
退化红壤区不同类型人工林土壤活性有机碳及其季节变化
2014-04-11 05:28:38谭桂霞刘苑秋李莲莲昝玉亭李晓东霍炳南何木姣
江西农业大学学报 2014年2期
谭桂霞,刘苑秋 ,李莲莲,刘 武,昝玉亭,李晓东,霍炳南,何木姣
(1.江西农业大学 理学院,江西 南昌 330045;2.江西农业大学 林学院,江西 南昌 330045)
森林土壤碳库是全球土壤碳库中最主要的部分(大约占73%),其变化被许多学者认为是导致大气碳库和全球气候变化的主要原因[1]。但由于土壤有机碳对气候变化、土地管理措施和土地利用方式的反应表现出一定的滞后性且背景值较高,因此,在短时间内很难检测出其发生的微小变化[2]。而土壤活性有机碳虽然占土壤有机碳的比例较小,但它是指示土壤有机碳动态变化的敏感性指标,可检测到微小的、短期的土壤碳动态[3],作为土壤潜在生产力和土壤管理措施变化引起土壤有机质变化的早期指标,可反映各种利用和管理条件下土壤恢复与退化的能力[4-5]。在森林土壤中土壤活性有机碳与树种组成、森林类型密切相关,阔叶林土壤活性有机碳含量高于针叶林[6-7],马尾松低效林采取不同措施改造后土壤有机碳(SOC)、微生物量碳(MBC)、水溶性有机碳(DOC)和易氧化碳(EOC)含量不同程度增加[8]。可见,活性有机碳是反映森林土壤质量和演变的重要指标。
我国南方红壤区由于自然和人为因素已成为中国水土流失范围最广、程度较高的地区[9],退化红壤生态系统的森林植被重建和恢复引起广泛关注。本课题组于1991年在位于江西泰和的退化红壤区开展了不同林分类型植被恢复与重建试验和长期监测研究,结果表明不同类型人工林土壤结构与养分、水分、有机碳、微生物多样性群落与多样性等有不同程度差异[10-12],本文作为长期跟踪研究,在本课题组前期研究的基础上,以21年生的林分为对象,通过不同林分类型土壤可溶性有机碳、易氧化有机碳、土壤微生物生物量碳数量及其季节动态特征,系统探讨林分类型对退化红壤活性有机碳的影响以及在中亚热带气候条件下土壤活性有机碳的季节动态,以进一步揭示退化红壤有机碳积累及土壤恢复机理,也为退化红壤区植被恢复树种选择与配置提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
试验基地位于江西省中部的泰和县境内(26°44'N,115°04'E)。气候属亚热带季风湿润性气候,4—6月份降水较集中,占全年降水量的49%,年降水量1726mm,年均温18.6℃,≥10℃活动积温5921℃。研究区内海拔最高为131.3 m,最低为74.7 m,土壤为第四纪红粘土发育的红壤,为典型的红壤低丘岗地。1991年以绿化荒山、重建森林、恢复退化的生态系统为目标,选择了马尾松(P.massoniana)、湿地松(Pinus elliottii)、枫香(Liquidambar formosana)、木荷(Schima superba)等针阔叶树种重建森林。试验区总面积达133.3 hm2。
本研究选取1991年营造的湿地松纯林、马尾松纯林、湿地松与枫香混交林及枫香纯林为研究对象,并于2012年4月进行了样地基本情况的调查见表1。表1中人工林类型Ⅰ:湿地松纯林;Ⅱ:马尾松纯林;Ⅲ:湿地松×枫香混交林;Ⅳ:枫香纯林。

