在成花素的应答途径上建立一种生物学分析。但是经历了二十多年都没有成功,甚至有人将成花素称为“圣杯”[15],足以表明当时在成花素的分离过程当中遇到的种种困难。直到遗传学和分子生物学的方法手段被应用到这个领域之后,“圣杯”的神秘面纱才逐渐被揭开。并且需要强调的是这个问题被清楚的定义是通过经典的生理学研究实现的[16、17]。
拟南芥是一种长日照植物(long-day plants),它在长日照下会更迅速的开花,在短日照下开花会延迟[18]。在拟南芥中,一般以莲座叶的数目多少来判断开花的早晚[19],而在其他植物中则不一定是按照叶子的数目来衡量,有的是按照天数来计算开花的时间[20]。
在拟南芥开花时间的研究中,有两类典型的晚花突变体被发现[19、21],只在长日照下,而不在短日照下晚花的突变体,被认为是损害了植物日照长度的感知途径。那些在长日照和短日照下均表现出晚花表型的突变体,则被认为是影响了组成型诱导的开花途径,后来被称为“自主开花途径”[22]。研究还发现,CO、GI和FT在长日照下参与到起始开花的过程中[19]。
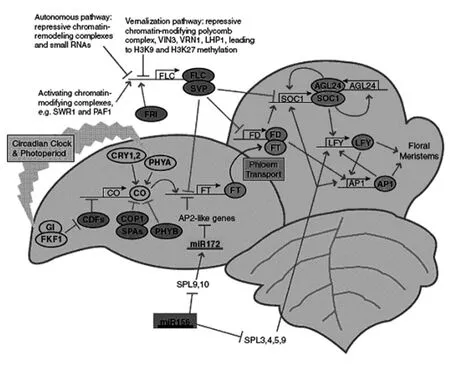
图1 诱导开花的几种信号途径示意图
到目前为止,FT作为一种成花素已经是毋庸置疑的事实,CO和GI在激活FT的表达中起着非常关键的作用。FT是一种有效的开花诱导因子,是一种类似于RAF激酶抑制子的一类小蛋白[23、24]。CO是一个锌指蛋白,可能作为一种转录因子参与到成花诱导的过程中[25]。GI是参与到昼夜节律钟功能的一种较大的植物特异性蛋白[26、27]。
CO的表达能够被昼夜节律钟所影响,并且对于在长日照条件下诱导FT的表达是十分必要的[28、29]。实际上,CO的唯一作用可能是在光周期的诱导下,诱导FT的表达(另外一个可能是,诱导一种FT的相关蛋白TWIN SISTER OF FT)TSF[30、31]。
第一次发现并证明CO可以在叶子中激发一种信号(FT)移动至SAM是源于在拟南芥中异位表达CO,当在叶子的韧皮部伴胞细胞中异位表达CO,但在SAM中没有表达CO的情况下,能够使开花正常进行[32、33]。另一方面,在叶子和SAM表达FT基因,都可以导致开花,且这种开花的诱导是不需要CO的活性的[32]。
FT在韧皮部和叶子中均是以单体的形式存在的[33],但是遗传学和分子学的手段发现,在分生组织中FT是与FD(一种bZIP家族的转录因子)相结合形成聚合物的形式来发挥作用[34、35],并且,构建FT::GFP融合蛋白的方法进一步证明了:FT是从幼嫩叶子中长途运输至SAM[36-38]。
在CO的转录,光周期和昼夜节律之间的相互作用的研究上,人们已经花费了大量的精力[39-42]。CDFs(CYCLING DOF FACTOR)呈现出了一种昼夜戒律的循环,并且可以与CO的启动子相结合,从而节律性的抑制CO的表达。而CDFs的含量还受到一种F-box(FLAVIN-BINDINGKELCH REPEAT F-BOXPROTEIN1)蛋白简称FKF1的调控,FKF1为一种蓝光受体,可以参与到CDFs蛋白的降解过程。时钟蛋白GI可与FKF1相互作用,并使FKF1变得更加稳定,然后在适当的条件下,起始CDFs的降解,使CO得以表达。并且这种情况在植物界中是保守的[42-45]。
光周期和光含量也可通过稳定CO蛋白的方式来影响FT的表达。CRY1和CRY2是蓝光受体,PHYA是一种红/远红光受体,这三种蛋白可以在光周期白天阶段的末尾起到稳定CO蛋白的作用。但是,另一个红/远红光受体PHYB能够促使CO蛋白的循环降解[67]。参与到CO降解过程的还有COP1以及与COP1相互作用的SPA蛋白家族成员[46-49]。
当FT被长途运输到茎顶端分生组织(SAM)之后,需要与FD相互作用才能起始开花。FT/FD复合体可以激活下游的花分生组织本体基因的表达。如,AP1[30-33]以及其他的促使开花的基因,如SOC1[50、51]。这些基因可以在SAM的拱侧翼特定细胞层中起始叶元基的产生。另外LFY可以诱导AP1的表达,并且LFY可以起始花原基向花分生组织的转化。另外赤霉素信号途径也可激活AP1的表达[52、53]。图1中所示为其他诱导开花的途径。
[1]Chory J. Light modulation of vegetative development[J]. Plant Cell. 1997, 9:1225-1234.
[2]Von Arnim A, Deng,X-W. Light control of seeding development[J]. Annual Review of plant physiology and Plant Molecular Biology. 1996,47:215-244.
