
渤海浮游植物生物量季节变化的模拟研究*
2014-04-17 07:47刘菲菲陈学恩
中国海洋大学学报(自然科学版) 2014年2期
刘菲菲,陈学恩
(中国海洋大学海洋环境学院,山东 青岛266100)
海洋浮游植物是海水中有机物的初级生产者和能量的转换者,是海洋食物链中更高营养级的食物来源,其生物量的变化直接影响着整个海洋食物链的物质循环和能量转换,从而影响渔业资源的数量,在海洋生态系统中具有重要作用[1-3]。影响浮游植物生物量变化的因素很多,包括营养盐、温度、光照等理化因子以及不同的生物过程。近几十年来,由于气候变化、环渤海工农业及养殖业的发展,渤海生态系统的理化环境发生了显著的变化[4],为了更好地保护渤海生态环境不受破坏,探究渤海生态系统动力学的特性,尤其是浮游植物生物量的季节变化特征及影响机制就显得尤为重要[5]。
迄今为止,我国在渤海已经进行了多次大规模的生态调查[6],1958—1959年的全国范围海洋普查、1982—1983年及1992—1993年的渤海渔业和生物资源调查[1,7-10]、1984—1985年的渤黄东海4季度月调查[11]、1998—1999年的中德合作项目“渤海生态系统的观测与模拟”[5,12-13]以及1998年9月~1999年9月在长岛的逐月监测[6]等渤海生态系统调查研究为渤海生态系统尤其是浮游植物生物量和初级生产的研究奠定了基础。然而,就观测资料的时空连续性来说,目前的数据资料仍然不能满足对渤海生态系统动力机制的全面认识,幸运的是,数值模型以其高时空分辨率和可重复性的优势成为渤海生态系统机制研究的重要手段[14]。目前,对渤海生态系统的模型研究主要集中在三维水动力-生态耦合模型上,用来模拟初级生产力、浮游植物生物量的时空分布特征[14-18]以及营养盐收支[19-20]。另外,Zhang等[21]根据1998—1999年的中德合作项目观测结果,在渤海建立了1个简单的海洋水体-底层多箱模型,研究了渤海生态系统的季节变化以及系统内部的物质、能流交换情况。与此同时,重要物理过程对渤海生态变量空间分布的影响机制也得到一定的研究。王强[22]利用三维耦合模式研究了渤海环流的季节变化对浮游植物空间分布的影响,指出海峡处的海水交换、黄河入流以及辽东湾东侧沿岸流是叶绿素浓度空间分布不一致的原因。Zhao和 Wei等[23]采用三维初级生产模型对渤海1982年浮游植物生物量的分布进行了模拟,指出湍流混合的不均匀性是造成浮游植物高生产区呈斑块分布的原因之一;水平对流通过改变营养基础的分布造成海域生物量的显著变化,从而降低了海域的生物量;河流输入只引起局部海域系统的调整。然而,目前对于影响浮游植物生物量季节变化的因素却鲜有详细的研究。为此,本文回归到1个包含4种生态变量的箱式生态模型,忽略对流、扩散等物理过程对生态变量空间分布的影响,旨在详细探讨渤海浮游植物生物量的季节变化特征及影响因素,同时对模型进行了参数敏感性分析,找出影响渤海生态系统的主要敏感参数。另外,针对外界理化强迫场的改变进行了一系列的数值试验,详细探讨了重要理化因子对渤海浮游植物生物量季节变化的影响,为更好地治理渤海生态系统提供依据。