表1 4种林分类型样地基本概况Tab.1 General site conditions of the four plantations
1.2 研究方法
2012年4月(春季)、7月(夏季)、10月(秋季)、12月(冬季),在4种林分类型中分别选择3个具有代表性的林分设置20 m×20 m样地。在每块样地内按S形选取5个点,去除地表凋落物层,取0~10 cm表层土,5个点混匀作为一个样品,过2mm筛后分成两份。一份放置于冰箱内(3~4℃)供测定土壤可溶性有机碳(DOC)和土壤微生物生物量碳(MBC);另一份自然风干后、进一步处理进行土壤总有机碳(TOC)、易氧化有机碳(ROC)及土壤基本化学性质的测定。测定方法如下:土壤DOC含量的测定采用Bolan等[13]的方法,土壤微生物量碳用氯仿熏蒸-K2SO4浸提法,过滤后滤液中有机碳含量均用TOC仪(ELEMENTAR vario)进行测定。土壤易氧化态碳测定采用重铬酸钾氧化的方法[14]。土壤有机碳(TOC)采用重铬酸钾-浓硫酸外加热法。土壤基本理化性质采用鲁如坤等[15]的分析方法。
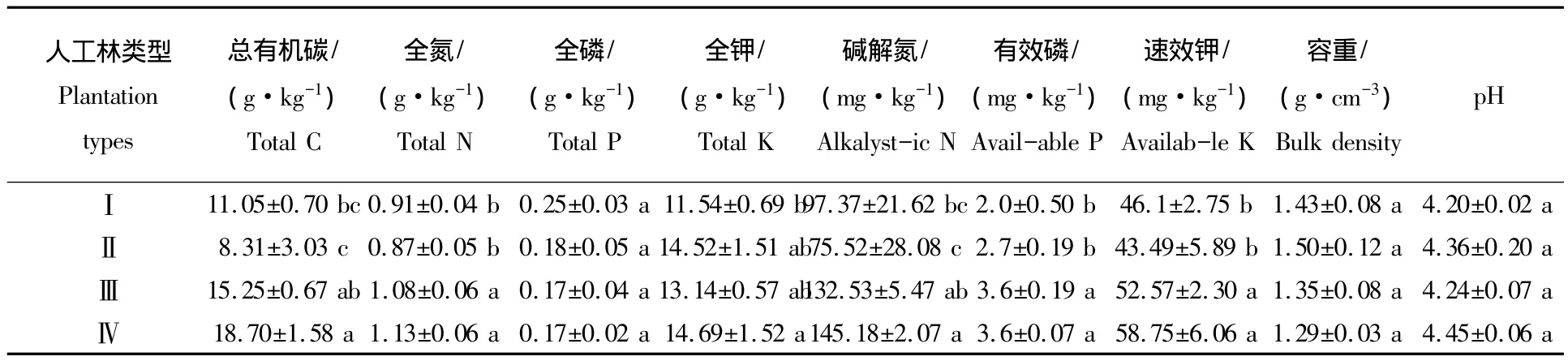
表2 4种林分类型表层土壤基本理化性质Tab.2 Soil physicochemical properties at four plantations
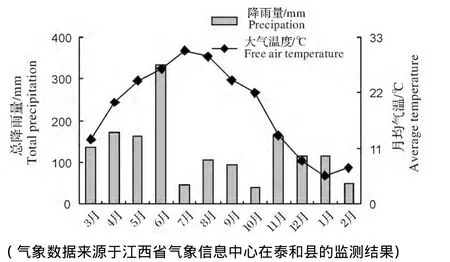
图1 泰和地区2012年3月至2013年2月总降雨量和月平均气温的动态Fig.1 Monthly changes of precipitation and air temperature in Taihe County from Mar.,2012 to Feb.,2013
1.3 数据处理方法
不同类型人工林土壤的基本理化性质、各活性碳年均含量及各种林分不同季节的DOC、ROC、MBC含量均采用单因素方差分析(One-Way ANOVA),再用Tukey多重比较法进行差异显著性分析。显著性水平设定为α=0.05。文中数据均利用Excel 2003和SPSS17.0软件进行数据处理。
2 结果与分析
2.1 不同类型人工林土壤的活性有机碳含量
人工林类型对土壤活性有机碳含量有显著影响(表3)。4种人工林土壤的DOC、ROC、MBC的年平均值分别为52.67 ~105.17 mg/kg、6.68 ~13.33 g/kg、57.70 ~154.21 mg/kg。枫香纯林中的3 种土壤活性有机碳含量都为最高,而湿地松纯林的为最低,且两种林分各活性有机碳含量差异显著。不同林分下土壤DOC的年平均值表现为:枫香纯林>枫香×湿地松混交林>马尾松纯林>湿地松纯林。从差异显著性水平来看,枫香纯林明显高于其它人工林,枫香×湿地松混交林明显高于湿地松纯林,枫香×湿地松混交林和马尾松纯林之间及马尾松纯林与湿地松纯林之间差异并不显著。另外,不同类型人工林土壤的ROC和MBC的年平均值也表现为枫香纯林>枫香×湿地松混交林>马尾松纯林>湿地松纯林,但枫香纯林与枫香×湿地松混交林之间并没有显著差异,马尾松纯林与湿地松纯林之间也没有显著差异。枫香纯林、混交林与两种针叶林之间则差异显著。

