
Identification and characterization of genes that control fat deposition in chickens
2014-03-25 03:15HirwaClaireAndreWallacePaulXuShenXinzhengJiaRongZhangLiangSunandXiquanZhang
Hirwa Claire D’Andre,Wallace Paul,Xu Shen,Xinzheng Jia,Rong Zhang,Liang Sunand Xiquan Zhang
Identification and characterization of genes that control fat deposition in chickens
Hirwa Claire D’Andre1,3*,Wallace Paul2,Xu Shen3,Xinzheng Jia3,Rong Zhang3,Liang Sun3and Xiquan Zhang3
Background:Fat deposits in chickens contribute significantly to meat quality attributes such as juiciness,flavor, taste and other organoleptic properties.The quantity of fat deposited increases faster and earlier in the fastgrowing chickens than in slow-growing chickens.In this study,Affymetrix Genechip®Chicken Genome Arrays 32773 transcripts were used to compare gene expression profiles in liver and hypothalamus tissues of fast-growing and slow-growing chicken at 8 wk of age.Real-time RT-PCR was used to validate the differential expression of genes selected from the microarray analysis.The mRNA expression of the genes was further examined in fat tissues.The association of single nucleotide polymorphisms of four lipid-related genes with fat traits was examined in a F2resource population.
Chicken,Fat deposition,Genes
Background
Fat deposition is a crucial aspect in modern chicken breeding schemes because it is associated with selection for increased body weight in broilers[1-7].The growth of broiler chicken is accompanied by an increased percentage of body fat with a concomitant increase in the mass of abdominal and visceral fat[8].The quantity of fat deposited increases faster and earlier in fastgrowing chickens than in slow-growing chickens[9-12]. Excessive adiposity is a problem in modern broiler industry[13];and needs to be controlled to reduce negative effects on productivity,acceptability,and health of consumers.In meat-type chickens,excessive adipose tissue decreases both feed efficiency during rearing and the yield of lean meat after processing.However,fat is the major contributor to meat flavor;and the presence of
intramuscular fat confers high eating quality of meat. Therefore,regulating fat deposition plays an important role in broiler chicken production.
In birds,lipogenesis,takes place primarily in the liver whereas adipocyte serves as the storage site for triglycerides[14].Hepatic lipogenesis contributes 80 to 85%of the fatty acids stored in adipose tissue[15]because lipogenic activity in chickens is much greater in the liver than in adipose tissue[16-18].
In the past decade,genetic mechanisms underlying chicken fat deposition were widely studied but few studies were conducted to determine the gene expression involved in pathways as well as mechanisms that lead to adiposity in chickens[19].In the present study,fastgrowing White Recessive Rock chickens(WRR)and slow-growing Xinghua chickens(XH)were used to characterize specific genes for fat deposition in chickens.Global gene expression patterns within the liver and hypothalamus tissue of WRR and XH chickens were determined using Partek GS 6.4 Affymetrix Genechip®Chicken Genome Arrays and the differentially expressed genes were identified.Some of the differentially expressed genes were validated by determining their mRNA expression in liver,hypothalamus and fat tissues.The association of single nucleotide polymorphisms of the genes with chicken fat traits was also investigated.
Materials and methods
Chicken populations
Eight WRR(4♂+4♀,Institute of Animal Science,Guangdong AcademyofAgriculturalSciences,Guangzhou, China),and 8 XH chickens(4♂+4♀,Fengkai Zhicheng Poultry Breeding Company,Guangdong,China),were used for differential expression observation with microarray hybridization.All the birds were fed a nutritionally balanced corn-soybean diet[20].The birds had free access to water.They were slaughtered at 8 wk of age,and the liver and hypothalamus were excised,snapped frozen in liquid nitrogen and stored at-80°C until required for further analyses.