1 模型介绍
鉴于研究浮游植物生物量季节变化的目的,本文更多地关注浮游植物生物量的时间变化而非空间分布,因此选用了 Tian[24]根据Fasham[25]和Popova[26]的模型修改的包含营养盐(N)、浮游植物(P)、浮游动物(Z)、底碎屑(D)4种生态变量的箱式生态模型,尽可能多地反应真实生物过程,而忽略对流、扩散等物理过程。1982年5月~1983年5月的渤海调查显示,氮为限制性营养盐[27-28],借鉴高会旺等[29]的作法,本文模型中的营养盐仅考虑无机氮,在下文中均以营养盐代指。概念模型如图1所示。
箱式模型考虑了水体与底层界面处的湍流混合引起的物质交换。另外,在底层设定了恒定的营养盐浓度,底层对箱式水体的营养盐补充取决于两者之间的营养盐浓度差,以此来作为水体中营养盐的外源补充[24],此假设的合理性将在文章第四部分进一步讨论。模式具体的生态过程如图1框中所示:浮游植物吸收光、营养盐进行光合作用,部分被浮游动物捕食,部分死亡变成碎屑,部分由于混合作用沉降到底层;浮游动物捕食浮游植物,同时摄取底碎屑进行同化吸收,部分死亡变成底碎屑,部分通过排泄变成营养盐;营养盐的来源有外源输入(水体与底层营养盐浓度差决定的营养盐输入)、浮游动物排泄、底碎屑矿化,汇是浮游植物的吸收及水体与底层界面处的湍流混合;底碎屑的来源有浮游植物死亡、浮游动物死亡、浮游动物捕食浮游植物过程中未被同化的部分等,汇是其矿化作用及浮游动物摄取,同样箱式水体与底层之间的混合作用及自身的沉降使碎屑离开水体进入底层。
模式的物质守衡方程如下:
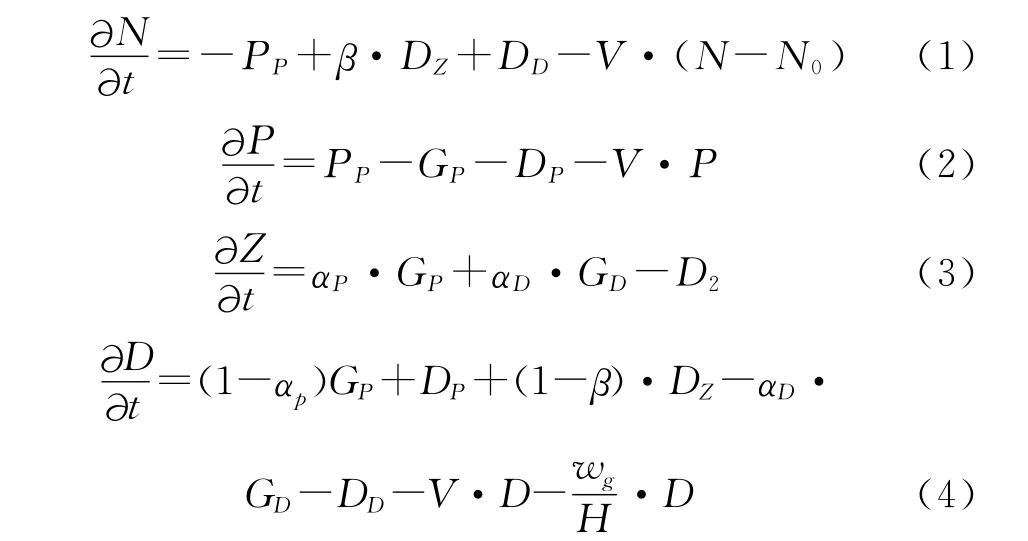
式中各符号的意义及表达式见表1。

表1 生态过程参量表达式Table 1 Meanings and expressions of the biological terms
其中:φ(T)= ekT,代表温度限制函数;φ(N)=代表营养盐影响因子代表浮游植物生长的光限制率;水下光强IZ=I0e-kz考虑浮游植物自身的遮挡作用,详细模式介绍参照文献[24]。
模式中温度场、太阳辐射、云量作为外界强迫给出。温度场取1982年5月~1983年5月的实测场[7,9],太阳辐射根据纬度和时间计算得出;云量取1983年NCEP/CFSR数据,由太阳辐射和云量计算得到海表面光强。由于渤海缺乏足够的观测实验来确定生态模型参数,本文在进行模型参数选取时参照了以往有关渤海生态研究的文献以及在相近海区(如德国的北海)的工作。在此基础上,通过进一步的参数优化确定了本模型的最优参数配置,具体配置见表2。*表示在参照文献的基础上通过参数优化对参数取值进行了一定的调整,其中底层营养盐浓度(N0)用来计算水体与底层的营养盐浓度差,并不能代表实际的底层营养盐浓度,因此无法通过观测或文献给出。本文参照Fasham[25]的做法,将其看做自由参数,通过参数优化给出最优值。模式计算11年,前10年用来使模式稳定,最后1年的结果作为对1983年渤海生物量的模拟结果进行分析。