表3 不同类型人工林土壤3种活性有机碳的年平均含量Tab.3 Annual average content of soil labile organic carbon in different plantations
2.2 不同类型人工林土壤可溶性有机碳(DOC)季节动态
4种不同林型土壤可溶性有机碳含量均具有明显的季节动态变化,变化规律基本一致,即夏、秋季高于冬、春季(图2)。
马尾松纯林和湿地松纯林DOC含量的最高值均出现在秋季,分别为80.60 mg/kg和64.49 mg/kg;而混交林与枫香林DOC含量的最高值出现在夏季,分别为97.17 mg/kg和105.28 mg/kg。湿地松、马尾松、混交林及枫香林4 种林分 DOC 含量的最低值均出现在春季,分别为44.62,47.12,77.23,88.61 mg/kg。马尾松与湿地松纯林DOC含量表现的季节性变化趋势为秋季>夏季>冬季>春季;混交林与枫香纯林表现出的趋势则是夏季>秋季>冬季>春季。但从差异显著性来看混交林的春、夏、秋、冬四季均呈显著性差异,而枫香纯林则是夏季DOC含量显著高于其它季节,但秋、冬与冬、春之间的DOC含量并无显著差异。湿地松纯林与马尾松纯林的为夏、秋与春、冬季存在显著性差异,而夏季与秋季及春季与冬季之间并无显著性差异。

图2 不同类型人工林土壤DOC的季节的动态Fig.2 Dynamics of dissolved organic carbon of different plantations
2.3 不同林分类型土壤易氧化碳(ROC)季节动态
湿地松纯林、马尾松纯林、混交林、枫香纯林的土壤ROC含量最高值均出现在夏季,分别为8.67、12.22、15.78和15.45 g/kg。湿地松与马尾松纯林的ROC含量最低值出现在冬季,分别为5.48 mg/kg和5.37 g/kg;混交林与枫香纯林的最低值出现在春季,分别为6.40 mg/kg和6.19 g/kg。4种人工林土壤ROC含量的季节变化趋势均夏季>秋季≈春季>冬季(图3)。
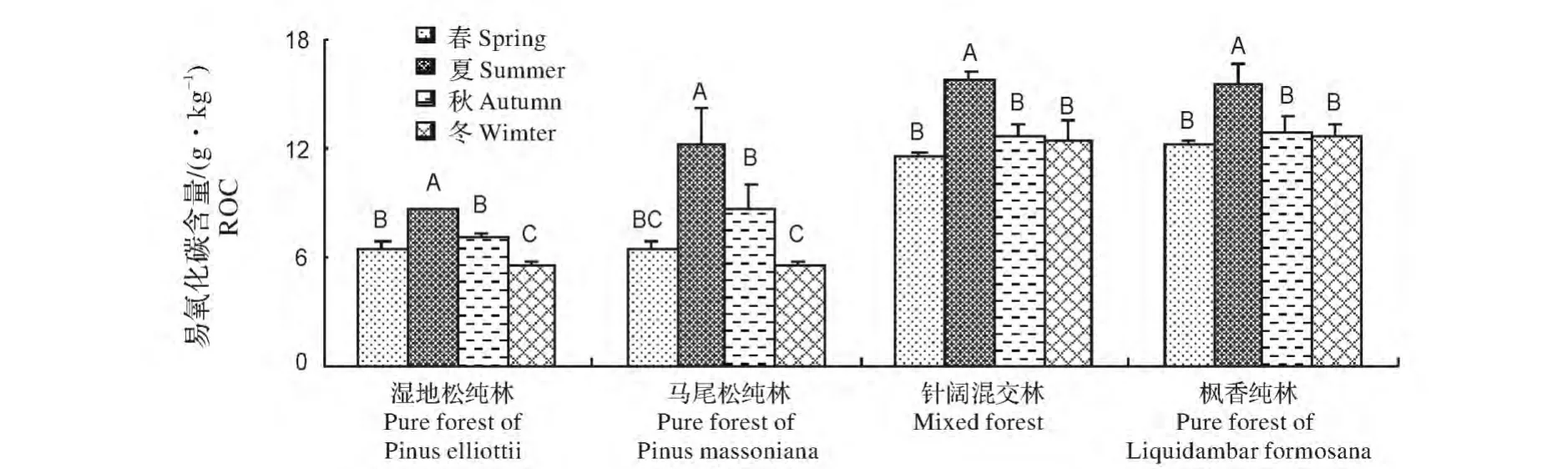
图3 不同类型人工林土壤ROC的季节的动态Fig.3 Dynamics of readily oxidizable organic carbon of different plantations
2.4 不同林分类型土壤微生物量有机碳(MBC)季节动态
湿地松纯林、马尾松纯林、湿地松×枫香混交林、枫香纯林的MBC含量最高值分别为80.58、77.49、195.17、229.71 mg/kg,最低值则分别为 33.87、51.97、81.61、76.03 mg/kg。4 种林分土壤 MBC 含量的最高值均出现在秋季,而最低值均出现在春季。各林分土壤MBC含量随季节变化趋势均为秋季>夏季>冬季>春季,且各季节MBC含量之间均呈显著性差异(图4)。

图4 不同类型人工林土壤MBC的季节的动态Fig.4 Dynamics of soil microbial biomass carbon of different plantations
3 讨论
3.1 退化红壤森林植被恢复土壤活性有机碳含量及其土壤恢复