Six sets of WRR(3♂+3♀),and another six of XH (3♂+3♀),were used to study mRNA expression of the SULT1B1,PNPLA3,GPAM,ELOVL6,LPL,FASN, ACSBG2,FDPS,and FRZB genes in abdominal fat,subcutaneous fat,breast muscle,and pituitary tissues in the liver and hypothalamus tissues.
For association analysis,an F2resource population was constructed by crossing WRR with XH chickens [21].The fat traits such as abdominal fat weight, subcutaneous fat thickness,fat band width,abdominal fat percentage were recorded in all F2full-sib individuals.
Ethics statement
The study was approved by the Animal Care Committee of South China Agricultural University(Guangzhou, People's Republic of China).Animals involved in this study were humanely sacrificed as necessary to ameliorate their suffering.
Microarray hybridization and data preprocessing
Total RNA was isolated from frozen tissues(50 mg) using TRIzol reagent(Invitrogen,CA,USA)according to the manufacturer’s instructions.Total RNA concentration was determined by spectrophotometry.The RNA labellingandmicroarrayhybridization were carried outaccordingto theAffymetrix Expression Analysis Technical Manual(Biochip Corporation,Shanghai, China).The arrays were scanned using the Affymetrix Scanner 3000.
The GeneChip Chicken Genome Array used in the present study was created by Affymetrix Inc.(Santa Clara,USA)at the end of 2006,with comprehensive coverage of over 38,000 probe sets representing 32,773 transcripts corresponding to over 28,000 chicken genes (Chicken Genome Sequencing Consortium 2.1).Sequence information for this array was selected from the following public data sources∶GenBank,UniGene and Ensembl.
Data normalization was used to eliminate dye-related artifacts.Consecutive filtering procedures were performed to normalize the data,and to remove noise derived from absent genes,background,and nonspecific hybridizations.Comparisons of expression levels were performed for each gene,and genes with the most significant differential expression(P<0.05)were retained. Raw data sets were normalized to total fluorescence, which represents the total amount of RNA hybridized to a microarray,using the Partek GS 6.4(Affymetrix Genechip®Chicken Genome Arrays,USA).QVALUE was used to obtain false-discovery rates(FDR).
The data obtained were subjected to Partek GS 6.4 for comparison using Affymetrix Expression Console Software,for expression algorithm robust multi-array (RMA)analysis.Multivariate ANOVA was used to determine significant differences among the replicates.Differentially expressed genes between WRR and XH chickens were identified by cutoff of fold-change(fold change)≥2 and P<0.05.Molecular functions of differentially expressed genes were classified according to molecule annotation system(MAS)3.0(http∶//bioinfo.capitalbio. com/mas3/).Database from the Kyoto Encyclopedia of Genes and Genomes(KEGG)were used for pathway analysis on differentially expressed genes using AgriGO (GO Analysis Toolkit and Database for Agricultural Community)http∶//bioinfo.cau.edu.cn/agriGO/)and Database for Annotation,Visualization and Integrated Discovery
(DAVID)Bioinformatics Resources(http∶//david.abcc. ncifcrf.gov/).
Validation of the differential expression with real-time RT-PCR
The primers were designed based on the published cDNA sequences of SULT1B1,the LPL,ELOVL6, ACSBG2,SCD5,FADS1,PNPLA3,GAPDH,BEAN, SLC22A2,DDT,PLA2G12A,and 18S genes(http∶// www.ncbi.nlm.nih.gov)usingGENETOOLsoftware (BioTools,Alberta,Canada).The RNA was reversetranscribed using the RevertAid Fist Strand cDNA Synthesis(Toyobo,Japan).After reverse transcription, the cDNA of the selected genes were amplified by real-time reverse transcription PCR.The relative level of each mRNA normalized to the 18 s gene was calculated using the following equation∶fold change= 2Ct target(WRR)-Ct target(XH)/2Ct 18S(WRR)-Ct 18S(XH).
The linear amount of target molecules relative to the calibrator was calculated by 2-ΔΔCT.Therefore,all gene transcription results are reported as the n-fold difference relative to the calibrator.Specificity of the amplification product was verified by electrophoresis on a 0.8%agarose-gel.The results were expressed as mean±SE.