表2 模型参数配置Table 2 Parameters of the ecosystem model
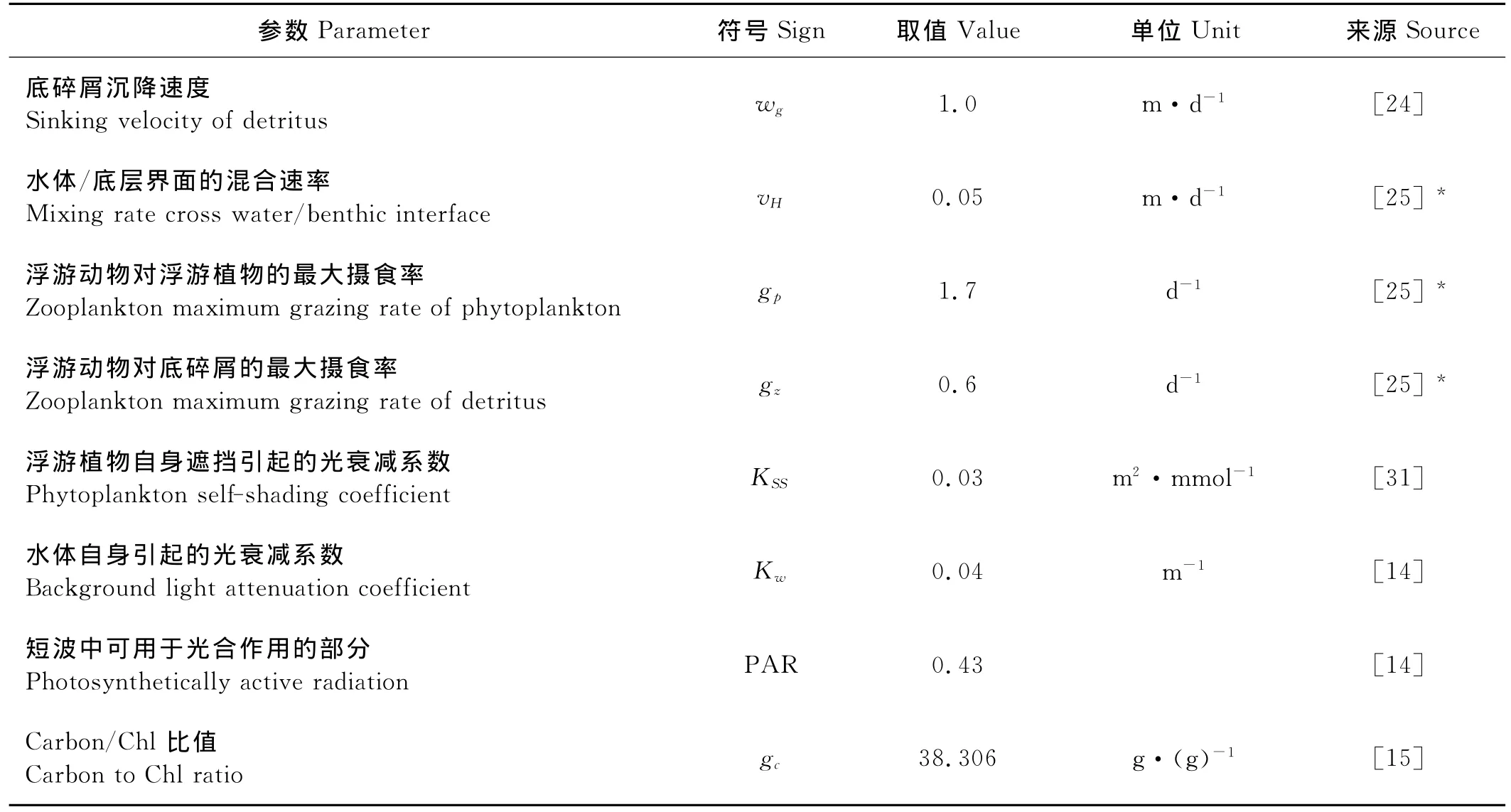
续表2
2 1983年浮游植物生物量、营养盐浓度季节变化的模拟结果
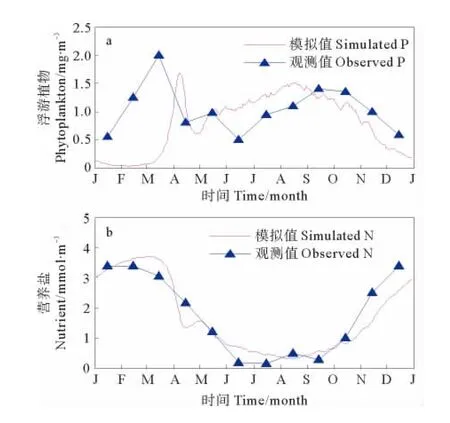
图2 1983年渤海 (a)浮游植物生物量 (b)营养盐浓度季节变化模拟结果(日平均)与观测值(月平均)对比Fig.2 Model results compared with observations:(a)phytoplankton biomass and(b)nutrient concentration
将逐日平均的模拟结果与1982—1983年的月平均观测结果[3]进行对比。由图2可以看出,模拟的渤海浮游植物生物量季节变化趋势与观测结果基本一致,总体来说呈双峰分布,2个峰值分别出现在春季和夏季,2个谷值分别出现在晚春和冬季,其中春季水华峰值大于夏季水华,而冬季浮游植物生物量达最小;营养盐浓度在冬季达到最高,在夏季达到全年的最低值。浮游植物生物量、营养盐浓度的逐月变化规律是:1、2月份浮游植物生物量最低,此时营养盐在水体中积累。从3月份开始,浮游植物生物量快速增加,在4月份浮游植物生物量达到最大值。随着浮游植物的生长,营养盐大量消耗,浮游植物的生长逐渐受到营养盐限制,生物量开始降低,在5月初进入低值期,与此对应,营养盐浓度出现一个小幅回升。从5月中旬开始,由于外界营养盐的补充,浮游植物量开始增加,但增加速度较春季缓慢,在8月中旬第二次达到最大值,较春季峰值稍小。营养盐浓度在8月份达到最低,之后水体中的营养盐开始积累,在冬末春初达到最大浓度值。
与观测结果相比,本文的模拟结果能够合理反映渤海浮游植物生物量和营养盐浓度的季节变化特征,但是,文本模拟的春季水华比实际观测晚半个月,在1、2月份浮游植物生物量与实际观测相差较大,另外,实际观测在6月中旬出现最小值,模拟结果却在5月初出现最小值并在5月中旬就开始增加,而夏季水华也比实际观测提前半个月。与前人的模拟研究相比,本文模拟的春、夏季水华出现时间与魏皓等[17]、赵亮[18]的模拟结果一致,但从模拟的水华强度上来看,文本的模拟结果与实际观测更为接近,能够再现春季水华强度大于夏季水华的季节变化特征;相比刘浩和尹宝树[16]的模拟结果,本文模拟的浮游植物生物量、营养盐浓度季节变化趋势与其模拟结果基本一致,但本文模拟的春季水华持续时间更短,夏季水华持续时间更长。