
血雉繁殖初期和育雏期的日活动模式及集群行为研究
2014-03-26 05:29:12段利娟张玉波康东伟王小蓉李俊清
生物学杂志 2014年6期
段利娟, 张玉波, 康东伟, 王小蓉, 李俊清
(1. 北京林业大学 省部共建森林培育与保护教育部重点实验室, 北京 100083; 2. 四川省王朗国家级自然保护区管理局, 四川 绵阳 622550)
日活动模式和集群行为是动物行为学研究的两个重要方面。研究动物的日活动模式有助于了解动物的生物学特性和动物对环境变量的适应性[1]。集群行为是动物常见的社会行为[2],主要研究集群的性别组成、群体大小等,从而反映动物的种群状况及变化趋势[3]。研究动物的日活动模式和集群行为,有助于了解动物的行为特征和生存状况,对于野生动物的保护具有十分重要的意义。
血雉(Ithaginiscruentus)是国家Ⅱ级重点保护野生动物,在中国主要分布于西藏、甘肃、青海、陕西、四川和云南[4]。目前,尽管有一些针对血雉社群组织和行为的研究[5-6],也取得了重要的结果和发现,然而涉及血雉繁殖期尤其是育雏期的日活动模式和集群行为的系统研究较少[7],因此有必要开展相应的研究。
本文利用红外相机监测并记录了王朗自然保护区血雉在繁殖初期和育雏期的行为,分析了血雉的日活动模式和集群行为特征,以期为血雉的保护工作提供科学依据。
1 材料与方法
1.1 研究区域概况
四川王朗国家级自然保护区(103°55′E~104°10′E,32°49′N~33°02′N)建于1965年,总面积32297 hm2。保护区隶属四川省平武县,地处横断山北缘的川西高山峡谷地区,位于青藏高原东缘。地势西北高,东南低,为深山切割型山地,海拔在2300~4980 m之间。气候上属丹巴—松潘半湿润气候区,有暖温带、温带、寒温带、亚寒带、永冻带等带谱类型,区内年平均气温2.9℃,7月平均气温12.7℃,1月平均气温-6.1℃[8]。保护区的植被垂直分布带谱明显,从低海拔到高海拔依次分布着落叶阔叶林、针阔混交林、针叶林、灌丛和高山草甸[9]。
1.2 研究方法
为监测血雉的行为,本研究于2013年4月1日至2013年7月31日在王朗保护区布设了36台被动式数码红外触发相机(5台Ltl AcornTM5210和31台Ltl AcornTM5210A),布设密度为1~4台/4 km2。相机固定于距地面高度30~70 cm的树干或竹子上,并使镜头与地面平行。在相机设置方面,5210型相机设置为拍照模式,5210A型相机设置为拍照后紧接着录像10 s的模式。相机均设置为24 h工作,连续触发间隔为1 min。每1个月检查1次相机,并更换电池和存储卡。将相机连续工作24 h视作一个相机工作日,根据安放相机、更换存储卡和电池的记录表统计每台相机的工作时间段和工作天数。
1.3 数据整理与分析
1.3.1 数据整理
针对设置为拍照兼录像模式的相机,视每单次触发拍摄的照片和视频为一次拍摄,视频作为辅助资料用于记录血雉行为和集群数量。针对所有相机,认为30 min内同一相机拍摄的血雉是一次独立拍摄并统计独立照片数量,以确保拍摄样本的独立性[10],并将30 min内同一相机拍摄到血雉个体最多的一次数据记录为集群数据,辨别并记录照片和视频中血雉的性别和年龄。
1.3.2 拍摄率 采用独立照片数来计算血雉的拍摄率,具体定义如下[11]:
相机工作天数(Total camera days,TCD) = Σcdi
(1)
其中c指工作的相机台数,di是第i台相机正常工作的天数。
拍摄率(Photo trap rate index,PTRI)= (N×100 / ∑cdi)×100%
(2)
其中N是血雉的独立照片数。
1.3.3 繁殖期的划分
红外相机于5月26日第一次拍摄到血雉幼体,因此我们将4月1日—5月25日划分为繁殖初期,5月26日—7月31日划分为育雏期。
1.3.4 日活动模式调查
采用各时间段独立照片数占独立照片总数的比例来描述血雉的日活动模式:
独立照片比例(Proportion of photos, PP)= (Ni/N)×100%
(3)
其中Ni是12个时间段(0:00—2:00, 2:00—4:00, … ,22:00—24:00)内每个时间段拍摄的独立照片总数,i= 1,2,3… 12。
1.3.5 集群类型划分
为便于统计,本研究将集群划分为孤雄、孤雌、雄性群、雌雄混合群和家庭群5种类型。孤雄:由单只成年雄性血雉个体组成。孤雌:由单只成年雌性血雉个体组成。雄性群:由多于一只的成年雄性血雉个体组成。混合群:由成年雄性和成年雌性血雉个体组成。家庭群:由成年雄性和幼体,或成年雌性和幼体,或成年雄性、雌性和幼体组成(幼体群也计入家庭群)。
2 结果
2.1 拍摄率
