
中华鳖Trionyx sinensis (Wiegmann)对泥鳅Misgurnus anguillicaudatus (Cantor)幼鱼的捕食效应*
2014-04-16 03:32袁向阳邹李昶申屠琰朱卫东王志铮
海洋与湖沼 2014年1期
袁向阳 邹李昶, 申屠琰 朱卫东 王志铮
中华鳖(Wiegmann)对泥鳅(Cantor)幼鱼的捕食效应*
袁向阳1邹李昶1, 2申屠琰2朱卫东2王志铮1①
(1. 浙江海洋学院 舟山 316022; 2. 余姚市水产技术推广中心 宁波 315400)
以规格6m × 4m × 1.2m的室内水泥池为实验单元, A[(37.53 ± 7.72)mm]、B[(63.13 ± 12.95)mm]肛长组泥鳅幼鱼为猎物, 采用捕食者-猎物间捕食效应研究方法, 研究了中华鳖对泥鳅幼鱼的捕食效应。结果表明: (1) 中华鳖捕食泥鳅幼鱼的时段主要集中于00:00—4:00, 其对A肛长组泥鳅幼鱼所具的捕食选择性与A、B肛长组间初始数量的配置比例无关; (2) 中华鳖捕食泥鳅幼鱼的功能反应属Holling-Ⅱ型, 其对A、B肛长组泥鳅幼鱼的理论最大捕食量分别为16.13ind/d和9.06ind/d; (3) 中华鳖对A、B肛长组泥鳅幼鱼的种内干扰模型可分别由各自的指数方程表达,用来描述实验鱼初始数量、实验期间实验鱼被捕食数量以及中华鳖个数等参数的统计关系, 并指导确定合理的中华鳖和泥鳅的初始放养数量。
中华鳖; 泥鳅幼鱼; 肛长; 捕食作用
中华鳖(Wiegmann)和泥鳅(Cantor)因具分布范围广、生态适应性强、营养经济价值高等特点而成为我国重要的淡水养殖对象。在水中除用鳃呼吸外, 中华鳖的口咽腔与付膀胱辅助呼吸特性及其攀爬运动习性(沈卉君等, 1983; 杨振才等, 1999), 泥鳅的皮肤与肠辅助呼吸特性及其钻泥习性(金燮理等, 1986), 不仅有益于它们适应稻田养殖环境, 而且也有助于它们参与稻田水、土相间生源要素的搬运与转移, 故在稻田生境科学配置中华鳖和泥鳅的数量密度及两者相互比例关系, 无疑对于持续推进稻田生态系统物质与能量的高效运转和促进稻米安全高效产出具有现实意义。但目前国内外有关捕食效应的研究多见于植保领域敌害昆虫的生物防治(严英俊, 1989; Hoy, 1991; Fan, 1994), 而有关水生生物间捕(滤)食效应的研究却仍较为少见(王伟定等, 2008; 谢志浩等, 2009; 王志铮等, 2011), 至于中华鳖与泥鳅种间相互关系的定量研究则迄今尚未见报道。