由此可见,模拟结果与实际观测及前人的结果存在一定的差异,考虑可能产生的原因,一是本模型为零维模型,重在强调生物过程的模拟,而忽略了物理过程对于系统的影响。二是模式进行了尽可能的简化,只考虑了一种营养盐、一种浮游植物,而没有细化考虑不同营养盐、浮游植物的生物过程。实际上,生态系统中的各个过程是极为复杂的,以一个简单的箱式模型远不能精确模拟出其中的变化过程。第三,由于观测资料有限,参数优化过程可利用的实测数据不足,造成模式参数配置存在一定的误差。第四,可利用的实测资料精度较低,进行插值后会产生一定的误差。
需要指出的是,本文的模拟并不是为了研究模式误差或是追求模拟结果的精确性,而是希望利用有限的实测数据与不同条件下的模拟结果进行对比,从而更加深入地了解模式中各个生物过程的机理以及不同理化因子的作用。因此,本文认为此模型在渤海的应用是合理的,可以利用此模型进行进一步的参数敏感性分析和数值试验。
3 参数敏感性分析
文本优先选取与浮游植物有关的参数进行敏感性分析,为了比较不同参数的敏感性,仿照Fasham的做法,定义敏感度为[25]:
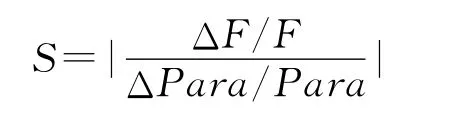
其中:S为参数敏感度;ΔF/F为年均浮游植物生物量(或营养盐浓度)的变化率;ΔPara/Para为参数变化率,本文中设置为50%。各参数敏感性实验结果如表3。
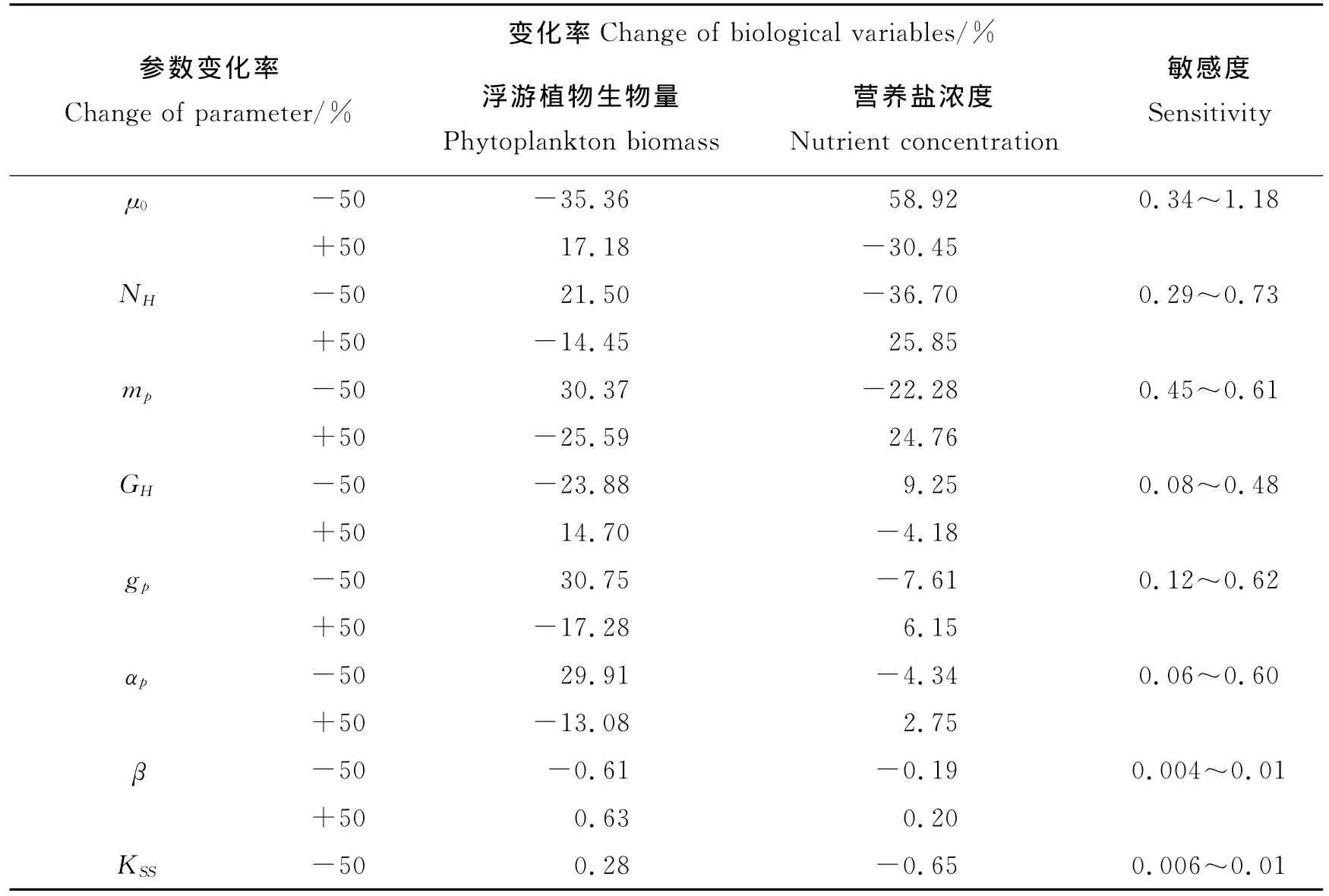
表3 生物参数敏感度Table 3 The sensitivity of the biological parameters
由表3可见,将浮游植物最大生长率(μ0)、浮游植物最大死亡率(mp)、浮游植物吸收营养盐的半饱和系数(NH)改变50%,年均浮游植物生物量变化都大于14%,年均营养盐浓度变化均大于22%;将浮游动物对浮游植物的最大摄食率(gp)、浮游动物捕食浮游植物同化比率(αp)、浮游动物摄食半饱和系数(GH)改变50%,虽然年均营养盐浓度变化小于10%,但年均浮游植物生物量变化都大于13%,说明模式对以上6个参数是敏感的。然而,将浮游动物损耗的再矿化比率(β)和浮游植物遮挡造成的光衰减系数(KSS)改变50%,年均浮游植物生物量和营养盐浓度的变化均不超过1%,因此可以得出结论:模型对参数β、KSS不敏感。
为了更好地研究上述敏感生物参数对模拟结果的影响,本文进一步作出不同参数条件下的浮游植物生物量、营养盐浓度变化曲线如图3,结合表3对参数的敏感性进行定性分析。

图3 不同参数下浮游植物生物量和营养盐浓度的季节变化Fig.3 The seasonal cycles of phytoplankton biomass and nutrient concentration
