
Restricted nutrient intake does not alter serum-mediated measures of implant response in cell culture
2014-03-25 03:15ChristopherReinhardtTiffanyLeeDanielThomsonLamanMamedovaandBarryBradford
Christopher D Reinhardt,Tiffany L Lee,Daniel U Thomson,Laman K Mamedovaand Barry J Bradford
Restricted nutrient intake does not alter serum-mediated measures of implant response in cell culture
Christopher D Reinhardt1*,Tiffany L Lee2,Daniel U Thomson2,Laman K Mamedova1and Barry J Bradford1
Background:During nutritional stress,reduced intake may reduce the efficacy of anabolic implants.This study was conducted to evaluate basic cellular responses to a growth promotant implant at two intake levels.
Methods:Sixteen crossbred steers(293±19.3 kg)were used to evaluate the impact of anabolic implants in either an adequate or a restricted nutritional state.Steers were trained to individual Calan gates,and then randomly assigned to 1 of 4 treatments in a 2×2 factorial arrangement.Treatments consisted of∶presence or absence of an anabolic growth implant(Revalor-XS,200 mg TBA and 40 mg estradiol;IMPLANT or CONTROL)and a moderate energy,pelleted,starting cattle diet fed at either 2.0×or 1.0×maintenance energy(NEM)requirements(HIGH or LOW).Serum(d 0,14,and 28)was used for application to bovine muscle satellite cells.After treatment with the serum(20%of total media)from the trial cattle,the satellite cells were incubated for 72 h.Protein abundance of myosin heavy chain(MHC),phosphorylated extracellular signal-related kinase(phospho-ERK),and phosphorylated mammalian target of rapamycin(phospho-mTOR)were analyzed to determine the effects of implant,intake,and their interaction(applied via the serum).
Results:Intake had no effect on MHC(P=0.85)but IMPLANT increased(P<0.01)MHC abundance vs.CONTROL. Implant status,intake status,and the interaction had no effect on the abundance of phospho-ERK(P≥0.23). Implanting increased phospho-mTOR(P<0.01)but there was no effect(P≥0.51)of intake or intake×implant.
Conclusions:The nearly complete lack of interaction between implant and nutritional status indicates that the signaling molecules measured herein respond to implants and nutritional status independently.Furthermore,results suggest that the muscle hypertrophic effects of anabolic implants may not be mediated by circulating IGF-1.
Beef cattle,Implant,Muscle,Myosin heavy chain,Nutrient restriction,Satellite cells
Background
Anabolic implants increase average daily gains 15 to 20%,and improve feed efficiency by 6 to 14%in feedlot cattle[1].However,when animals first arrive in a feedlot many are in a state of stress and have not yet become accustomed to the environment,feeding routine,other animals,and other factors,especially if they have traveled long distances or have recently been weaned.These cattle are considered at high risk to develop bovine respiratory disease(BRD).Many of these high-risk cattle have reduced intakes for up to three weeks after arrival at the feedlot[2].
Stress and infection can lead to a hypermetabolic state, characterized by weight loss,increased resting metabolic rate,and muscle protein catabolism[3].In cattle infected with BRD,decreased feed intake,weight loss,and decreased body condition are often observed[3].Although ruminants may have a greater capacity to store digesta because of their larger digestive tracts,this weight loss and decrease in body condition may be comparable to the hypermetabolic state that develops in non-ruminants[3].Munson et al.[4]reported that delaying implant administration in high-risk calves did not alter feedlot performance or the risk of respiratory disease,suggesting that stress brought on by the
transition to the feedlot did not diminish the anabolic signals from implanting early in the finishing phase.
Anabolic implants impact growth rate at least in part by stimulating muscle hypertrophy[5].Androgenic and estrogenic steroids increase in vitro bovine satellite cell proliferation,and these effects are mediated in part by the extracellular signal-regulated kinase(ERK)and phosphatidylinositide 3-kinase/AKT/mammalian target of rapamycin (mTOR)signaling pathways[6].Activation of these pathways,however,is dependent on adequate nutritional status, which is sensed through a complex array of signals[7,8]. Therefore,animals in a depleted nutritional state,accompanied by alterations in hormonal signals,may not have the expected muscle growth response to anabolic implants.
The objectives of this study were to examine the effects of anabolic implants on measures of nutrient balance,metabolic status,and growth factors in animals consuming nutrients either adequate or inadequate to support growth.Moreover,effects of serum-derived signals on in vitro proliferation of bovine muscle satellite cells were also assessed.
Materials and methods
Animals
All procedures conducted in the present study were approved by the Kansas State University Institutional Animal Care and Use Committee(Protocols 2745.0 and 2745.1).
Sixteen crossbred steers(293±19.3 kg)were purchased locally and transported to Kansas State University.They were weighed,tagged,and vaccinated with a 5-way respiratory viral vaccine and a 7-way clostridial vaccine.The steers were trained to individual Calan®gates over a period of 4 wk.
After training was complete,the steers were randomly assigned to 1 of 4 treatments in a 2×2 factorial arrangement.Main effect treatments were as follows∶administration or absence of an anabolic implant(Revalor-XS,200 mg TBA and 40 mg estradiol 17-β[Merck Animal Health,Whitehouse Station,NJ];IMPLANT or CONTROL,respectively)and either 2.0×or 1.0×maintenance energy(NEM)requirements of a moderate energy,pelleted,starting cattle diet(HIGH or LOW, respectively;Table 1).Energy content of the diet was calculated from ingredient composition and tabular estimates of individual ingredient energy values[9].
Steers were weighed on d 0,14,and 28 of the trial.At each of these time points,maintenance energy requirements were calculated for each steer[10],and the diet was fed based these calculations and the treatment assignment for each animal.
Biochemical assays
All assays were conducted in duplicate.Satellite cell isolation was conducted as described by Johnson et al.[5].