Fat tissue expression of the differential expression genes with real-time RT-PCR studies
The same primers as those used in validation were used for determining fat tissue expression.The real-time RTPCR reactions were performed using the iCycler Real-Time PCR detection System.Each sample reaction was ran in triplicate and the expression quantified as the number of cycles(CT)after which fluorescence exceeds the background threshold minus the CT for the housekeeping control(ΔCT).The calculation of absolute mRNA levels was based on the PCR efficiency and the threshold cycle(Ct)deviation of unknown cDNA versus the control cDNA.The quantitative values were obtained from the Ct values,which were the inverse ratios relative to the starting PCR product.The linear amount of target molecules relative to the calibrator was calculated by 2-ΔΔCT.Briefly,the relative levels of each mRNA were expressed as the same as above.
SNP identification and association analysis
Tree variation sites were identified in intronic of chicken genes ACSBG2,FASN and ELOVL6;and one variation site was identified as non synonymous of chicken ACSBG2 and synonymous coding region of chicken DDT gene by using GENBANK(Table 1).
The data for association study were analyzed by ANOVA(SAS 8.1).The statistical significance threshold was set at P<0.05.Values were expressed as the mean± SEM,and the differences in the means were compared using Duncan’s Multiple Range Test at 5%level of significance.
Results
Differentially expressed genes in fast-growing WRR and slow-growing XH chickens at 8 wk of age
After normalization and statistical analyses,400 and 220 genes with at least 2-fold differences were identified(P<0.05,FC≥2)in liver and hypothalamus tissues of WRR and XH chickens,respectively.When fast-growing WRR chickens were compared with slow-growing XH chickens,214 and 91 genes were up-regulated,and 186 and 129 genes were down-regulated in liver and hypothalamus tissues(Figure 1A and B;Tables 2 and 3).
In the liver,lipid metabolism genes viz SULT1B1, ACSBG2,LPL,AACS,PNPLA3,wereup-regulated while AOAH gene was down-regulated.The carbohydrate metabolism genes∶MGAT4B,XYLB,GBE1,PGM1, and HKDC1,were up-regulated(Table 2;Figure 1A). The fatty acid biosynthesis genes,ELOVL6 and FASN, cholesterol biosynthesis genes,LSS,HMGCR,FDPS, DHCR24,malate metabolism process gene,ME1,proline biosynthesis process genes,PYCR2 and ALDH18A1, oxidation-reduction reactions genes,CYP1A4,CYP1A1 similar to aldose reductase,AKR1B1,CYP4V2,and DDO, cyclic nucleotide catabolic process gene,N4BP2L1,and multicellular organism development genes,SEMA5A and
C1orf107,were identified highly expressed in WRR chickens.In contrast,genes highly expressed in XH chickens were associated with multicellular development,FRZB,immune response,DNA integration,melanin biosynthetic process,DDT,muscle organ development,DMD,transforming growth factor beta receptor signaling pathway, FUT8,and oxidation-reduction,CYP2C45,DHRSX, MICAL1,and CYP2C18.In addition,the genes for the biosynthesis of steroids and fatty acid,ELOVL6,and FASN were also observed highly expressed in XH chickens (Table 2;Figure 1).The metabolic process genes,ACSM5 (hypothetical protein),were down-regulated by 5-fold, while another metabolic process genes,ENPEP,were upregulated by 5-fold(Table 2).