图3(a)~(c)显示,分别改变参数μ0、mp、NH的大小,浮游植物生物量的季节变化趋势发生了相同的变化。春季,浮游植物生物量和水华发生时间均变化显著,而夏季水华变化很小。另外,这3个参数的变化都改变了营养盐浓度在春季水华期间的变化趋势,但是对夏季水华期间的变化趋势却没有明显的改变,说明这3个参数所对应的生物过程都与浮游植物吸收营养盐进行光合作用有关,是模型最敏感的参数。
图3(d)~(f)中,随着参数gp、αp、GH的改变,虽然年平均浮游植物生物量有明显的变化,但是浮游植物生物量的季节变化趋势并没有显著的改变,春季水华的发生时间不受这3个参数变化的显著影响,只有春季水华的消失阶段及夏季水华期间的浮游植物生物量发生一定的变化。与此同时,营养盐浓度的季节变化几乎不受影响。这是由于以上3个参数所对应的生物过程都跟浮游动物摄食有关,说明浮游动物摄食带来的压力对夏季水华影响显著,而这3个参数是模型次敏感的参数。
4 理化因子对渤海浮游植物生物量季节变化的影响
不考虑海水的运动状态和湍流特征,则生态系统的物理强迫为温度、辐射和透明度,化学强迫为营养盐[12]。为研究不同理化因子对渤海浮游植物生物量季节变化的影响,根据高会旺等[29]的做法,设置数值试验如表4,得到的浮游植物生物量季节变化见图4。
4.1 温度变化对渤海浮游植物生物量季节变化的影响
温度具有相对稳定的特点,因此,实验1中仅考虑10%的变化范围[29]。温度升高和降低10%,浮游植物生物量的大小和变化趋势变化都很小。模式中温度限制公式采用Skogen的公式φ(T)=ekT,当温度增加10%时,在春季水华期间生物量有所降低(见图4(a)),这是由于温度同时影响浮游植物的生长和死亡以及浮游动物对浮游植物的捕食,其共同的效果有可能表现出春季温度的增加反而造成浮游植物生物量减少。反之,温度减小10%,在夏季浮游植物生物量减少,这可能是因为在夏季温度的变化对浮游植物生长的影响要大于对浮游植物死亡及浮游动物捕食的影响。

表4 数值试验设计Table 4 Design of numerical experiments
实验6中(见图4(f)),温度设为常数(年平均值,高于春季水温,低于夏季水温),光照条件为实际情况,夏季水华消失,春季水华仍然出现,只是强度和发生时间均发生一定的变化。相反,若将光照条件设为全年一致而温度变化为实际情况(见图4(g)),春季水华消失而夏季水华明显。通过以上2个试验的比较可以看出,实际的温度强迫是夏季水华发生的控制因素之一。实验6中设定温度为全年的平均值13.74°C,高于渤海春季的水温,此时春季水华发生时间提前(见图4(f)),这与实验1(见图4(a))中增加温度的结果是一致的,说明在春季水华发生初期,在实际的光照条件下,更高的水温有利于浮游植物的生长,虽然也有利于浮游动物的捕食,但对前者的促进作用更大一些,因此会出现更早的水华。综上所述,温度对浮游植物生物量季节变化的影响:春季表现为与辐射、透明度共同作用产生影响,夏季则是浮游植物净生物量增加的主要影响因素。
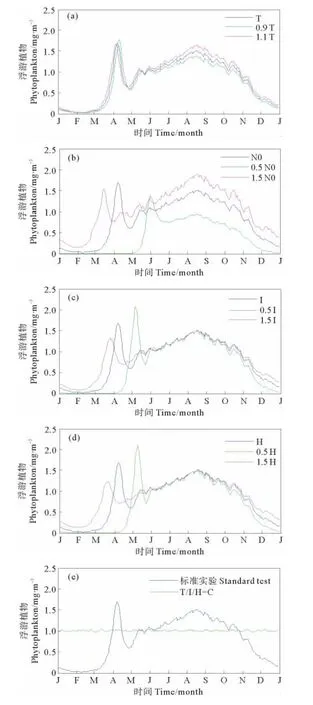