Using sterile techniques,500 g of the semimembranosus muscle was dissected from 2 steers which were not part of the present study(18 mo of age).After removal of connective tissue,the muscle was passed through a sterile meat grinder.The ground muscle was incubated with 0.1%pronase in Earl’s Balanced Salt Solution(EBSS)for 1 h at 37°C with frequent mixing.After incubation the mixture was centrifuged at 1,500×g for 5 min,the pellet was suspended in phosphate buffered saline(PBS;Ca-and Mg-free PBS;Life Technologies,Grand Island,NY),and the suspension was centrifuged at 500×g for 10 min.The supernatant was centrifuged at 1,500×g for 10 min to pellet the mononucleated cells.The PBS wash and differential centrifugation were repeated 2 more times.Cell viability was over 90%.The resulting mononucleated cell preparation was suspended,pooled(density 3×104cells/mL)in cold DMEM containing 10%fetal bovine serum(FBS)and 10%dimethylsulfoxide and frozen in liquid nitrogen.Cells were counted using a hemocytometer and 0.4%Trypan Blue solution.
Blood samples(serum and plasma)were collected from each animal at d 0,14,and 28 of the trial and used in determining levels of total IGF-1,and additional serum samples were collected from each animal for application to satellite cells[11-13]which had been previously isolated.Total IGF-1 concentration was determined in plasma samples using a commercial ELISA kit(Octeia
IGF-1,catalog#AC27F1,Immunodiagnostic Systems Inc.,Scottsdale,AZ).The kit includes reagents for pretreatment of samples to release IGF-1 from binding proteins[14].Frozen satellite cells were re-suspended in DMEM supplemented with 5%FBS and 1%penicillinstreptomycin mixture(containing 10,000 IU/mL penicillin and 10,000 μg/mL streptomycin).Cells were passaged once,then at 70%confluence,cells were incubated in DMEM for 72 h with serum(20%of total media)taken from individual animals on d 0,14,or 28 and with an additional control containing 20%FBS and no serum from trial cattle.Halting incubation at 72 h prevents overconfluency of the cells,which can lead to excessive differentiation;since these are skeletal muscle cells and would contract or twitch after differentiation,excessive differentiation would lead the cell layers to come off the cell culture plate.All media contained 1%penicillin-streptomycin mixture,but FBS was not added to wells containing treatment serum.
Protein abundance of myosin heavy chain(MHC;d 0, 14,and 28),phosphorylated ERK(phospho-ERK;d 0 and 28),and phosphorylated mTOR(phospho-mTOR; d 0 and 28)was analyzed in differentiated satellite cells to determine the effects of implant,intake,and their interaction(applied via the serum).MHC was used as a marker of myotube formation,and phospho-ERK and phospho-mTOR are regulators of cell proliferation.
Western blot analysis
Cultured cells were lysed at 4°C with RIPA lysis buffer (50 mmol/L Tris-HCl,pH 7.5,150 mmol/L NaCl,1%NP-40,0.5%sodium deoxycholate,and 0.%SDS;Santa Cruz Biotechnology,Santa Cruz,CA)containing X1 broadspectrum protease inhibitor cocktail(Protease inhibitor cocktail I;Calbiochem,Gibbstown,NJ)and centrifuged at 15,000×g for 10 min at 4°C.Protein concentration was measured using a Coomassie protein assay kit(Pierce, Rockford,IL)according to Bradford[15].The proteins in the supernatant were denatured with SDS buffer containing dithiothreitol for 5 min at 95°C.Forty micrograms of protein were loaded in each lane,and proteins were separated using SDS-PAGE on a 4-12%TRIS-glycine gel, then transferred to a nitrocellulose membrane(iBlot;Invitrogen,Carlsbad,CA).Membranes were blocked in Tris buffer(pH 7.4)with 5%dry milk powder for 2 h at room temperature and then incubated with the following antibodies∶a rabbit anti-MYH(H-300)polyclonal antibody raised against amino acids 1641-1940 of myosin heavy chain 3(sc-20641;Santa Cruz Biotechnology,Santa Cruz, CA);rabbit anti-phospho-mTOR(Ser 2481)polyclonal antibody(Cell Signaling Technology,Beverly,MA),and rabbit anti-phospho-ERK(Tyr 204;sc-7976,Santa Cruz Biotechnology)overnight at 4°C.All primary antibodies were diluted 1∶1,000.After washing,membranes were incubated for 1 h at room temperature with a secondary antibody(anti-rabbit IgG,sc-2301;Santa Cruz Biotechnology)diluted 10,000-fold in Tris buffer(pH 7.4). Immunodetection was performed by chemiluminescence(West-Dura;Thermo Scientific,Waltham,MA), and band images were visualized using a photodocumentation system(ChemiDoc-It Imaging System;UVP Inc.,Upland,CA).Densitometry analysis was carried out using Image J(http∶//rsbweb.nih.gov/ij/).
Statistical analysis
Animal was the experimental unit for all measures.Data were analyzed using the MIXED procedure of SAS®(v.9.3; SAS Institute,Cary,NC);IGF-1 and MHC were analyzed using repeated measures and d 0 values were included in the model as a covariate;phospho-ERK and phosphomTOR were analyzed as a completely randomized design but included d 0 values as a covariate.The independent variables included the main and interactive effects of implant status and intake level.Effects were considered significant with a protected F-test with α<0.05;effects were considered a trend if P<0.10.
Results and discussion
Level of feeding achieved different growth rates(0.76 vs. 0.02 kg/d for 2.0×vs.1.0×maintenance intake,respectively;P<0.01);implant status did not affect 28-d growth rates(0.36 vs.0.41 kg/d for implanted vs.control,respectively;P=0.38).There was no interaction between level of intake and implant status for growth rate(P=0.38).