鉴于此, 笔者借鉴陆上昆虫捕食者-猎物间捕食效应的研究方法, 于2012年7月在余姚市水产技术推广中心实验基地开展了中华鳖对两肛长组泥鳅幼鱼捕食效应的实验研究, 以期通过探析中华鳖对泥鳅的捕食规律, 为进一步优化稻田生境中中华鳖、泥鳅混养模式参数和切实提高稻田养殖产出效益提供理论依据。
1 材料与方法
1.1 材料
1.1.1 泥鳅幼鱼 购自余姚市马渚镇青丰水产养殖场, 运回余姚市水产技术推广中心实验基地并用3%的食盐水浸洗10min后, 立即将其暂养于若干直径120cm、高100cm的白色塑料桶内(实际水位40cm)。暂养期间, 每日早晚各投喂1次, 日投喂量为体质量的3%—5%。实验时选取其中体表光滑、运动活泼、无伤无畸的健壮个体作为实验用鱼。实验鱼按肛长依次分为A [(37.53±7.72)mm]、B [(63.13±12.95)mm]二组。其中, A肛长组实验鱼肛长范围设置与《DB34/T 743-2007无公害食品泥鳅稻田生态养殖技术规程》(安徽省质量技术监督局, 2007)1)规定的放养规格近似。
1.1.2 中华鳖 购自余姚市冷江鳖业有限公司, 运回余姚市水产技术推广中心实验基地后立即移入若干规格为6m×4m×1.2m的水泥池内暂养, 待其排空体内异物后, 选取其中规格相近、肢体完整、反应灵敏、无伤病的健壮个体作为实验对象, 规格为: 体质量(416.67±14.21)g、甲长(146.27±1.94)mm、甲宽(121.70±1.20)mm。
1.1.3 实验条件 以4m×6m×1.2m的室内水泥池为实验单元(实验实际水位70cm), 在室内自然光照条件下进行, 各实验总时间均为24h, 均以当日上午8:00作为实验开始时刻, 以次日上午8:00作为实验结束时刻, 实验期间全程充气。实验用水为经48h自然曝气的自来水, 水温30.5—33.5°C, pH 7.1, 水质符合《NY 5051-2001无公害食品淡水养殖用水水质》(中华人民共和国农业部, 2001)要求。
1.2 试验方法
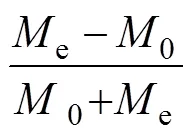
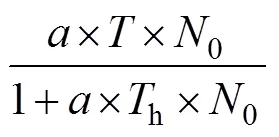
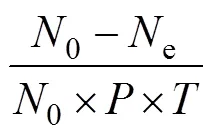
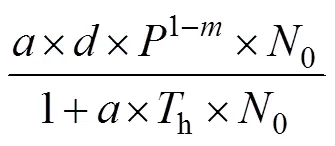