国内外众多学者对森林土壤DOC和MBC含量进行了大量研究,一些研究结果表明土壤DOC含量在 84 ~376.7 mg/kg[18-19]。森林生态系统土壤 MBC 的变化范围为 500 ~ 2180 mg/kg[18],何友军等[20]对湖南会同杉木林土壤MBC的研究结果为370.9~800.5 mg/kg,朱志建等[21]研究浙江省不同森林植被下土壤MBC含量为282~338 mg/kg,而同处于亚热带的本研究结果为:4种经过21年植被恢复后人工林土壤的DOC和MBC的年平均值仅为52.7~105.2 mg/kg和57.7~154.2 mg/kg。此研究表明,江西泰和退化红壤区的各人工林土壤DOC和MBC含量偏低,而针叶纯林的DOC与MBC含量则更低。
3.2 不同林分类型土壤活性有机碳的差异
本研究结果表明,枫香阔叶纯林活性有机碳含量明显高于湿地松林和马尾松林这两种针叶纯林,湿地松×枫香混交林的土壤各活性有机碳含量也均高于湿地松纯林和马尾松纯林,进一步表明不同树种组成的林分类型是影响土壤活性有机碳的主要方面[19]。这主要是由于针叶林凋落物、根系及其分泌物比常绿阔叶树少得多,土壤中可溶性有机碳产生的少,被微生物快速分解利用[17],而阔叶林和针阔混交林凋落物与根系分泌物诱导而成的微生物区系更加复杂,从而为土壤微生物的活动提供更多能源。刘苑秋等[22]的研究结果表明细根量的多少将影响土壤微生物数量和活性。针阔混交林与针叶纯林相比,细根的生物量增加,与湿地松林和马尾松林等针叶林的凋落物和细根相比,阔叶细根和树凋落物初始氮含量高,碳氮比较小、分解速度较快、较容易被微生物利用,更易于使土壤活性有机碳含量升高。
3.3 土壤各活性有机碳的季节动态
由于土壤的活性有机碳组分对温度、降水等环境条件变化具有高度敏感性,所以土壤活性有机碳含量具有明显的季节变化,降水淋溶和土壤腐殖质的差异是可溶性碳与微生物量碳季节动态差异的重要原因之一[23]。但目前季节动态模式与现有报道结果不尽相同[23-26]。如范跃新等[23]研究中亚热带常绿阔叶林的结果是活性有机碳最低值出现在秋季,最高值随演替进程由冬季逐步转向夏季,刘荣杰等[26]研究浙西北丘陵地区次生林与杉木林的结果是土壤水溶性有机碳含量季节动态均表现为冬季>春季>秋季>夏季,Zhou等[27-28]的结果为DOC含量在秋季达到最高值,在冬季或春季含量最低。
本研究的结果表明4种不同类型人工林土壤DOC、MBC、ROC含量均呈夏、秋高,而春、冬低的趋势。这主要是由于本研究区当年夏、秋两季的降雨量偏少,尤其是取样的7月份和10月份,降雨量仅有39.8mm和45.2mm(图1),为全年降水量最低的月份。所以导致DOC的淋失量也相对偏低,被赋存在土壤中;而且夏、秋季温度高、生物活动比较旺盛,分解土壤中有机质转化为DOC累积在土壤中同时合成自身物质;另外,秋季植物凋落物较多,尤其是阔叶纯林和针阔混交林,DOC经淋溶作用从凋落物进入土壤中,使土壤DOC含量明显增加。而春季降雨量偏大,尤其是春季取样的4月份,降雨量达到了171.5mm(图1),而且整个4月份降雨频繁,大量的DOC可能随雨水淋失,所以DOC含量也为全年最低。冬季时气温较低,微生物活性随之降低,DOC含量也随之下降,同时取样的12月的降雨量也偏大,为113.1mm(图1),有部分DOC随雨水淋失。另外,因为本试验区地处亚热带东部湿润季风气候区,降水量及其季节动态受季风影响年际变化大,因此要准确揭示土壤DOC季节动态规律需要进一步进行连续多年的定位监测研究。