There was no intake×implant for serum IGF-1 levels (P=0.56).High-intake steers had greater IGF-1 levels (P=0.02)vs.low-intake steers(Figure 1).Elsasser et al. [16]reported that greater energy intake increases circulating IGF-1 levels in growing steers,and Hayden et al. [17]reported reduced IGF-1 values in steers when intake was restricted by 23%.
The IGF-1 concentrations in implanted steers were not different from those in control cattle in this study.Bryant et al.[18,19]reported no effects of implants on IGF-1 in feedlot heifers until d 42 after implant administration; however,Johnson et al.[18,19]and Reinhardt[20]reported that circulating IGF-1increased in implanted steers by d 21 and d 28,respectively,compared to non-implanted steers. Anderson et al.[21]reported that exogenous estradiol supplementation in post-menopausal women increased circulating growth hormone 2.1-fold but actually decreased circulating IGF-1 by 27%.

Figure 1Effects of implant or intake level on plasma ureanitrogen.Plasma urea nitrogen concentration of steers(ChartA) either implanted(solid line)or not implanted(dashed line)with a long-acting TBA∶E2implant;and(ChartB)fed a common diet at either 2.0(solid line;HIGH)or 1.0(dashed line;LOW)×maintenance NE requirements.(Effect of intake P=0.56;implant P=0.62;intake× implant P=0.99;day×intake P<0.01;day×implant P=0.33;day× intake×implant P=0.12.Pooled SE for treatment means was 0.261.a,bmeans without a common superscript differ(P<0.05).
When serum was applied to the cultured satellite cells from the semimembranosus muscle of non-study steers, level of intake had no effect(P=0.85;Figure 2)on MHC abundance but implant increased MHC abundance(P<0.01),with no interaction observed between intake level and implant status(P=0.23).In this study, MHC was used as a marker of myotube formation, reflecting satellite cell proliferation and differentation. Baxa et al.[22]found that implanting increased MHC type IIa mRNA in vivo.In a smaller study,Baxa[23] found that implanting with a combined estradiol-17β (E2)/trenbolone acetate(TBA)led to numerical increases in MHC type I and IIa mRNA.Chung et al.[24]found no effect of E2/TBA implants on MHC type I,IIa or IIx mRNA,but did find that MHC IIa increased over time which was also seen in the present study(effect of day P<0.01).Implanting with E2/TBA increases muscle fiber diameter[25,26],due to an initial increase in DNA transcription followed by an increase in nuclei within the muscle fiber which support hypertrophy,but ultimately due to an increase in total protein produced by the cell[27],of which MHC is a key constituent.
Extracellular signal-regulated kinase plays a role in regulating insulin-and potentially IGF-I-mediated glucose uptake by skeletal muscle cells[28],and certain fatty acids reduce muscle cell proliferation via ERK[29]. Implant status did not affect the phospho-ERK abundance(P=0.46;Figure 3).Intake had no effect(P= 0.65)on phospho-ERK,and there was no interaction between the two main effects(P=0.41).Because ERK is phosphorylated(activated)by IGF-I signaling[6,30],the lack of phospho-ERK response to implant in the present study is understandable,given the lack of effect on circulating IGF-1.However,intake level also failed to influence phospho-ERK despite the fact that HIGH increased serum IGF-1 compared to LOW.
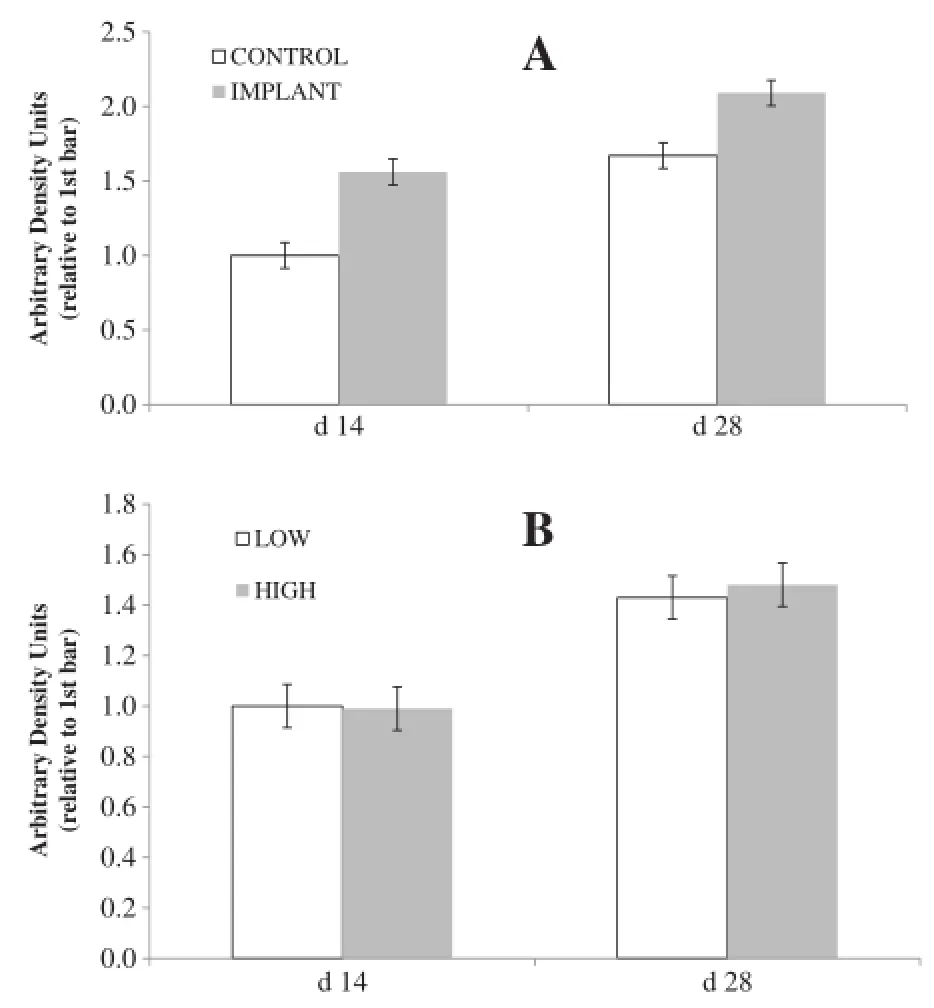
Figure 2Effects of implant and intake level on myosin heavy chain abundance in cultured myocytes.Myocytes from a nonstudy steer were treated with serum(20%of total media)from study steers drawn on d 0,14,and 28,and myosin heavy chain abundance was measured.In chartA,steers were either implanted(gray bars) or not implanted(open bars)with a long-acting TBA∶E2implant;in chartB,steers were fed a common diet at either 2.0(gray bars; HIGH)or 1.0(white bars;LOW)×maintenance NE requirements.(Effect of day P<0.01;intake P=0.85;implant P<0.01;intake×implant P=0.23;day×intake P=0.88;day×implant P<0.01;day×intake× implant P=0.44;Error bars represent pooled SEM for treatment means=0.103).