2 结果
2.1 中华鳖对泥鳅幼鱼的捕食特征
将实验对象放入实验池后即下潜、静伏并集群混栖于某一池角。20:00许, 实验对象趋于分散, 实验鳖开始沿池壁爬行, 少数实验鱼开始在池底做缓慢运动, 延至22:00, 实验对象完全分散, 实验鳖活动范围已扩展至整个池底, 绝大多数泥鳅幼鱼开始朝池底中央区域集中, 此前未发现实验鳖捕食泥鳅幼鱼现象。22:00—24:00间, 实验鳖对游经其近前方的泥鳅幼鱼开始表露伸脖捕食行为, 但捕食成功率较低, 捕食量仅占整个实验周期总捕食量的10%左右。24:00起实验对象运动速度均明显加快, 已有近75%泥鳅幼鱼游离池底层, 实验鳖频繁运动于实验水体空间, 并开始间歇性追捕运动于其近前方的实验鱼, 至翌日04:00, 实验鱼被捕食量已达整个实验周期总捕食量的80%左右, 7:00后未见捕食现象发生, 实验对象渐次下潜并静伏于池底。观察发现, 20:00至翌日4:00为实验对象的主要活动期。期间, B肛长组实验鱼的运动量和逃避追捕的窜游速度均明显大于A肛长组, 实验鳖对A肛长组实验鱼的追捕行为较B肛长组更为频繁。
2.2 中华鳖对泥鳅幼鱼的捕食选择性
由表1可见, 在稻田鳅种上限放养密度水平下, 中华鳖对A、B肛长组实验鱼的捕食选择特征表现为: (1) A肛长组实验鱼的被捕食量随其初始数量的下降而显著减少, 呈Ⅰ>Ⅱ>Ⅲ>Ⅳ (<0.05); B肛长组实验鱼的被捕食量与其初始数量占比有关, 呈Ⅴ>Ⅳ>Ⅲ≈II; A、B肛长组实验鱼复配组别中, 仅Ⅳ组A、B间被捕食量无差异(>0.05), 其余均呈A>B (<0.05); (2) 实验鱼总被捕食量与其肛长组组成以及不同肛长组间初始数量的配置比例均有极为密切的关系, 单一肛长组呈Ⅰ>Ⅴ (<0.05), 两肛长组复配组别呈Ⅱ≈Ⅲ>Ⅳ, A、B肛长组初始数量占比较高的组别分别呈Ⅰ>Ⅱ (<0.05)和Ⅳ≈Ⅴ (>0.05), 与A、B肛长组初始数量占比相等的Ⅲ均无差异的组别则呈Ⅱ>Ⅳ (<0.05); (3) 两肛长组复配组别中, A肛长组实验鱼被捕食选择系数均为正值且无差异(>0.05), 而B肛长组被捕食选择系数则均为负值且呈Ⅱ≈Ⅲ>Ⅳ。
综上可知, A、B肛长组实验鱼初始数量比例配置的变化虽可显著影响实验鳖对A肛长组实验鱼的捕食强度(<0.05), 进而分化其对实验鱼的日总捕食量, 但并未能改变实验鳖对A肛长组实验鱼的捕食选择强度(>0.05), 表明实验鳖对小肛长规格实验鱼具极为明显的捕食选择性。