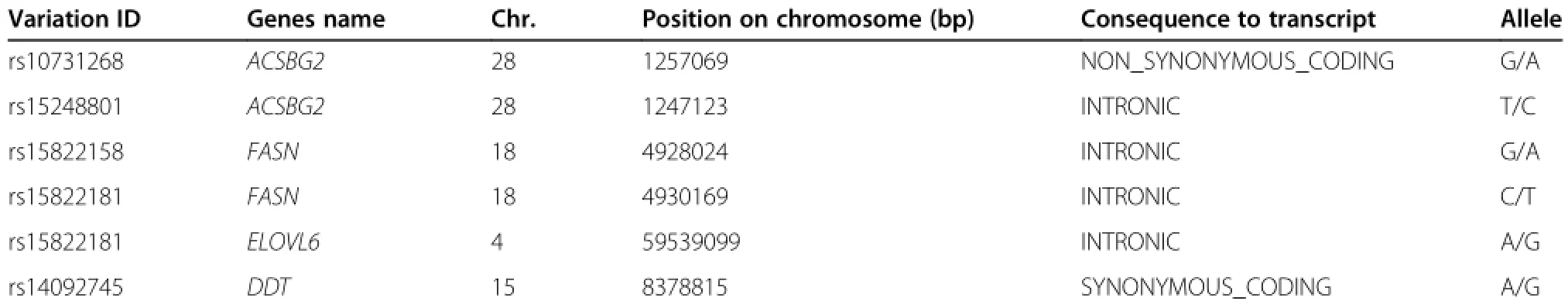
Table 1 The identified SNPs of the 4 fat deposition related genes
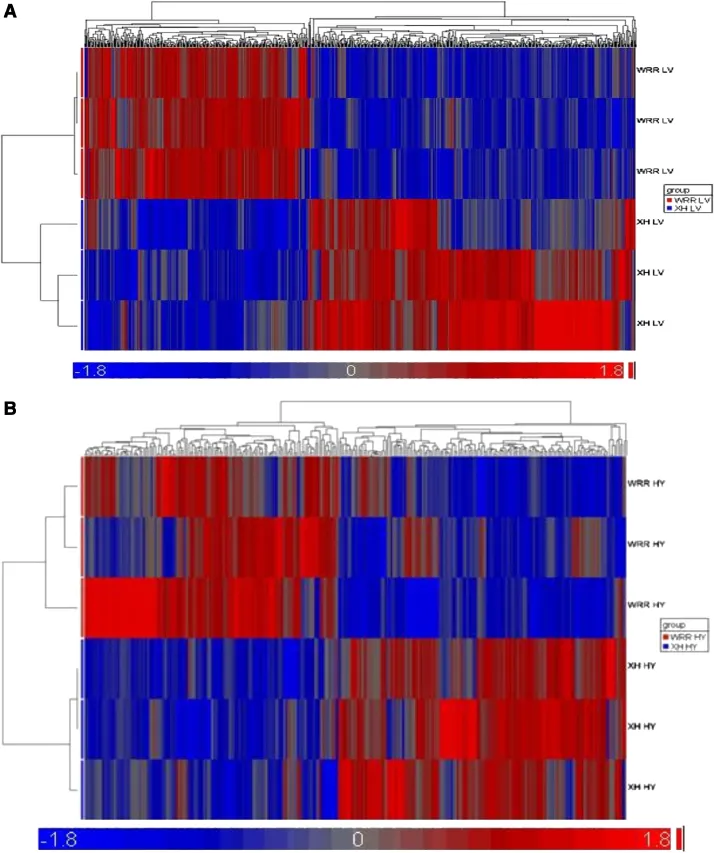
Figure 1(See legend on next page.)