Figure 3Effect of implant and intake level on phosphorylated extracellular signal-related kinases(phosphor-ERK).Myocytes from a non-study steer were treated with serum(20%of total media)from study steers drawn on d 0 and 28,and phospho-ERK abundance was measured.Steers were(ChartA)either implanted (gray bars)or not implanted(white bars)with a long-acting TBA∶E2implant and(ChartB)fed a common diet at either 2.0(gray bars; HIGH)or 1.0×(white bars;LOW)maintenance NE requirements. (Effect of intake P=0.65;implant P=0.47;intake×implant P=0.41). Pooled SE for treatment means was 0.065.
The protein kinase mTOR plays an essential part in transcription,translation,muscle cell size,and protein stability[31,32].To help regulate these processes,mTOR integrates information about nutrient status,energy status, and many growth factors and may be a central component in the regulation of cellular processes that determine cell growth and proliferation,and even ultimate overall animal size[33].Phosphorylation of mTOR can occur at both Ser 2448[34]and Ser 2481[35].We investigated Ser 2481 phosphorylation of mTOR.This site is responsive to nutrient supply and Ser 2481 phosphorylation status is directly correlated with activity of mTOR complex 1,which is responsible for promoting protein synthesis through activation of S6 kinase 1 and eukaryotic translation initiation factor 4E-binding protein 1[35].There was no interaction between intake and implant status(P=0.88;Figure 4)for phospho-mTOR,and intake had no effect(P=0.51);however,serum from implanted steers(P<0.01)stimulated greater phospho-mTOR levels vs.serum from CONTROL steers.Phosphorylated mTOR normally increases when nutrition is adequate for growth and decreases when the nutrient environment is not favorable for growth[31,36]. Although we did not see this response,it is important to recognize that our experimental design only partially subjected satellite cells to the milleu that muscle tissue of LOW animals would have seen.Our protocol provided cells with essentially unlimited nutrient supply via the DMEM that comprised 80%of the media,thereby negating effects of intake level on nutrient supply to the satellite cells.The lack of intake effect on phospho-mTOR,therefore,suggests only that endocrine signals(or compounds not supplied in DMEM)that differed by intake failed to alter satellite cell mTOR signaling.Yang et al.[32]reported that mTOR is highly sensitive to upregulation by IGF-1 and that the stress caused by low nutrient availability can inhibit mTOR signaling.In the present study,although we saw no effect of implant on IGF-1,we did measure a strong response of implant on phospho-mTOR. We saw a significant effect of increased intake level on IGF-1 but no corresponding effect of high nutrient intake stimulating phospho-mTOR.

Figure 4Effects of implant and intake level on phosphorylated mammalian target of rapamycin(phospho-mTOR)abundance in cultured myocytes.Myocytes from a non-study steer were treated with serum(20%of total media)from study steers drawn on d 0 and 28.In chartA,steers were either implanted(gray bars; IMPLANT)or not implanted(open bars;CONTROL)with a longacting TBA∶E2implant;in chartB,steers were fed a common diet at either 2.0(gray bars;HIGH)or 1.0(white bars;LOW)×maintenance NE requirements.(Effect of intake P=0.51;implant P<0.01;intake× implant P=0.88.Error bars represent pooled SEM=0.134).
Johnson et al.[18,19]reported increased mitogenic activity of serum from implanted vs.control steers when applied directly to satellite cell cultures,but that study also reported increased levels of circulating IGF-I in the implanted steers.Results from the present study suggest that elevated IGF-I is not the sole serum-mediated factor which increases signalling for increased mitogenic activity in vitro,given that serum from nutrient abundant and nutrient restricted calves,which differed greatly in IGF-I concentration,did not alter MHC or phosphomTOR.The phospho-mTOR and MHC response to implant,in the absence of a circulating IGF-I response, could be explained by a number of additional,nonsomatomedin-mediated pathways associated with growth regulation.Binding proteins specific for growth hormone and some,although not all,for IGF-I increase in plasma in response to elevated nutrient availability independent of treatment with exogenous growth hormone[37],and the same study indicated that elevated nutrient status actually reduced growth hormone in plasma.Concentration of IGF-I mRNA is elevated in longissimus muscle
cells from animals previously administered exogenous E2[38,39],and muscle mRNA concentrations for the adipogenic regulators PPARγ and stearoyl CoA desaturase are reduced after administration of exogenous TBA[24].In addition,both E2and TBA,which were used in combination in the present study,stimulate protein synthesis and reduce protein degradation in a dose-dependent manner when directly applied to fused satellite cell cultures[40,41].This action is at least partially modulated through stimulation of muscle IGF-I mRNA by E2and TBA[42].