2.3 中华鳖捕食泥鳅幼鱼的功能反应
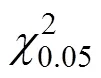
2.4 中华鳖自身数量对泥鳅幼鱼捕食作用的影响
由表3可见, 实验鳖对A、B肛长组实验鱼的捕食作用率值均随实验鳖数量的逐步增加而渐次表露快速下降、缓慢下降和最后趋于稳定的负密度效应, 表明在捕食过程中实验鳖相邻个体间存在较为明显的干扰反应, 且该反应强度随实验鳖自身数量密度的提高而逐渐趋于稳定。
表1 中华鳖对不同肛长组泥鳅幼鱼的捕食选择性
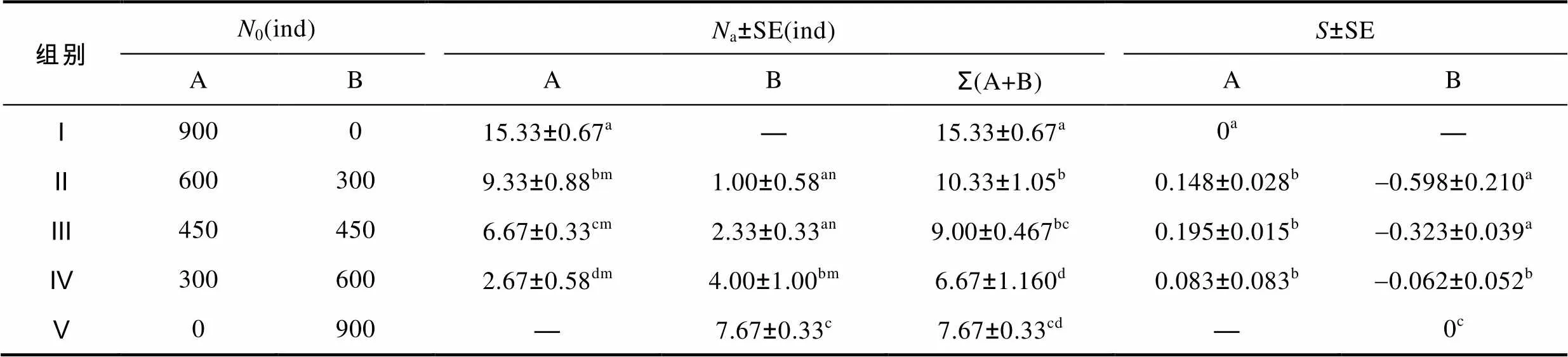
Tab.1 Selective predation of T. sinensis on M. anguillicaudatus with different snout-lengths
A表示肛长范围为(37.53±7.72)mm; B表示肛长范围为(63.13±12.95)mm, 下同; 用上标a、b、c、d标注组间差异(<0.05), 字母相同的表示无差异; 用上标m、n标注组内差异(<0.05), 字母相同的表示无差异
表2 中华鳖捕食不同肛长组泥鳅幼鱼的功能反应模型
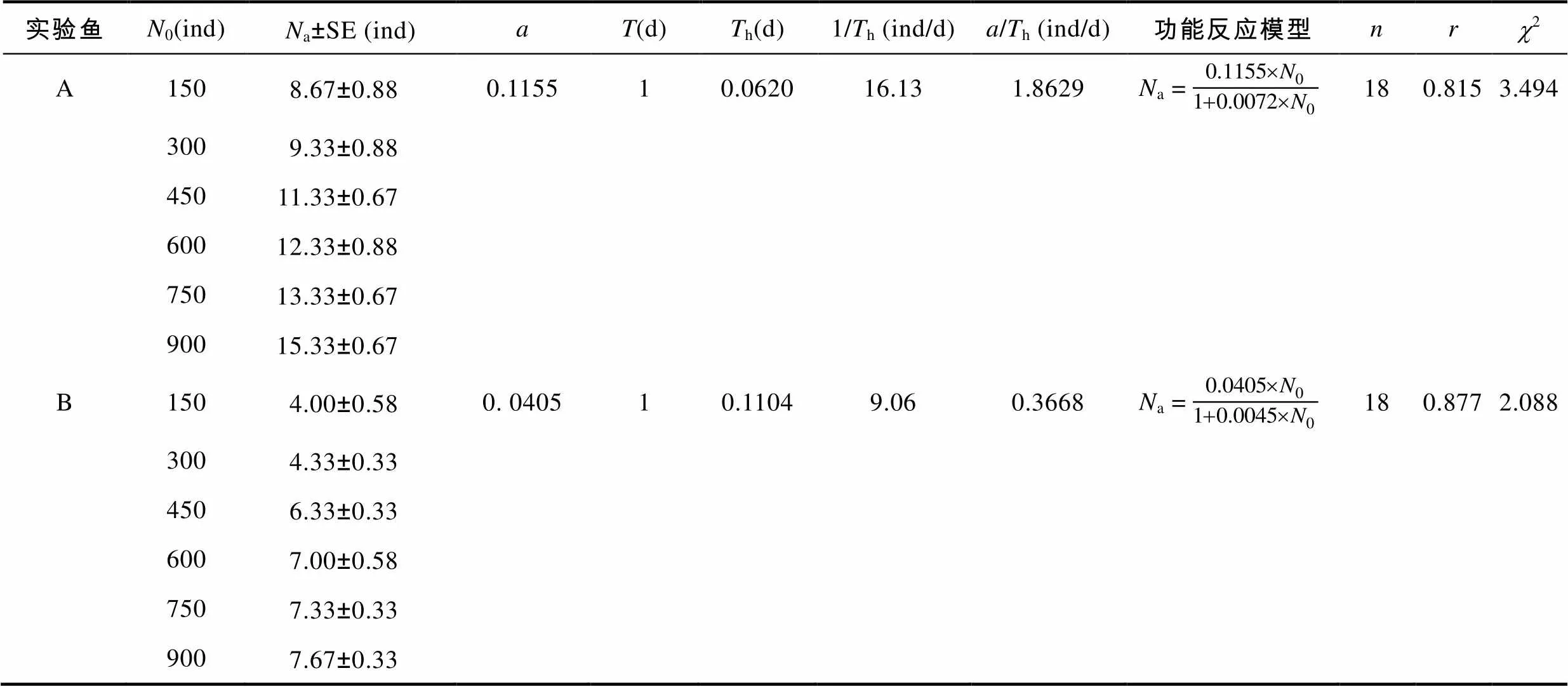
Tab.2 The predatory functional response model of T. sinensis on M. anguillicaudatus with different snout-lengths
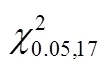
2.5 中华鳖捕食泥鳅的种间相互干扰反应
表3 中华鳖自身数量对不同肛长组泥鳅幼鱼捕食作用的影响
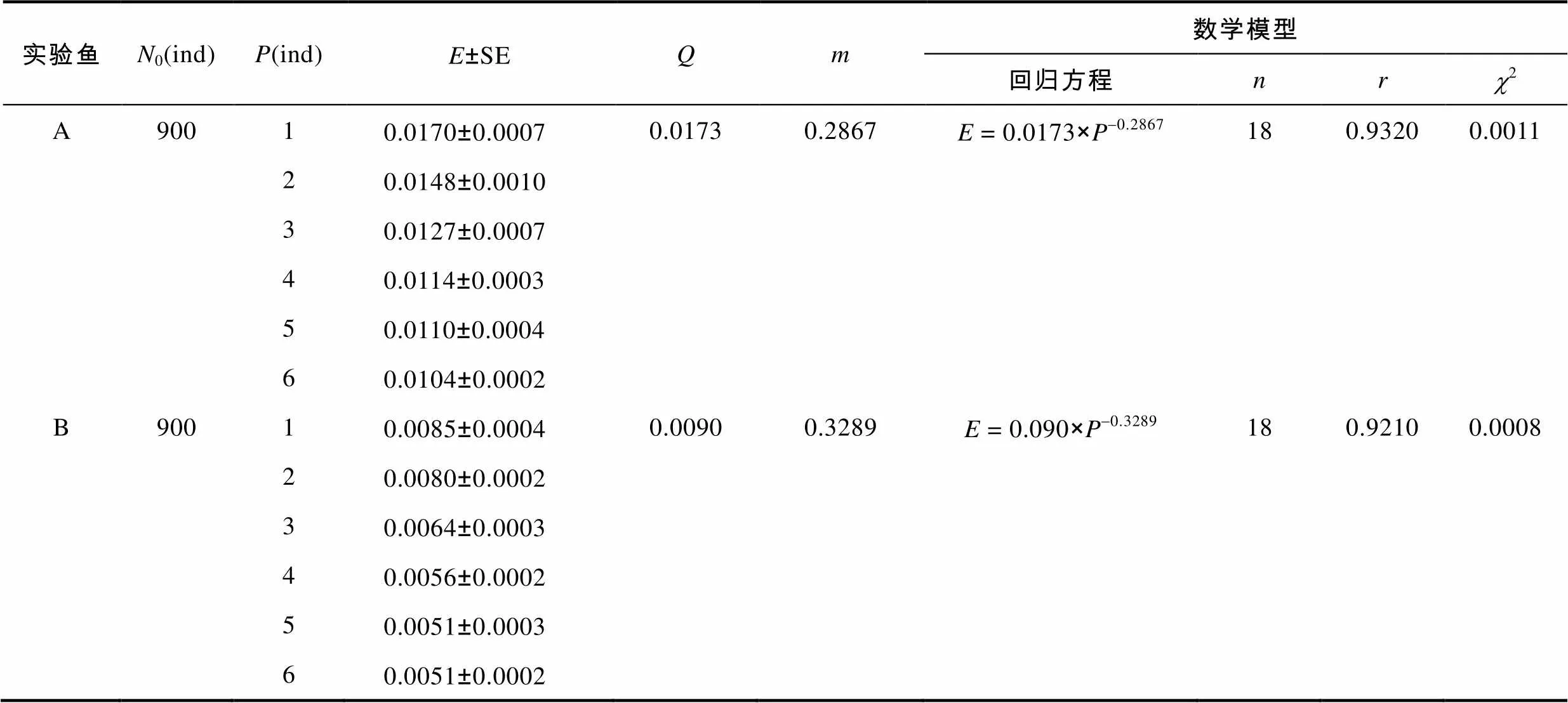
Tab.3 The predatory effect of varying number of T. sinensis on M. anguillicaudatus with different snout-lengths
表4 中华鳖捕食不同肛长组泥鳅幼鱼的相互干扰反应
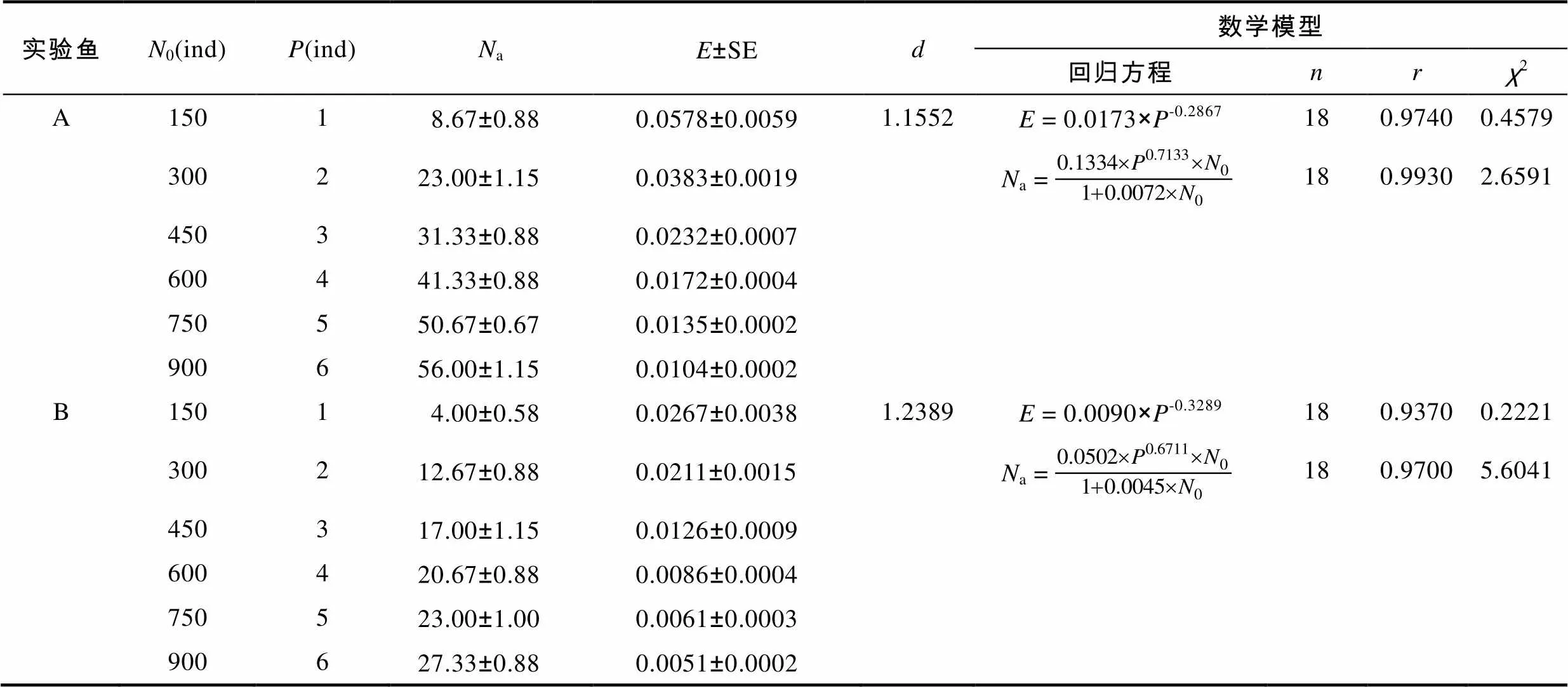