[3]Nagy F, Schafer E. Nuclear and cytosolic events of light-induced phytochrome-related signaling in higher plants [J]. EMBO Journal. 2000, 19:157-163.
[4]Quail P.H. Phytochrome photosensory signalling networks [J]. Nat Rev Mol Cell Biol. 2002, 3:85-93.
[5]Neff,M.M., Fankhauser, C Chory, J. Light:an indicator of time and place[J]. Gene and Development. 2000, 14:157-271.
[6]Cashmore,A.R. Jarillo,J.A Wu,Y-J Liu,D. Cryptochromes: blue light receptors for plants and animals[J]. Science. 1999, 284:760-765.
[7]Lin,C. Plant blue-light receptors [J]. Trends in Plant Science. 2000, 5:337-342.
[8]Christie,J.M.,Reymond,P.Powell,G.K. Arabidopsis NPH1: a flavoprotein with the properties of a photoreceptor for phototropism [J]. Science.1998, 282:1698-1701.
[9]Christie J.M. and Briggs W.R. Blue light sensing in higher plants[J]. J Biol Chem. 2001, 276:11457-11460.
[10]Harad A. and Shimazaki K. Phototropins and blue light-dependent calcium signaling in higher p lants [J]. Photochemistry and Photobiology.2007,83:102-111.
[11]Mouradov A,Cremer F,Coupland G.Control of flowering time:interacting pathways as a basis for diversity [J]. Plant Cell. 2002,14 (Suppl):S111-S130.
[12]Simpson G,Dean C.Arabidopsis the Rosetta stone of flowering time?[J]. Science. 2002, 296(5566):285-289.
[13]Richard Amasino. Seasonal and developmental timing of flowering [J]. The Plant Journal.2010, 61:1001-1013.
[14]Chailakhyan, M.K. New facts in support of the hormonal theory of plant development [J]. C.R. (Dokl.) Acad. Sci. USSR.1936, 13:79-83.
[15]Zeevaart, J.A. Leaf-produced floral signals[J].Curr. Opin. Plant. Biol. 2008, 11:541-547.
[16]Lang, A. Physiology of flower initiation Ⅰn Encyclopedia of Plant Physiology [J]. (Ruhland,W., ed). Berlin: Springer-Verlag. 1965, pp:1371-1536.
[17]Yanovsky, M.J. and Kay, S.A. Molecular basis of seasonal time measurement in Arabidopsis[J].Nature. 2002, 419:308-312.
[18]Gregory, F.G. and Hussey, G.G. Photoperiodic responses of Arabidopsis thaliana[J]. Proc. Linn.Soc. Lond. 1953, 164:137-139.