Treatment of proliferating bovine satellite cells cultures with either E2or TBA resulted in increases in IGF-1 mRNA as well as increased3H-thymidine incorporation rate[42].Direct treatment of fused bovine satellite cell cultures with E2increases protein synthesis and decreases protein degradation rates,and blocking of estradiol binding suppresses estradiol-induced modifications in protein synthesis and degradation[40].Testosterone also increases satellite cell activation in rats[43].These documented effects indicate a direct mechanism for both estradiol and TBA on satellite cell proliferation and protein synthesis.
Implanting with a combination of E2and TBA increases circulating estradiol and TBA,but only exogenous E2increases circulating growth hormone[44-46]. Circulating estradiol is correlated to circulating growth hormone in humans[47],and exogenous E2supplementation increased growth hormone secretion in postmenopausal women[48]and ovariectamized ewes[49]. Sotiropoulos et al.[50]reported that growth hormone stimulates myoblast fusion with myotubes.Ge et al.[51] reported that growth hormone did not stimulate myoblast fusion or proliferation,but growth hormone did increase protein synthesis.
Hepatocyte growth factor increases satellite cell activation[52-54]and fibroblast growth factor enhances proliferation of satellite cells in culture[55,56].Although hepatocyte growth factor[57-59]and fibroblast growth factor[60,61]are elevated in vitro in response to treatment with E2,liver and muscle mRNA for hepatocyte growth factor are not affected in vivo after implanting cattle with E2and TBA[62].
The satellite cell results,where serum from implanted cattle activated mTOR signaling,make it tempting to speculate that mTOR is a more critical mediator of muscle hypertrophy in response to implants than ERK. Certainly,mTOR has been shown to be a key regulator of protein synthesis[8]and is activated by testosterone treatment[63].The primary upstream regulators of mTOR are AMP-activated protein kinase(AMPK),AKT, and ERK.Blockade of either the AKT and ERK signalling pathways ablated the mitogenic response to trenbalone acetate in bovine satellite cells,suggesting that at least basal activation of both of these pathways is required [6].Conversely,activation of AMPK,which is triggered by nutrient deficiency,impairs mTOR phosphorylation, and this pathway has been implicated in loss of muscle mass in nutrient-restricted cows[36].Given that implant status did not enhance phospho-ERK abundance in this study,we propose that basal ERK signaling was likely necessary to permit activation of mTOR,but that activation of the AKT pathway was probably more directly involved in stimulating mTOR signaling.This is also consistent with the unique roles of ERK and mTOR signaling pathways.Although ERK does contribute to mTOR activation,it is probably more important as a regulator of cell proliferation[64].Proliferation of satellite cells is important for maintainance of future hypertrophic potential in vivo,but myotube formation is driven by cell differentation and activation of protein synthesis,which is more closely related to the function of mTOR.
Conclusions
No interactions were found between nutrient restriction and implant status in muscle satellite cell culture treated with serum from calves either implanted or not implanted and fed at either 1×or 2×maintenance.This suggests that the effects of exogenous anabolic agents and nutrient status on growth regulation,at least insofar as it is regulated by the molecules measured in this study,are independent of one another.Serum from implanted cattle promoted increased abundance of the active phospho-mTOR regulatory protein as well as myotube formation,measured by MHC abundance,in bovine satellite cells.These finding suggest the presence of circulating factors in implanted cattle that influence cell signals and promote hypertrophy of muscle tissue.
Competing interests
The authors declare that they have no competing interests.
Authors’contributions
CR is the corresponding author;TL gathered all samples,participated in laboratory procedures,conducted statistical analysis,and wrote the primary draft of the manuscript;DT created the concept and design of the study;LM coordinated all laboratory analyses;BB was responsible for determining the appropriate analytical methods to be conducted.All authors have read and approved the final manuscript.
Author details
1Department of Animal Sciences and Industry,232 Weber Hall,Kansas State University,Manhattan,KS 66506,USA.2Department of Clinical Sciences, A-111 Mosier Hall,Kansas State University,Manhattan,KS 66506,USA.
Received∶31 May 2013 Accepted∶22 October 2013
Published∶19 November 2013
1. Duckett SK,Andrae JG∶Implant strategies in an integrated beef production system.J Anim Sci 2001,79(E.Suppl)∶E110-E117.
2. Hutcheson DP,Cole NA∶Management of transit-stress syndrome in cattle∶nutritional and environmental effects.J Anim Sci 1986,62∶555-560.
3. Cole NA,Delaney DD,Cummins JM,Hutcheson DP∶Nitrogen metabolism of calves inoculated with bovine adenovirus-3 or with infectious bovine rhinotracheitis virus.Am J Vet Res 1986,47∶1160-1164.
4. Munson RD,Thomson DU,Reinhardt CD∶Effects of delayed steroid implanting on health,performance,and carcass quality in high health risk,auction market-sourced feedlot steers.J Anim Sci 2012,90∶4037-4041.
5. Johnson BJ,Halstead JN,White ME,Hathaway MR,DiConstanzo A,Dayton WR∶Activation state of muscle satellite cells isolated from steers implanted with a combined trenbolone acetate and estradiol implant. J Anim Sci 1998,76∶2779-2786.
6. Kamanga-Sollo E,White ME,Hathaway MR,Chung KY,Johnson WR,Dayton WR∶Roles of IGF-I and the estrogen,androgen and IGF-I receptors in estradiol-17β-and trenbolone acetate-stimulated proliferation of cultured bovine satellite cells.Domest Anim Endocrinol 2008,35∶88-97.