土壤微生物生物量碳的季节变化主要是由土壤温度、土壤湿度、植物生长节律和土壤中可利用碳和养分资源的限制等因素决定[29]。本研究的结果是除了马尾松纯林外,其余3种林分土壤MBC含量均呈秋季>夏季>冬季>春季的变化趋势。这与杨芳等[24]、张水印等[25]的研究结果:MBC含量秋季最高一致。这主要是由于本研究试验点处于亚热带地区,秋季取样是10月份,当月平均气温为22.0℃(图1)处于最适宜微生物活动的温度范围内。另一方面是因为秋季植物根系分泌物和脱落物以及地表凋落物的分解在很大程度上弥补了土壤微生物在能源供应上的不足,促进土壤微生物生物量增长[30]。除此之外,秋季时林木逐渐进入非生长期,植物根系与微生物对营养的竞争减少,有利于微生物生物量增长,导致微生物生物量碳明显增加[23]。各林分土壤MBC含量在春季最低,这是因为春季恰恰是当年的雨季(图1),这与Singh等[31-32]的雨季低、旱季高的研究结果一致。总之,土壤微生物生物量具有复杂的季节波动模式,是各种生态因子综合作用的结果,而土壤温度和土壤湿度的季节性变化是调控土壤微生物生物量季节波动的主要因子[33]。
4 结论
从土壤活性有机碳含量的角度看,江西泰和退化红壤区虽然经过21年森林植被恢复,但土壤质量依然相对较差,土壤恢复还处于初级阶段,土壤质量的恢复是一个较漫长的过程。在不同植被恢复人工林类型中,针阔混交林与阔叶纯林比针叶纯林可以更明显增加土壤活性有机碳的积累,这在改善土壤质量方面比单纯提高土壤总有机碳含量更有意义。由于气温和降水等气候因子的影响,因此,土壤各活性碳组分含量均随季节呈明显动态变化,表现出夏、秋季高,冬、春季低的趋势,淋溶很可能是土壤DOC损失的重要途径;土壤MBC表现为秋季>夏季>冬季>春季。
[1]Sedjor R A.The carbon cycle and global forest ecosystem[J].Water,Air &Soil Pollute,1993,70:295-307.
[2]Haynes R J,Beare M H.Aggregation and organic matter storage in meso-thermal,humid soils[C]//Carter M R,Stewart B A.Advances in soil science,structure and organic matter storage in agriculture soils.Boca Raton:CRC Lewis Publishers,1996.
[3]Xu X,Cheng X L,Zhou Y,et al.Variation of soil labile organic carbon pools along an elevational gradient in the Wuyi Mountains,China[J].Journal of Resources and Ecology,2010,1:368 -374.