图4 不同物理、化学强迫下浮游植物生物量的季节变化Fig.4 Seasonal cycles of phytoplankton biomass in the Bohai Sea under different physical/chemical forcing conditions
4.2 营养盐变化对渤海浮游植物生物量季节变化的影响
由于模型中假定营养盐的补充是由底层营养盐与水体中营养盐的浓度差决定的,图5刻画了两者营养盐浓度差的季节变化,可以看到浓度差最大出现在夏季,最小出现在春季水华发生之前,这就意味着夏季营养盐的外源补充最丰富,这与实际情况是相符的,因此本文把底层营养盐的补充作为包括河流输入、大气沉降等在内的所有外源补充是合理的。邹立和张经[32]通过现场实验证明渤海大部分地区在春季不受营养盐限制的影响,因此本文将讨论重点放在夏季水华上。实验2(见图4(b))显示高营养盐补充会引起更强的夏季水华,而低营养盐补充会使夏季水华变弱甚至消失,这说明夏季营养盐的补充决定了夏季水华的强度,外源营养盐补充是夏季营养盐的主要来源,其多少决定了夏季水华的强度。

图5 底层与箱式水体的营养盐浓度差的季节变化Fig.5 Seasonal cycle of the nutrient concentration difference between the benthic layer and the water column
4.3 光照条件对渤海浮游植物生物量季节变化的影响
图4(c)~(d)显示,辐射强度和透明度大小的改变对浮游植物生物量的季节变化影响结果基本相同:春季,辐射强度越大,透明度越大,水华产生的时间就越早,但是浮游植物生物量的峰值却减小;夏季,两者对于浮游植物生物量的变化基本没有影响。以上现象很好地说明了在营养丰富的浅海生态系统中,水下光强的变化对于春季浮游植物水华的发生起着极其重要的作用[33-35],春季光的限制程度要比营养盐限制大,尤其是在沿岸海域和河口处。数值试验7当辐射和透明度都为常数时,春季水华不出现(见图4(g)),也证明了光是春季水华的主要限制因子,而夏季浮游植物生物量没有明显变化,说明在夏季光不是浮游植物生物量变化的限制因子。
4.4 物理强迫对渤海浮游植物生物量季节变化的综合作用
试验5的条件为透明度、辐射、温度都为常数,此时春季水华和夏季水华都消失,浮游植物生物量保持一个基本稳定的值(见图4(e)),这说明恒定的物理强迫不可能引起浮游植物生物量的季节变化,实际观测到的浮游植物生物量的季节变化需要温度、太阳辐射、透明度等物理因素的综合强迫。
5 结论
本文通过建立1个包含4种生态变量的箱式生态模型,模拟了渤海1983年浮游植物生物量和营养盐浓度的季节变化,模拟结果与实际观测定性吻合。对模型进行了参数敏感性分析和一系列数值试验,讨论了理化因子对渤海浮游植物生物量季节变化的影响。主要结论如下:
(1)浮游植物生物量的季节变化呈现双峰分布,春季水华强度大于夏季水华;营养盐浓度冬季最大,夏季最小。
(2)模型对浮游植物最大生长率、浮游植物最大死亡率、浮游植物吸收营养盐的半饱和系数最敏感,对浮游动物对浮游植物的最大摄食率、浮游动物捕食浮游植物同化比率、浮游动物摄食半饱和系数次敏感,对浮游动物损耗的再矿化率、浮游植物遮挡造成的光衰减系数不敏感。
(3)理化因子的强迫不仅影响浮游植物生物量的大小,也决定了其季节变化的特征:温度在春季与辐射、透明度共同影响春季水华的发生时间及强度,在夏季是水华发生的主要决定因素;光照是春季水华发生的主要决定因子,对夏季水华没有显著的影响;外源营养盐补充是夏季渤海营养盐的主要来源,其多少决定了夏季水华的强度。
[1]康元德.渤海浮游植物的数量分布和季节变化 [J].海洋水产研究,1991,12:31-44.
[2]刘浩.渤海生态系统关键物理生物过程的数值研究 [D].青岛:中国科学院研究生院 (海洋研究所),2005.
[3]费尊乐,毛兴华,朱明远,等.渤海生产力研究:叶绿素,a初级生产力与渔业资源开发潜力[J].海洋水产研究,1991,12:55-69.
[4]Zhang J,Yu Z G,Raabe T,et al.Dynamics of inorganic nutrient species in the Bohai seawaters [J].Journal of Marine Systems,2004,44(3-4):189-212.
[5]Sündermann J,Feng S Z.Analysis and modeling of the Bohai Sea ecosystem-ajoint German-Chinese study [J].Journal of Marine Systems,2004,44(3-4):127-140.
[6]魏皓,赵亮,于志刚,等.渤海浮游植物生物量时空变化初析[J].青岛海洋大学学报,2003,33(2):173-179.
[7]唐启升,孟田湘.渤海生态环境和生物资源分布图集 [M].青岛:青岛出版社,1997.
[8]费尊乐,毛兴华,朱明远,等.渤海生产力研究——Ⅰ.叶绿素a的分布特征与季节变化 [J].海洋学报,1988,10(1):99-106.
[9]费尊乐,毛兴华,朱明远,等.渤海生产力研究II:初级生产力及潜在渔获量的估算 [J].海洋学报,1988,10(4):481-489.
[10]吕瑞华,夏滨,李宝华,等.渤海水域初级生产力10年间的变化[J].黄渤海海洋,1999,17(3):80-86.
[11]海洋图集编委会.渤海、黄海、东海海洋图集 [M].北京:海洋出版社,1991.
[12]孙军,刘冬艳,柴心玉,等.1998—1999年春秋季渤海中部及其邻近海域叶绿素a浓度及初级生产力估算[J].生态学报,2003,23(3):517-526.