7. Lewis MI,Bodine SC,Kamangar N,Xu X,Da X,Fournier M∶Effect of severe short-term malnutrition on diaphragm muscle signal transduction pathways influencing protein turnover.J Appl Physiol 2006,100∶1799-1806.
8. Howell J,Manning M∶mTOR couples cellular nutrient sensing to organismal metabolic homeostasis.Trends Endocrinol Metab 2011,22∶94-102.
9. NRC∶Nutrient requirements of beef cattle.7threv.ed.Washington,D.C∶Natl. Acad.Press;1996.
10.NRC∶Nutrient requirements of beef cattle.6threv.ed.Washington,D.C∶Natl. Acad.Press;1984.
11.Loomis RJ,Marshall LA,Johnston PV∶Sera fatty acid effects on cultured rat splenocytes.J Nutr 1983,113∶1292-1298.
12.Jewell DE,Drewry MM,Martin RJ,Hausman GJ∶Effect of sera from control and overfed rats on preadipocyte growth in culture.J Nutr 1988, 118∶803-808.
13.Reecy JM,Williams JE,Kerley MS,MacDonald RS,Thornton WH Jr,Wallace LM∶Abomasal casein infusion enhances the mitogenic activity of serum from protein-restricted steers.J Nutr 1994,124∶67-77.
14.Krebs A,Wallaschofski H,Spilcke Liss E,Kohlmann T,Brabant G,Völzke H, Nauck M∶Five commercially available insulin-like growth factor I(IGF-I) assays in comparison to the former Nichols Advantage IGF-I in a growth hormone treated population.Clin Chem Lab Med 2008,46∶1776-1783.
15.Bradford MM∶A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Anal Biochem 1976,72∶248-254.
16.Elsasser TH,Rumsey TS,Hammond AC∶Influence of diet on basal and growth hormone-stimulated plasma concentrations of IGF-1 in beef cattle.J Anim Sci 1989,67∶128-141.
17.Hayden JM,Williams JE,Collier RJ∶Plasma growth hormone,insulin-like growth factor,insulin,and thyroid hormone association with body protein and fat accretion in steers undergoing compensatory gain after dietary energy restriction.J Anim Sci 1993,71∶3327-3338.
18.Bryant TC,Engle TE,Galyean ML,Wagner JJ,Tatum JD,Anthony RV,Laudert SB∶Effects of ractopamine and trenbolone acetate implants with or without estradiol on growth performance,carcass characteristics, adipogenic enzyme activity,and blood metabolites in feedlot steers and heifers.J Anim Sci 2010,88∶4102-4119.
19.Johnson BJ,Hathaway MR,Anderson PT,Meiske JC,Dayton WR∶Stimulation of circulating insulin-like growth factor I(IGF-I)and insulin-like growth factor binding proteins(IGFBP)due to administration of a combined trenbolone acetate and estradiol implant in feedlot cattle.J Anim Sci 1996,74∶372-379.
20.Reinhardt CD∶Metabolic indices for growth:Endocrine profile of steers on different nutritional and growth regulation regimes,Master’s thesis.College Station,TX∶Texas A&M University;1991.
21.Anderson SM,Shah N,Evans WS,Patrie JT,Bowers CY,Veldhuis JD∶Short-term estradiol supplementation augments growth hormone(GH) secretory responsiveness to dose-varying GH-releasing peptide infusions in healthy postmenopausal women.J Clin Endocrinol Metab 2001, 86∶551-560.
22.Baxa TJ,Hutcheson JP,Miller MF,Brooks JC,Nichols WT,Streeter MN,Yates DA,Johnson BJ∶Additive effects of a steroidal implant and zilpaterol hydrochloride on feedlot performance,carcass characteristics,and skeletal muscle messenger ribonucleic acid abundance in finishing steers.J Anim Sci 2010,88∶330-337.
23.Baxa TJ∶Effect of zilpaterol hydrochloride and steroid implantation on yearling steer feedlot performance,carcass characteristics,and skeletal muscle gene expression,Master’s Thesis.Manhattan,KS∶Kansas State University;2004.
24.Chung KY,Baxa TJ,Parr SL,Luqué LD,Johnson BJ∶Administration of estradiol,trenbolone acetate,and trenbolone acetate/estradiol implants alters adipogenic and myogenic gene expression in bovine skeletal muscle.J Anim Sci 2012,90∶1421-1427.
25.Gonzalez JM,Carter JN,Johnson DD,Ouellette SE,Johnson SE∶Effect of ractopamine-hydrochloride and trenbolone acetate on longissimus muscle fiber area,diameter,and satellite cell numbers in cull beef cows. J Anim Sci 2007,85∶1893-1901.
26.Kellermeier JD,Tittor AW,Brooks JC,Galyean ML,Yates DA,Hutcheson JP, Nichols WT,Streeter MN,Johnson BJ,Miller MF∶Effects of zilpaterol hydrochloride with or without an estrogen-trenbolone acetate terminal implant on carcass traits,retail cutout,tenderness,and muscle fiber diameter in finishing steers.J Anim Sci 2009,87∶3702-3711.
27.Johnson BJ,Reinhardt CD∶Growth promotants for beef production∶Anabolic steroids∶Performance responses and Mode of action.In Current Veterinary Therapy:Food Animal Practice 5thVolume.Edited by Anderson DE, Rings DM.St.Louis,MO∶Saunders/Elsevier,Inc;2009∶643-651.
28.Hwang SL,Jeong YT,Li X,Kim YD,Yue L,Chang YC,Lee IK,Chang HW∶Inhibitory cross-talk between the AMPK and ERK pathways mediates endoplasmic reticulum stress-induced insulin resistance in skeletal muscle.Br J Pharmacol 2013.doi∶10.1111/bph.12124.[Epub ahead of print].