In the hypothalamus,the cyclic nucleotide catabolism gene,N4BP2L1,was up-regulated in fast growing WRR chickens by a 3.7-fold change.The negative regulation of endothelial cell proliferation gene,TNFSF15,was upregulated by a 2.5-fold change.The proteolysis gene, ITGBL1,the protein amino acid phosphorylation genes, SGK1 and RIPK2,are up-regulated in the WRR chickens. The copper ion transport gene,SLC31A1,was localized on chr17∶1874758-1884555,and was up-regulated in the WRR chickens by a 7.3-fold change.The PODXL,RAD54B, PODXL,PMP2 and TMSB10,were up-regulated in the WRR chickens.The melanin biosynthesis gene,DDT,ion transport genes,SLC22A2,and GLRA1,lipid metabolism process gene,P20K(also known as EX-FABP),cellular amino acid metabolism gene,LOC772201,protein complex gene,ATPAF1,proteolysis genes,FOLH1, C1R,and VSP13B,striated muscle contraction gene, MYBPC2,nitrogen compound metabolism process gene,Vanin1,porphyrin biosynthesis process gene, FECH,and response to stress genes,HSP70,HSP25, and HSPB1,were down-regulated in slow-growing XH chickens.In addition,the signal transduction genes, similar to KIAA0712 protein,and ANK2,small GTPase mediated signal transduction gene,RAB30,DNA integration gene,LOC770705,amino acid phosphorylation gene,PRKD3,carbohydrate metabolic process gene, CBR1,and NAT13,neuron migration gene,MDGA1, hemophilic cell adhesion gene,PCDH24,sodium transport gene,SLC13A5,regulation of transcript DNA-dependent genes,CREB3L2,and MLL3,were also down-regulated in slow-growing XH chickens(Table 3).
Different gene ontology(GO)terms for biological process were identified in the livers of WRR and XH chickens.The highest GO clustered was in lipid biosynthesis process and fatty acid metabolism process (Figure 2).
In hypothalamus tissue,the GO terms for biological process in the WRR and XH chickens were mostly observed in response to stimulus,response to stress,and response to abiotic stimulus.Pigment metabolic process, melanin metabolic process,response to radiation,response to heat,response to temperature stimulus, leucocyte proliferation,pigment biosynthesis process, lymphocyte proliferation,mononuclear cell proliferation and response to ionizing radiation were also observed(Figure 3).
In the pathway study,a number of lipid-related genes∶ACSBG2,FASN,LPL,GPAM,FDPS,and others were identified.The cicardian clock gene,ARNTL also known as Bmal1,was observed.
Based on the pathways,differentially expressed genes participated in several function related to lipid(Tables 2 and 4).The lipid related genes were ACSBG2,SULT1B1, and LDLR of lipid metabolism,LPL of glycerolipid metabolism,and MTTP of lipid transporter activity,FASN and ELOVL6 of biosynthesis of unsaturated fatty acids, LSS,HMGCR,NSDHL,DHCR24,IDI1,of HSD17B7 of biosynthesis of steroid,AGPAT4 and FRZB of triacylglyceride synthesis,GPAM of glycerolipid metabolism,PHOSPHO1 and PTDSS1 of glycerophospholipid metabolism, ATP6V1C2 of oxidative phosphorylation,ACSS2 of glycolysis,GCNT2 of glycosphingolipid biosynthesis-lactoseries, and ME1 of pyruvate metabolism(Figure 3).
In hypothalamus tissue,three genes related to VEGF signaling pathway,four genes related to MAPK signaling pathway,one gene each related to alpha-linolenic acid metabolism,nitrogen metabolism,linoleic acid metabolism,porphyrin and chlorophyll metabolism were identified.Then a homologous recombination,heparan sulfate biosynthesis,ether lipid metabolism,arginine and proline metabolism,arachidonic acid metabolism,N-glycan biosynthesis,glycerophospholipid metabolism,ErbB signalingpathway,Wnt signaling pathway were also observed in our present study(Table 5).