Tab.4 The mutual interferences in the predation of T. sinensis on M. anguillicaudatus with different snout-lengths
3 讨论
3.1 中华鳖对泥鳅幼鱼的捕食选择特征
据报道, 虽中华鳖在水中的视觉差, 运动也较迟钝, 但其位听感觉和触觉均较灵敏, 对外界任何微弱的声响都会迅速作出趋向和逃避反应, 其捕食水生动物主要依靠嗅觉(沈卉君等, 1984)。本研究中, 中华鳖对A肛长组实验鱼的捕食选择特征不受A、B肛长组实验鱼初始数量比例配置的变化而发生改变(表1), 实验鳖对A肛长组实验鱼的瞬时攻击率()、日理论最大捕食量(1/h)以及日捕食效能(/h)均大于B肛长组(表2), 以及中华鳖受种内、种间相互干扰下的搜索常数、干扰常数均依次呈A>B和A3.2 中华鳖与泥鳅幼鱼间捕食效应的生态学意义及其在稻田养殖中的应用价值
由表2可知, 中华鳖捕食A、B肛长组实验鱼的功能反应均属Holling-Ⅱ型, 表明被捕食过程中泥鳅幼鱼群体的空间分布格局具有较为明显的离散性和无序性, 与其遇中华鳖捕食时做间歇性窜游逃逸的观察结果相吻合; 由表3、表4可知, 受种内和种间干扰下, 中华鳖对A、B肛长组实验鱼的捕食均具较为明显的负密度效应, 表明中华鳖与泥鳅幼鱼间数量密度的变化对功能反应的作用是交互的, 符合自然种群中猎物的密度与捕食者的密度对功能反应的作用特征(严英俊, 1989)。与此同时, 中华鳖捕食作用的强时效性(主要捕食时段为0:00—4:00, 约占实验周期总捕食量的70%)、对运动逃逸能力较弱群体的强捕食选择性(表1), 以及鳖鱼混养在水体耗氧间的互利关系(常培新, 2011; 田功太等, 2012), 均有助于实现泥鳅养殖群体的保优汰劣和安全高质产出, 对于切实保障和促进“稻-中华鳖-泥鳅幼鱼”种养复合系统生源要素的正常运转和食物安全高效产出具有重要的生态学意义。上述这些都为本研究模型的应用提供了重要的理论与实践支持。
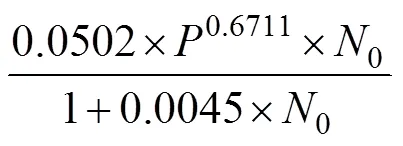
王伟定, 王志铮, 杨 阳等, 2008. 黑暗条件下缢蛏()对牟氏角毛藻()和青岛大扁藻()的滤食效应. 海洋与湖沼, 39(5): 523—528
王志铮, 杨 磊, 朱卫东等, 2011. 日本鳗鲡()对日本沼虾()的捕食效应. 海洋与湖沼, 42(1): 107—113
中华人民共和国农业部, 2001. NY5051-2001无公害食品淡水养殖用水水质. 北京: 中国标准出版社, 1—5