29.Peng Y,Zheng Y,Zhang Y,Zhao J,Chang F,Lu T,Zhang R,Li Q,Hu X,Li N∶Different effects of omega-3 fatty acids on the cell cycle in C2C12 myoblast proliferation.Mol Cell Biochem 2012,367∶165-173.
30.Fuentes EN,Bjornsson BT,Valdes JA,Einarsdottir IE,Lorca B,Alvarez M, Molina A∶IGF-I/PI3K/AkT and IGF-I/MAPK/ERK pathways in vivo in skeletal muscle are regulated by nutrition and contribute to somatic growth in the fine flounder.Am J Physiol 2011,300∶R1532-R1542.
31.Corradetti MN,Guan KL∶Upstream of the mammalian target of rapamycin∶do all roads pass through mTOR?Oncogene 2006, 25∶6347-6360.
32.Yang X,Yang C,Farberman A,Rideout TC,de Lange CFM,France J,Fan MZ∶The mammalian target of rapamycin-signaling pathway in regulating metabolism and growth.J Anim Sci 2008,86∶E36-E50.
33.Sorbassov DD,Ali SM,Sabatini DM∶Growing roles for the mTOR pathway. Curr Opin Cell Biol 2005,17∶596-603.
34.Nave BT,Ouwens M,Withers DJ,Alessi DR,Shepherd PR∶Mammalian target of rapamycin is a direct target for protein kinase B∶identification of a convergence point for opposing effects of insulin and amino-acid deficiency on protein translation.Biochem J 1999,344(Pt 2)∶427-431.
35.Soliman GA,Acosta-Jaquez HA,Dunlop EA,Ekim B,Maj NE,Tee AR,Fingar DC∶mTOR Ser-2481 autophosphorylation monitors mTORC-specific catalytic activity and clarifies rapamycin mechanism of action.J Biol Chem 2010,285∶7866-7879.
36.Du M,Zhu MJ,Means WJ,Hess BW,Ford SP∶Nutrient restriction differentially modulates the mammalian target of rapamycin signaling and the ubiquitin-proteasome system in skeletal muscle of cows and their fetuses.J Anim Sci 2005,83∶117-123.
37.Vestergaard M,Purup S,Frystyk J,Løvendahl P,Sørensen MT,Riis PM,Flint DJ,Sejrsen K∶Effects of growth hormone and feeding level on endocrine measurements,hormone receptors,muscle growth and performance of prepubertal heifers.J Anim Sci 2003,81∶2189-2198.
38.Pampusch MS,Johnson BJ,White ME,Hathaway MR,Dunn JD,Waylan AT, Dayton WR∶Time course of changes in growth factor mRNA levels in muscle of steroid-implanted and non-implanted steers.J Anim Sci 2003, 81∶2733-2740.
39.Pampusch MS,White ME,Hathaway MR,Baxa TJ,Chung KY,Parr SL, Johnson BJ,Weber WJ,Dayton WR∶Effects of implants of trenbolone acetate,estradiol,or both,on muscle insulin-like growth factor-I, insulin-like growth factor-I receptor,estrogen receptor-a,and androgen receptor messenger ribonucleic acid levels in feedlot steers.J Anim Sci 2008,86∶3418-3423.
40.Kamanga-Sollo E,White ME,Hathaway MR,Weber WJ,Dayton WR∶Effect of Estradiol-17 beta on protein synthesis and degradation rates in fused bovine satellite cell cultures.Domest Anim Endocrinol 2010,39∶54-62.
41.Kamanga-Sollo E,White ME,Hathaway MR,Weber WJ,Dayton WR∶Effect of trenbolone acetate on protein synthesis and degradation rates in fused bovine satellite cell cultures.Domest Anim Endocrinol 2011,40∶60-66.
42.Kamanga-Sollo E,Pampusch MS,Xi G,White ME,Hathaway MR,Dayton WR∶IGF-I mRNA levels in bovine satellite cell cultures∶effects of fusion and anabolic steroid treatment.J Cell Physiol 2004,201∶181-189.
43.Joubert Y,Tobin C∶Testosterone Treatment Results in Quiescent Satellite Cells Being Activated and Recruited into Cell Cycle in Rat Levator Ani Muscle.Devel Biol 1995,169(1)∶286-294.
44.Breier BH,Gluckman PD,Bass JJ∶Influence of nutritional status and oeshadiol-17B on plasma growth hormone,insulin-like growth factors-I and-II and the response to exogenous growth hormone in young steers. J Endocrinol 1988,118∶243.
45.Hayden JM,Bergen WG,Merkel RA∶Skeletal muscle protein metabolism and serum growth hormone,insulin,and cortisol concentrations in growing steers implanted with estradiol-17 beta,trenbolone acetate,or estradiol-17 beta plus trenbolone acetate.J Anim Sci 1992,70∶2109-2019.
46.Hongerholt DD,Crooker BA,Wheaton JE,Carlson KM,Jorgenson DM∶Effects of a growth hormone-releasing factor analogue and an estradioltrenbolone acetate implant on somatotropin,insulin-like growth factor I, and metabolite profiles in growing Hereford steers.J Anim Sci 1992, 70∶1439-1448.
47.Milewicz T,Krzysiek J,Sztefko K,Radowicki S,Krzyczkowska-Sendrakowska M∶17 beta-estradiol regulation of human growth hormone(hGH),insulinlike growth factor-I(IGF-I)and insulin-like growth factor binding protein-3(IGFBP-3)axis in hypoestrogenic,hypergonadotropic women. Endokrynol Pol 2005,56∶876-882.
48.Dawson-Hughes B,Stern D,Goldman J,Reichlin S∶Regulation of growth hormone and somatomedin-C secretion in postmenopausal women∶effect of physiological estrogen replacement.J Clin Endocrinol Metab. 1986,63∶424-432.