[13]Wei H,Sun J,Moll A,et al.Phytoplankton dynamics in the Bohai Sea—observations and modelling[J].Journal of Marine Systems,2004,44(3):233-251.
[14]刘浩,尹宝树.渤海生态动力过程的模型研究——Ⅰ.模型描述[J].海洋学报,2007,28(6):21-31.
[15]高会旺,王强.1999年渤海浮游植物生物量的数值模拟 [J].中国海洋大学学报:自然科学版,2004,34(5):867-873.
[16]刘浩,尹宝树.渤海生态动力过程的模型研究 Ⅱ.营养盐以及叶绿素a的季节变化 [J].海洋学报,2007,29(4):20-33.
[17]魏皓,赵亮,冯士筰.渤海浮游植物生物量与初级生产力变化的三维模拟 [J].海洋学报,2003,25(2):66-72.
[18]赵亮.渤海浮游植物生态动力学模型研究 [D].青岛:青岛海洋大学,2002.
[19]刘浩,潘伟然.营养盐负荷对浮游植物水华影响的模型研究[J].水科学进展,2008,19(3):345-351.
[20]魏皓,赵亮,冯士筰.渤海碳循环与浮游植物动力学过程研究[J].海洋学报,2003,25(Supp.2):151-156.
[21]Zhang X L,Wu Z M,Li J,et al.Modeling study of seasonal variation of the pelagic-benthic ecosystem of the Bohai Sea[J].Journal of Ocean University of China:English Edition,2006,5(1):21-28.
[22]王强.渤海环流的季节变化及浮游生态动力学模拟 [D].青岛:中国海洋大学,2004.
[23]Zhao L,Wei H.The influence of physical factors on the variation of phytoplankton and nutrients in the Bohai Sea [J].Journal of Oceanography,2005,61(2):335-342.
[24]Tian T,Merico A,Su J,et al.Importance of resuspended sediment dynamics for the phytoplankton spring bloom in a coastal marine ecosystem [J].Journal of Sea Research,2009,62(4):214-228.
[25]Fasham M J R,Duklow H W,McKelvie S M.A nitrogen-based model of plankton dynamics in the oceanic mixed layer[J].Journal of Marine Research,1990,48(3):591-639.
[26]Popova E.Non-universal sensitivity of a robust ecosystem model of the ocean upper mixed layer[J].Ocean Modelling,1995,109:2-5.
[27]蒋红,崔毅,陈碧鹃,等.渤海近20年来营养盐变化趋势研究[J].海洋水产研究,2005,26(6):61-67.
[28]于志刚,米铁柱,谢宝东,等.二十年来渤海生态环境参数的演化和相互关系 [J].海洋环境科学,2000,19(1):15-19.
[29]高会旺,杨华,张英娟,等.渤海初级生产力的若干理化影响因子初步分析 [J].青岛海洋大学学报,2001,31(4):487-494.
[30]Kourafalou V H,Oey L Y,Wang J D,et al.The fate of river discharge on the continental shelf 1.Modeling the river plume and the inner shelf coastal current[J].Journal of Geophysical Research,1996,101:3415-3434.
[31]Taylor A,Watson A,Ainsworth M,et al.A modeling investigation of the role of phytoplankton in the balance of carbon at the surface of the North Atlantic[J].Global Biogeochemical Cycles,1991,5(2):151-171.
[32]邹立,张经.渤海春季营养盐限制的现场实验 [J].海洋与湖沼,2001,32(6):672-678.
[33]Cloern J.The relative importance of light and nutrient limitation of phytoplankton growth:a simple index of coastal ecosystem sensitivity to nutrient enrichment[J].Aquatic Ecology,1999,33(1):3-15.
[34]Colijn F,Cadée G.Is phytoplankton growth in the Wadden Sea light or nitrogenlimited?[J].Journal of Sea Research,2003,49(2):83-93.
[35]Kromkamp J,Peene J,Rijswijk P,et al.Nutrients,light and primary production by phytoplankton and microphytobenthos in the eutrophic,turbid Westerschelde estuary (The Netherlands)[J].Hydrobiologia,1995,311(1):9-19.
猜你喜欢
草业学报(2022年5期)2022-07-08
环境工程技术学报(2022年3期)2022-06-05
当代水产(2021年8期)2021-11-04
环境科学导刊(2021年2期)2021-01-09
当代水产(2019年8期)2019-10-12
当代水产(2019年9期)2019-10-08
农家科技下旬刊(2017年6期)2017-07-05
绿色科技(2016年24期)2017-03-30
天津农业科学(2016年4期)2016-04-20
湖北农业科学(2014年1期)2014-03-22