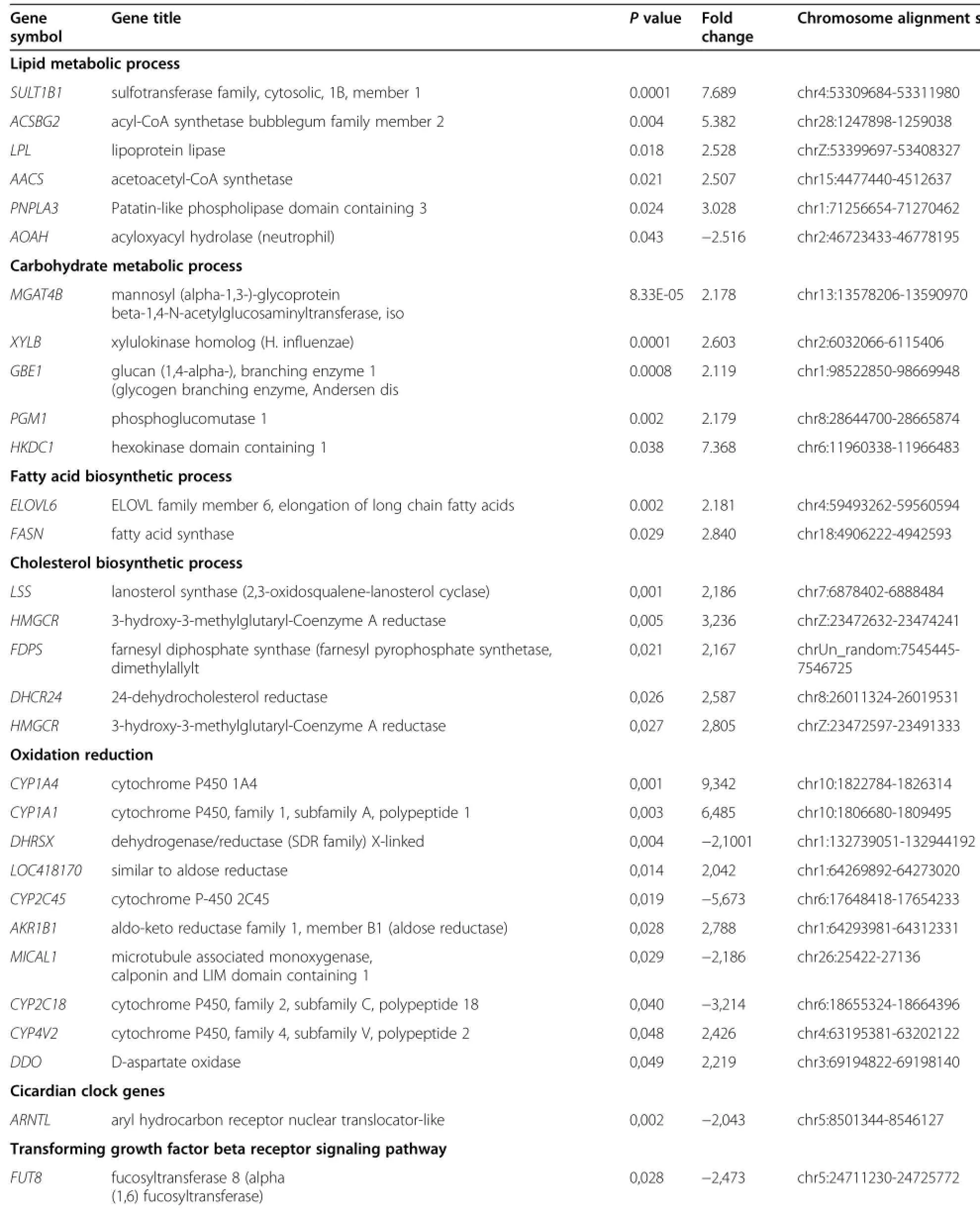
Table 2 Fold-changes of significantly differential expressed genes in WRR and XH chickens

Table 2 Fold-changes of significantly differential expressed genes in WRR and XH chickens(Continued)
Validation of differential expression by real-time RT-PCRThe mRNA levels of 9 genes involved in fat deposition were further quantified using real-time RT-PCR (Table 6).The level of 18S rRNA was chosen as reference and confirmed to be invariable.The expression levels(normalized to 18S)of the 9 genes were determined.Fold changes of gene expression determined by real-time RT-PCR were compared with the fold changes obtained from microarray analysis(Table 6).The highest fold changes in WRR chickens compared with XH chickens were confirmed in the SULT1B1,ACSBG2,ELOVL6, SLC31A1,and PNPLA3 genes.The lowest fold-changes were observed in the DDT and BEAN genes.