49.Scanlan N,Skinner DC∶Estradiol modulation of growth hormone secretion in the ewe∶No growth hormone-releasing hormone neurons and few somatotropes express estradiol receptor α.Biol Reprod 2002, 66∶1267-1273.
50.Sotiropoulos A,Ohanna M,Kedzia C,Menon RK,Kopchick JJ,Kelly PA, Pende M∶Growth hormone promotes skeletal muscle cell fusion independent of insulin-like growth factor 1 up-regulation.Proc Natl Acad Sci U S A 2006,103∶7315-7320.
51.Ge X,Yu J,Jiang H∶Growth hormone stimulates protein synthesis in bovine skeletal muscle cells without altering insulin-like growth factor-I mRNA expression.J Anim Sci 2012,90∶1126-1133.
52.Allen RE,Sheehan SM,Taylor RG,Kendall TL,Rice GM∶Hepatocyte growth factor activates quiescent skeletal muscle satellite cells in vitro.J Cell Physiol 1995,165∶307-312.
53.Tatsumi R,Anderson JE,Nevoret CJ,Halevy O,Allen RE∶HGF/SF is present in normal adult skeletal muscle and is capable of activating satellite cells.Devel Biol 1998,194∶114-128.
54.Sheehan SM,Tatsumi R,Temm-Grove CJ,Allen RE∶HGF is an autocrine growth factor for skeletal muscle satellite cells in vitro.Muscle Nerve 2000, 23∶239-245.
55.Gospodarowicz D,Moran J,Braun D,Birdwell C∶Clonal growth of bovine vascular endothelial cells∶fibroblast growth factor as a survival agent. Proc Natl Acad Sci U S A 1976,73∶4120-4124.
56.Rusnati M,Dell’Era P,Urbinati C,Tanghetti E,Massardi ML,Nagamine Y, Monti E,Presta M∶A distinct basic fibroblast growth factor(FGF-2)/FGF receptor interaction distinguishes urokinase-type plasminogen activator induction from mitogenicity in endothelial cells.Mol Biol Cell 1996, 7∶369-381.
57.Taylor KM,Chen C,Gray CA,Bazer FW,Spencer TE∶Expression of messenger ribonucleic acids for fibroblast growth factors 7 and 10, hepatocyte growth factor,and insulin-like growth factors and their receptors in the neonatal ovine uterus.Biol Reprod 2001,64∶1236-1246.
58.Coleman KD,Wright JA,Ghosh M,Wira CR,Fahey JV∶Estradiol modulation of hepatocyte growth factor by stromal fibroblasts in the female reproductive tract.Fertil Steril 2009,92∶1107-1109.
59.Coleman KD,Ghosh M,Crist SG,Wright JA,Rossoll RM,Wira CR,Fahey JV∶Modulation of hepatocyte growth factor secretion in human female reproductive tract stromal fibroblasts by poly(I∶C)and estradiol. Am J Reprod Immunol 2012,67∶44-53.
60.Frey RS,Johnson BJ,Hathaway MR,White ME,Dayton WR∶Growth factor responsiveness of primary satellite cell cultures from steers implanted with trenbolone acetate and estradiol-17b.Basic Appl Myol 1995,5∶71-79.
61.Wing LY,Chuang PC,Wu MH,Chen HM,Tsai SJ∶Expression and mitogenic effect of fibroblast growth factor-9 in human endometriotic implant is regulated by aberrant production of estrogen.J Clin Endocrinol Metab 2003,88∶5547-5554.
62.White ME,Johnson BJ,Hathaway MR,Dayton WR∶Growth factor messenger RNA levels in muscle and liver of steroid-implanted and nonimplanted steers.J Anim Sci 2003,81∶965-972.
63.Wu Y,Bauman WA,Blitzer RD,Cardozo C∶Testosterone-induced hypertrophy of L6 myoblasts is dependent upon Erk and mTOR. Biochem Biophys Res Commun 2010,400∶679-683.
64.Jones NC,Fedorov YV,Rosenthal RS,Olwin BB∶ERK1/2 is required for myoblast proliferation but is dispensable for muscle gene expression and cell fusion.J Cell Physiol 2001,186∶104-115.
doi∶10.1186/2049-1891-4-45
Cite this article as:Reinhardt et al.∶Restricted nutrient intake does not alter serum-mediated measures of implant response in cell culture. Journal of Animal Science and Biotechnology 2013 4∶45.
Submit your next manuscript to BioMed Central and take full advantage of:
· Convenient online submission
· Thorough peer review
· No space constraints or color fi gure charges
· Immediate publication on acceptance
· Inclusion in PubMed, CAS, Scopus and Google Scholar
· Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
*Correspondence∶cdr3@ksu.edu
1Department of Animal Sciences and Industry,232 Weber Hall,Kansas State University,Manhattan,KS 66506,USA
Full list of author information is available at the end of the article
©2013 Reinhardt et al.;licensee BioMed Central Ltd.This is an open access article distributed under the terms of the Creative Commons Attribution License(http∶//creativecommons.org/licenses/by/2.0),which permits unrestricted use,distribution,and reproduction in any medium,provided the original work is properly cited.
 Journal of Animal Science and Biotechnology2014年1期
Journal of Animal Science and Biotechnology2014年1期
- Journal of Animal Science and Biotechnology的其它文章
- GMOs in animal agriculture∶time to consider both costs and benefits in regulatory evaluations
- Dual potential of microalgae as a sustainable biofuel feedstock and animal feed
- Identification and characterization of genes that control fat deposition in chickens
- Haplotype phasing after joint estimation of recombination and linkage disequilibrium in breeding populations
- Comparative analysis of proteomic profiles between endometrial caruncular and intercaruncular areas in ewes during the peri-implantation period
- Changes in feed intake,nutrient digestion,plasma metabolites,and oxidative stress parameters in dairy cows with subacute ruminal acidosis and its regulation with pelleted beet pulp