繁殖初期相机工作1925 d,拍摄血雉照片57张,独立照片数50张,拍摄率为259.74%。
育雏期相机工作2123 d,拍摄血雉照片246张,独立照片数184张,拍摄率为866.70%。
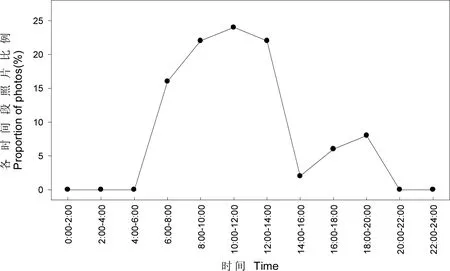
图1 繁殖初期血雉日活动模式
2.2 日活动模式
繁殖初期血雉日活动时间的区间为6:13—19:48,活动高峰出现在6:00—14:00(n=42,P=84.00%),14:00后活动频率很低(图1)。育雏期血雉日活动时间的区间为5:58—19:35,活动高峰出现在6:00—12:00(n=111,P=60.33%),12:00后至18:00也较为活跃(n=63,P=34.24%),而后随日落而停止(图2)。在繁殖初期和育雏期,血雉的日活动都集中在白昼(图1和图2)。

图2 育雏期血雉日活动模式
2.3 集群大小和性比
繁殖初期记录了50群76只血雉,雌雄比例为1∶2.41,平均集群大小为(1.52±0.68)只/群,最大集群数为3。育雏期记录了184群321只血雉,成幼比例为1∶0.34,成体雌雄比例为1∶1.81,平均集群大小为(1.74±1.21)只/群,最大集群数为8。
2.4 繁殖初期和育雏期的集群类型组成
在繁殖初期和育雏期,不同集群类型出现的频率差异均极显著(繁殖初期:x2= 17.680,df=3,P<0.01;育雏期:x2=79.478,df=4,P<0.01)。繁殖初期血雉群体的组成以孤雄(n=23,P=46.00%)和混合群(n=29,P=30.00%)为主,育雏期血雉群体的组成以孤雄(n=75,P=40.76%)和家庭群(n=51,P=27.72%)为主。两个时间段均未记录到雌性群体的出现(图3)。一个家庭群有1~5个幼体,单雄或单雌或一雄一雌与幼体的组合出现频率为67.86%,家庭群中有两个雄性的频率是11.76%。

图3 不同时期不同血雉类型的出现频率(%)
3 讨论
3.1 日活动模式
很多雉类都有昼行性的活动特征[12-14],本文结果表明血雉也存在着明显的昼行性活动规律,即每天在日出后开始活动,至日落时分停止,且血雉在上午的活动频率明显较下午高,表明光照和温度可能是影响血雉活动的重要因素,此外,这一现象也反映了血雉自身的生物学特征。
从血雉自身的日活动规律来看,繁殖初期时血雉的活动高峰为6:00—14:00,14:00之后活动频率突然下降;与繁殖初期的活动规律相似,育雏期时血雉在6:00—14:00时也保持着较高的活动频率,但14:00—18:00的活动频率明显较繁殖初期时高。贾陈喜等[6]对血雉的观察显示,繁殖初期时雌鸟一般只在6:30—14:30之间离巢觅食,到育雏期时,雌鸟不用再回巢孵卵,而是与雄鸟一起育雏,因此,雌性血雉在育雏期时的日活动模式与繁殖初期时有着不同的特征。
3.2 集群大小和性比
本研究发现血雉在繁殖初期的集群大小以1~3只为主,这与鲁庆彬等[15]在4月时观察到的以8~16只为主要集群规模不同。原因可能是:其一、本文研究期间,血雉已经开始分群成对,越冬期形成的大群已分散成小群[16]或形成一雄一雌的配偶关系[6, 16],此外,在雌鸟孵卵期间,雄鸟常单独活动[17];其二、受红外相机的视角范围限制,可能无法完全记录到群体中的全部个体。
从性别看,王朗自然保护区的雄性比例较高,这与李晟等[18]和李桂垣等[16]的结果比较相似。由于繁殖初期雌鸟担负着孵卵的任务,因此,这一期间雄雌比的性比可能较实际数据偏高,而育雏期的性比可能更接近实际情况。
3.3 集群类型组成
血雉为单配制,并在整个繁殖季节都维持稳定的配偶关系[19]。本次研究的集群类型频率分布表示,繁殖初期时混合群的比例较大(32%),其中一雄一雌群体的比例为24%,表明繁殖初期时血雉已经开始分群形成配对;育雏期时家庭群的比例较大(27.72%),且除常见的雌雄亲鸟带领雏鸟活动外,本次研究还记录到家庭群中有两只雄鸟的情况(P=11.76%),这表明血雉的集群性较强,即使在以家庭生活为主的育雏期也常集群活动[17]。动物的集群行为与气候条件、食物丰富度、被捕食压力、资源竞争等环境因素都有密切的关系,也和动物自身息息相关[20]。而血雉在育雏期的集群行为是其自身的生物学特性导致还是存在合作繁殖的现象,这还需要对集群个体间的亲缘关系,非双亲个体在育雏期间的行为和作用等多方面来进一步研究。
参考文献:
[1]Lucherini M, Reppucci J I, Walker R S, et al. Activity pattern segregation of carnivores in the high andes [J]. Journal of Mammalogy, 2009, 90(6): 1404 - 1409.