[4]Xing S H,Chen C R,Zhou B Q,et al.Soil soluble or ganic nitrogen and active microbial characteristics under adjacent coniferous and broadleaf plantation forests[J].Journal of Soils and Sediments,2010,10:748-757.
[5]王清奎,范冰,徐广标.亚热带地区阔叶林与杉木林土壤活性有机质比较[J].应用生态学报,2009,20(7):1536-1542.
[6]姜培坤.不同林分下土壤活性有机碳库研究[J].林业科学,2005,41(1):l0-13.
[7]耿玉清,余新晓,岳永杰,等.北京山地针叶林与阔叶林土壤活性有机碳库的研究[J].北京林业大学学报,2009,31(5):19-24.
[8]赖家明,李开志,黄丛德,等.不同改造措施对马尾松低效林土壤活性有机碳的影响[J].林业科学研究,2013,26(2):167-173.
[9]赵其国.我国红壤的退化问题[J].土壤,1995,27(6):281-285.
[10]刘苑秋,薛建辉,杜天真,等.森林重建对退化红壤结构特性的影响[J].南京林业大学学报:自然科学版,2003,27(1):16-20.
[11]Gong X,Liu Y Q,Li Q L,et al.Sub-tropic degraded red soil restoration:Is soil organic carbon build-up limited by nutrients supply[J].Forest Ecology and Management,2013,300:77-87.
[12]Jiang Y M,Chen C R,Xu Z H,et al.Effects of single and mixed species forest ecosystems on diversity and function of soil microbial community in subtropical China[J].Soils Sediments,2012,12:228-240.
[13]Bolan N S,Baskaran S,Thiagarajan S.An evaluation of the methods of measurement of dissolved organic carbon in soils,manures,sludges,and stream water[J].Communications in Soil Science and Plant Analysis,1996,27(13):2723-2737.
[14]朱志建,姜培坤,徐秋芳.不同森林植被下土壤微生物量碳和易氧化态碳的比较[J].林业科学研究,2006,19(4):523-526.
[15]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:231-232.
[16]王连峰,潘根兴,石盛莉,等.酸沉降影响下庐山森林生态系统土壤溶液溶解有机碳分布[J].植物营养与肥料学报,2002,8(1):29-34.