田功太, 巩俊霞, 张金路等, 2012. 中华鳖不同生态养殖模式对池塘水环境及养殖效果的影响. 水生态学杂志, 33(3): 96—100
安徽省质量技术监督局, 2007. DB34/T 743-2007无公害食品泥鳅稻田生态养殖技术规程
沈卉君, 虞 快, 1983. 中华鳖的解剖研究Ⅳ: 消化、呼吸、泌尿生殖系统. 上海师范学院学报, 12(4): 43—50
沈卉君, 虞 快, 1984. 中华鳖的解剖研究V: 感觉器官和神
经系统. 上海师范学院学报, 13(1): 77—86
严英俊, 1989. 草间小黑蛛对褐稻虱的捕食作用及其模拟模型. 福建农学院学报, 18(3): 289—294
周集中, 陈常铭, 1987. 捕食者对猎物选择性的数量测定方法. 生态学报, 7(1): 50—55
杨振才, 牛翠娟, 孙儒泳, 1999. 中华鳖生物学研究进展. 动物学杂志, 34(6): 41—44
金燮理, 李传武, 1986. 泥鳅(Cantor)生物学的初步研究. 湖南师范大学: 自然科学学报, 9(2): 59—66
常培新, 2011. 鱼鳖混养的生物学基础及技术. 现代农业科技, 40(15): 332—333
谢志浩, 王 悠, 唐学玺, 2009. 东海原甲藻()对中华哲水蚤()摄食和消化酶活性的影响. 海洋与湖沼, 40(4): 494—499
Fan Y Q, Petitt F L, 1994. Functional response ofHughes on two-spotted spider mite (Acari: Tetranychidae). Experimental and Applied Acarology, 18(10): 613—521
Hassell M P, Varely G C, 1969. New inductive population model for insect parasites and its bearing on biological. Nature, 223: 1133—1136
Hoill C S, 1959. Some characteristics of simple type of predation and parasitism. Can Entonol, 91: 385—398