[19]Koornneef, M., Hanhart, C.J. and van der Veen, J.H. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana[J]. Mol. Gen. Genet. 1991, 229:57-66.
[20]Buckler, E.S., Holland, J.B., Bradbury, P.J. The genetic architecture of maize flowering time[J].Science. 2009, 325:714-718.
[21]Redei, G.P. Supervital mutants in Arabidopsis[J]. Genetics. 1962, 47: 443-460.
[22]Koornneef, M., Hanhart, C.J. and van der Veen, J.H. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana[J]. Mol. Gen. Genet. 1991, 229:57-66.
[23]Kardailsky, Ⅰ., Shukla, V.K., Ahn, J.H., Dagenais, N., Christensen, S.K.,Nguyen, J.T., Chory,J., Harrison, M.J. and Weigel, D. Activation tagging of the floral inducer FT[J]. Science.
[24]Kobayashi, Y., Kaya, H., Goto, K., Ⅰwabuchi,M. and Araki, T. A pair of related genes with antagonistic roles in mediating flowering signals[J]. Science. 1999, 286:1960-1962.
[25]Putterill, J., Robson, F., Lee, K., Simon, R. and Coupland, G. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors[J]. Cell. 1995, 80:847-857.
[26]Fowler, S. Lee, K., Onouchi, H., Samach, A.,Richardson, K., Morris, B., Coupland, G. and Putterill, J. GⅠGANTEA: a circadian clockcontrolled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains[J]. EMBO J. 1999, 18:4679-4688.
[27]Park, D.H., Somers, D.E., Kim, Y.S., Choy, Y.H., Lim, H.K., Soh, M.S., Kim, H.J.,Kay, S.A.and Nam, H.G. Control of circadian rhythms and photoperiodic flowering by the Arabidopsis GⅠGANTEA gene [J]. Science. 1999, 285:1579-1582.
[28]Suarez-Lopez, P., Wheatley, K., Robson, F.,Onouchi, H., Valverde, F. and Coupland, G.CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis[J]. Nature. 2001, 410:1116-1120.
[29]Yanovsky, M.J. and Kay, S.A. Molecular basis of seasonal time measurement in Arabidopsis[J].Nature. 2002,419:308-312.
[30]Wigge, P.A., Kim, M.C., Jaeger, K.E., Busch,W., Schmid, M., Lohmann, J.U. and Weigel, D.Ⅰntegration of spatial and temporal information during floral induction in Arabidopsis [J]. Science. 2005, 309:1056-1059.
[31]Yamaguchi, A., Kobayashi, Y., Goto, K., Abe,M. and Araki, T. TWⅠN SⅠSTER OF FT (TSF)acts as a floral pathway integrator redundantly with FT [J]. Plant Cell Physiol. 2005, 46:1175-1189.
[32]An, H., Roussot, C., Suarez-Lopez, P. et al.CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis [J]. Development.2004,131:3615-3626.
[33]Ayre, B.G. and Turgeon, R. Graft transmission of a floral stimulant derived from CONSTANS[J]. Plant Physiol. 2004,135:2271-2278.
[34]Takada, S. and Goto, K. Terminal flower2, an Arabidopsis homolog ofheterochromatin protein1, counteracts the activation of flowering locus T by constans in the vascular tissues of leaves to regulate flowering time [J]. Plant Cell.2003,15:2856-2865.
[35]Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A., Ⅰkeda, Y.,Ⅰchinoki, H.,Notaguchi, M., Goto, K. and Araki, T. FD a bZⅠP protein mediating signals from the floral pathway integrator FT at the shoot apex[J].Science. 2005, 309:1052-1056.
[36]Corbesier, L., Vincent, C., Jang, S. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis[J].Science.2007, 316:1030-1033.
[37]Jaeger, K.E. and Wigge, P.A. FT protein acts as a long-range signal in Arabidopsis[J]. Curr. Biol. 2007, 17:1050-1054.
[38]Tamaki, S., Matsuo, S., Wong, H.L., Yokoi, S.and Shimamoto, K. Hd3 a protein is a mobile flowering signal in rice[J]. Science. 2007, 316:1033-1036.