[2]孙儒泳. 动物生态学原理(第3版) [M]. 北京: 北京师范大学出版社, 2001: 304-314.
[3]李新庆, 刘振生, 王小明, 等. 发情交配期贺兰山岩羊的集群特征 [J]. 兽类学报, 2007, 27(1): 39 - 44.
[4]郑光美. 中国鸟类分类与分布名录(第2版) [M]. 北京: 科学出版社, 2011: 57-58.
[5]贾陈喜, 郑光美, 周小平, 等. 冬季血雉行为的研究 [C]. 第四届海峡两岸鸟类学术研讨会, 2000. 116 - 120.
[6]贾陈喜, 郑光美, 周小平, 等. 卧龙自然保护区血雉的社群组织 [J]. 动物学报, 1999, 45(2): 135 - 142.
[7]贾陈喜, 郑光美, 周小平, 等. 卧龙血雉繁殖期行为特征分析 [J]. 动物学杂志, 2003, 38(1): 37 - 40.
[8]王梦君, 李俊清. 四川省王朗自然保护区地震干扰后大熊猫栖息地的恢复 [J]. 生态学报, 2008, (12): 5848 - 5855.
[9]曾宗永, 岳碧松, 冉江洪, 等. 王朗自然保护区大熊猫对生境的利用 [J]. 四川大学学报:自然科学版, 2002, 39(6): 1140 - 1144.
[10]Kelly M J, Holub E L. Camera trapping of carnivores: trap success among camera types and across species, and habitat selection by species, on salt pond mountain, Giles County, Virginia [J]. Northeastern Naturalist, 2008, 15(2): 249 - 262.
[11]Mohd-Azlan J, Engkamat L. Camera trapping and conservation in lanjak entimau wildlife sanctuary, sarawak, borneo [J]. The Raffles Bulletin of Zoology, 2013, 61(1): 397 - 405.
[12]隆廷伦, 邵开清, 郭 耕, 等. 绿尾虹雉冬季生态的跟踪观测研究 [J]. 四川动物, 1998, 17(3): 104 - 105.
[13]丛培昊, 郑光美. 四川老君山地区红腹角雉的夜栖行为和夜栖地选择 [J]. 生物多样性, 2008, 16(4): 332 - 338.
[14]文陇英, 弓加文, 刘迺发, 等. 四川茂县雉鹑习性的调查 [J]. 动物学杂志, 2008, 43(2): 73 - 76.
[15]鲁庆彬, 王小明, 王正寰. 四川省石渠县高山血雉繁殖初期的集群和生境需求及其相互关系 [J]. 动物学研究, 2006, 27(3): 243 - 248.
[16]李桂垣, 张清茂. 四川王朗自然保护区血雉的野外考察 [J]. 四川动物, 1987, (01): 18 - 21.
[17]贾陈喜, 孙悦华. 血雉 [J]. 动物学杂志, 2008, (05): 143.
[18]Li S, McShea W J, Wang D J, et al. The use of infrared-triggered cameras for surveying phasianids in Sichuan Province, China [J]. Ibis, 2010, 152(2): 299 - 309.
[19]贾陈喜, 郑光美, 周小平, 等. 血雉繁殖期活动区与栖息地特征 [J]. 四川动物, 2004, 23(4): 349 - 352.
[20]刘 强, 杨晓君, 朱建国, 等. 云南省纳帕海自然保护区越冬黑颈鹤的集群特征 [J]. 动物学研究, 2008, 29(5): 553 - 560.
猜你喜欢
垂钓(2023年11期)2024-01-21 16:07:04
小学生优秀作文·趣味阅读(2023年11期)2023-12-30 10:04:58
垂钓(2023年9期)2023-12-10 19:39:30
小学生优秀作文(趣味阅读)(2023年11期)2023-12-02 09:42:18
阅读(快乐英语中年级)(2023年6期)2023-05-24 17:33:15
军事运筹与系统工程(2019年4期)2019-09-11 06:39:58
电子制作(2018年11期)2018-08-04 03:25:40
中国交通信息化(2017年3期)2017-06-08 06:09:28
知识就是力量(2017年2期)2017-01-21 18:29:36
飞碟探索(2016年5期)2016-05-10 23:44:30