Hoy C W, Glenister C S, 1991. Releasingspp. (Acarina: Phytoseiidae) to control(Thysanoptera: Thripidae) on cabbage. BioControl, 36(4): 561—573
PREDATION HABIT OF TRIONYX SINENSIS ON JUVENILE MISGURNUS ANGUILLICAUDATUS
YUAN Xiang-Yang1, ZOU Li-Chang1, 2, SHENTU Yan2,ZHU Wei-Dong2, WANG Zhi-Zheng1
(1. Zhejiang Ocean University, Zhoushan 316022, China; 2. Fishery Technology Extension Center of Yuyao, Ningbo 315400, China)
A method concerning was used to study the predatory efficiencies of(the Chinese softshell turtle) juvenile(the pond loach,), we studied the predation-pray relationship in two groups in snout-vent length at A [(37.53±7.72)mm] and B [(63.13±12.95)mm] of the juveniles cultured in indoor cement tanks (6m×4m×1.2m). The results show that: (1) The hunting time ofconcentrated in 00:00 to 4:00, and its preferential predation onin group A has no relationship to the initial proportion of the juveniles from A and B groups. (2) The type of predatory functional response oftois of Holling-Ⅱmodel, and the theoretical maximal preying quantity were 16.13 (Group A) and 9.06 ind (Group B). (3) The intraspecific interference and interspecific interference models could be expressed in exponential equations for each group case to describe statistically the relationships between turtle and fish of different groups, involving parameters of the number of fish initially fed, number of the fish preyed during the experiment, and number of the turtle farmed. These models or equations can be used to determine best economic practice and provide guidance for the maximum profit to favor turtle farmers.
; juvenile; snout-vent length; predation efficiency
10.11693/hyhz20140100016
* 国家科技部星火计划项目, 2010GA701077号; 宁波市农业领域科技重大攻关择优委托项目, 2012C10032号。袁向阳, 硕士研究生, E-mail: 739226829@qq.com
王志铮, 研究员, E-mail: wzz_1225@163.com
2012-09-12,
2012-12-16
S962, Q958.12
猜你喜欢
作文周刊·小学一年级版(2022年16期)2022-05-07
趣味(作文与阅读)(2021年5期)2021-08-19
作文大王·低年级(2019年2期)2019-01-23
创新作文(小学版)(2018年19期)2018-11-30
小学生导刊(2018年22期)2018-08-21
启蒙(3-7岁)(2018年8期)2018-08-13
琴童(2018年2期)2018-03-06
金色少年(奇趣科普)(2017年8期)2017-09-07
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