[17]王清奎,汪思龙,冯宗炜,等.杉木纯林与常绿阔叶林土壤活性有机碳库的比较[J].北京林业大学学报,2006,28(5):l-6.
[18]Smith J L,Paul E A.The role of soil type and vegetation on microbial biomass and activity.In:Mergusar F,Gantar M eds.Perspectives in microbial Ecology[J].Slovene Society for Microbial Ecology,1988:460-466.
[19]Arunachalarn A,Arunachalarn K.Influence of gap size and soil properties on microbial biomass in a subtropical humid forest of Norih-east India[J].Plant Soil,2000,223:185-193.
[20]何友军,王清奎,汪思龙,等.杉木人工林土壤微生物生物量碳氮特征及其与土壤养分的关系[J].应用生态学报,2006,17(12):292 ~2296.
[21]朱志建,姜培坤,徐秋芳.不同森林植被下土壤微生物量碳和易氧化态碳的比较[J].林业科学研究,2006,19(4):523-526.
[22]刘苑秋,罗良兴,刘亮英,等.退化红壤重建森林初期细根特征及其作用机理研究[J].应用生态学报,2005,16(9):1735-1739.
[23]范跃新,杨玉盛,杨智杰,等.中亚热带常绿阔叶林不同演替阶段土壤活性有机碳含量及季节动态[J].生态学报,2013,33(18):5751-5759.
[24]杨芳,吴家森,钱新标,等.不同施肥雷竹林土壤微生物量碳的动态变化[J].浙江林学院学报,2006,23(1):70-74.
[25]张水印,余明泉,朱玲,等.退化红壤区不同重建模式森林土壤微生物碳氮特征研究[J].江西农业大学学报,2010,32(1):101-107.
[26]刘荣杰,李正才,王斌,等.浙西北丘陵地区次生林与杉木林土壤水溶性有机碳季节动态[J].生态学杂志,2013,32(6):1385-1390.
[27]Zhou G M,Xu J M,Jiang P K.Effect of management practices on seasonal dynamics of organic carbon in soils under bamboo plantations[J].Pedosphere,2006,16:525-531.
[28]黄黎英,曹建华,周莉.不同地质背景下土壤溶解有机碳含量的季节动态及其影响因子[J].生态环境,2007,16(4):1282-1288.
[29]尉海东,董彬.鲁东南杨树人工林土壤微生物生物量碳的时空动态[J].生态环境学报,2013,22(2):233-238.
[30]Wardle D A.A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J].Biological Reviews,1992,67:352-358.
[31]Singh J S,Raghubanshi A S,Singh R S,et al.Microbial biomass acts as a source of plant nutrient in dry tropical forest and savanna[J].Nature,1989,399:499-500.
[32]Barbhuiya A R,Arunachalam A,Pandey H N,et al.Dynamics of soil microbial biomass C,N and P in disturbed and undisturbed stands of a tropical wet-evergreen forest[J].European Journal of Soil Biology,2004,40:113-121.
[33]王国兵,阮宏华,唐燕飞,等.森林土壤微生物生物量动态变化研究进展[J].安徽农业大学学报,2009,36(1):100-104.
猜你喜欢
乡村地理(2022年2期)2022-05-30 10:48:04
农业与技术(2021年6期)2021-12-12 05:33:47
环境与生活(2021年5期)2021-06-24 18:09:52
林业与生态(2020年2期)2020-04-16 12:48:31
森林工程(2018年4期)2018-08-04 03:23:10
方圆(2018年23期)2018-01-07 09:06:18
当代贵州(2016年23期)2016-07-21 08:55:34
广西林业科学(2016年4期)2016-03-16 05:44:59
广西林业科学(2016年4期)2016-03-16 05:44:58
广西林业科学(2016年3期)2016-03-16 05:43:30