[39]Ⅰmaizumi, T., Schultz, T.F., Harmon, F.G., Ho,L.A. and Kay, S.A. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis [J]. Science. 2005,309:293-297.
[40]Sawa, M., Nusinow, D.A., Kay, S.A. and Ⅰ-maizumi, T. FKF1 and GⅠGANTEA complex formation is required for day-length measurement in Arabidopsis [J]. Science. 2007, 318:261-265.
[41]Fornara, F. and Coupland, G. Plant phase transitions make a SPLash [J]. Cell. 2009, 138:625-627.
[42]Ⅰmaizumi, T. Arabidopsis circadian clock and photoperiodism: time to think about location[J]. Curr. Opin. Plant Biol. 2009, 13:83-89.
[43]Harmer, S.L. The circadian system in higher plants [J]. Annu. Rev. Plant Biol. 2009, 60:357-377.
[44]Mas, P. and Yanovsky, M.J. Time for circadian rhythms: plants get synchronized[J]. Curr. Opin.Plant Biol. 2009, 12:574-579.
[45]McClung, C.R. Circadian rhythms. Linking the loops[J]. Science. 2009, 323:1440-1441.
[46]Valverde, F., Mouradov, A., Soppe, W.,Ravenscroft, D., Samach, A. and Coupland, G.Photoreceptor regulation of CONSTANS protein in photoperiodic flowering [J]. Science.2004, 303:1003-1006.
[47]Laubinger, S., Marchal, V., Le Gourrierec, J.,Wenkel, S., Adrian, J., Jang, S., Kulajta, C.,Braun, H., Coupland, G. and Hoecker, U. Arabidopsis SPA proteins regulate photoperiodic flowering and interact with the floral inducer CONSTANS to regulate its stability[J]. Development. 2006, 133:3213-3222.
[48]Jang, S., Marchal, V., Panigrahi, K.C., Wenkel,S., Soppe, W., Deng, X.W., Valverde, F. and Coupland, G. Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response [J]. EMBO J. 2008, 27:1277-1288.
[49]Liu, L.J., Zhang, Y.C., Li, Q.H., Sang, Y.,Mao, J., Lian, H.L., Wang, L. and Yang, H.Q.COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis [J]. Plant Cell. 2008b,20:292-306.
[50]Michaels, S.D., Himelblau, E., Kim, S.Y.,Schomburg, F.M. and Amasino, R.M. Ⅰntegration of flowering signals in winter-annual Arabidopsis[J]. Plant Physiol. 2005, 137:149-156.
[51]Yoo, S.K., Chung, K.S., Kim, J., Lee, J.H.,Hong, S.M., Yoo, S.J., Yoo, S.Y., Lee, J.S. and Ahn, J.H. CONSTANS activates SUPPRESSOR OF OVEREXPRESSⅠON OF CONSTANS 1 through FLOWERⅠNG LOCUS T to promote flowering in Arabidopsis [J]. Plant Physiol. 2005, 139:770-778.
[52]Blazquez, M.A., Green, R., Nilsson, O., Sussman, M.R. and Weigel, D. Gibberellins promote flowering of Arabidopsis by activating the LEAFY promoter [J]. Plant Cell. 1998, 10:791-800.
[53]Liljegren, S.J., Gustafson-Brown, C., Pinyopich, A., Ditta, G.S. and Yanofsky, M.F. Ⅰnteractions among APETALA1, LEAFY, and TERMⅠNAL FLOWER1 specify meristem fate[J]. Plant Cell. 1999, 11:1007-1018.
猜你喜欢
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
山东陶瓷(2021年5期)2022-01-17 02:35:46
金桥(2021年9期)2021-11-02 05:54:52
金桥(2021年9期)2021-11-02 05:54:30
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
星星·散文诗(2017年2期)2017-07-05 11:34:16
小布老虎(2016年18期)2016-12-01 05:47:41
光学精密工程(2016年5期)2016-11-07 09:06:00