Expression levels of the Fat deposition related genes in the Fat tissues of WRR and XH chickens
When WRR males were compared with XH males,the expression of the LPL,FDPS,PNPLA3,GPAM,and SULT1B1 genes were up-regulated,and the FASN, ACSBG2,and FRZB were down-regulated in the abdominal fat tissue(Figure 4).In the subcutaneous fat tissue, the LPL,FDPS,PNPLA3,and SULT1B1 were upregulated,and the FASN,GPAM,ACSBG2,and FRZB genes were down-regulated.In the breast muscle tissues, theFDPS,PNPLA3,GPAM,andFRZBwereupregulated,and the LPL,FASN,ACSBG2,SULT1B1,and ELOVL6 genes were down-regulated(Figure 4).In the pituitary tissues,the LPL,FASN,SULT1B1,and ELOVL6 geneswereup-regulated,andtheFDPS,PNPLA3, GPAM,ACSBG2,and FRZB genes were up-regulated (Tables 7,4 and 5).
Polymorphisms of fat deposition genes associated with fat trait in chickens
The SNP rs10731268 of the ACSBG2 gene was associated with abdominal fat weight(P=0.005),and abdominal fat percentage(P=0.022).The SNP rs15248801 of the ACSBG2 gene was associated with abdominal fat weight(P=0.039)[Table 8].The SNP rs15822158 of the FASN gene was associated with fat band width(P= 0.0003),abdominalfatpercentage (P=0.001),and abdominal fat percentage(P=0.005)[Table 9].The SNP rs15822181 of the FASN gene was associated with abdominalfatweight(P=0.049)while the SNP rs16418687 of the ELOVL6 gene was associated with subcutaneous fat(P=0.034).The SNP rs14092745 of the DDT gene was associated with fat band width(P=0.048) [Table 9,10].
Discussion
The approach of selective-fat-deposition-related-genes in animals is a relatively new strategy aimed at improving production efficiency while enhancing meat quality.Efforts to reduce fat deposition in animals include genetic selection,feeding strategies,housing and environmental strategies as well as hormone supplementation.While these efforts have improved production efficiency and reduced carcass lipid deposition,negatively impacts on meat quality were due to reduced intramuscular fat deposition[22].Based on the comparison of two types of breeds of chicken whose fat deposition and growth rate are exceptionally varied,a functional genomics approach was chosen in order to identify chicken fat-depositionrelated-genes.In this genomic approach,liver tissue was used.The liver is the site of fat synthesis,and hypothalamus,which is a major gland for the endocrine system. Few studies focused on global gene expression surveys in chickens.Wang et al.[19]provided analysis of chicken adipose tissue gene expression profile.Other hepatic transcriptional analyses had been reported,using dedicated chicken 3.2 K liver-specific microarray[14,23] or a 323 cDNA microarray[24].
Differential gene expressions in liver during the fat developmental stage in fast growing WRR and slow growing XH chickens were related to lipid metabolism in our study.It has been reported that some genes,e.g.3-hydroxyacyl-CoA dehydrogenase,long chain acyl-CoA thioesterase,fatty-acid elongation enzymes and cytosolic fatty-acid-and acyl-CoA-binding proteins,are known to play key roles in mammalian fat or lipid metabolism [25].Glyco-metabolism such asglycol-sphingolipids (GCNT2),biosynthesis of steroids,fatty acid biosynthesis (ELOVL6 and FASN)was observed in this study.Collin et al.[26]reported that fast growing chickens developed excessive adiposity besidesthe high muscle massresulting from selection.The suggestion is that differential expression of the lipid metabolism related genes might be one of the factors in the differences of fat deposition between fast growing and slow growing chickens at the developmental stage.
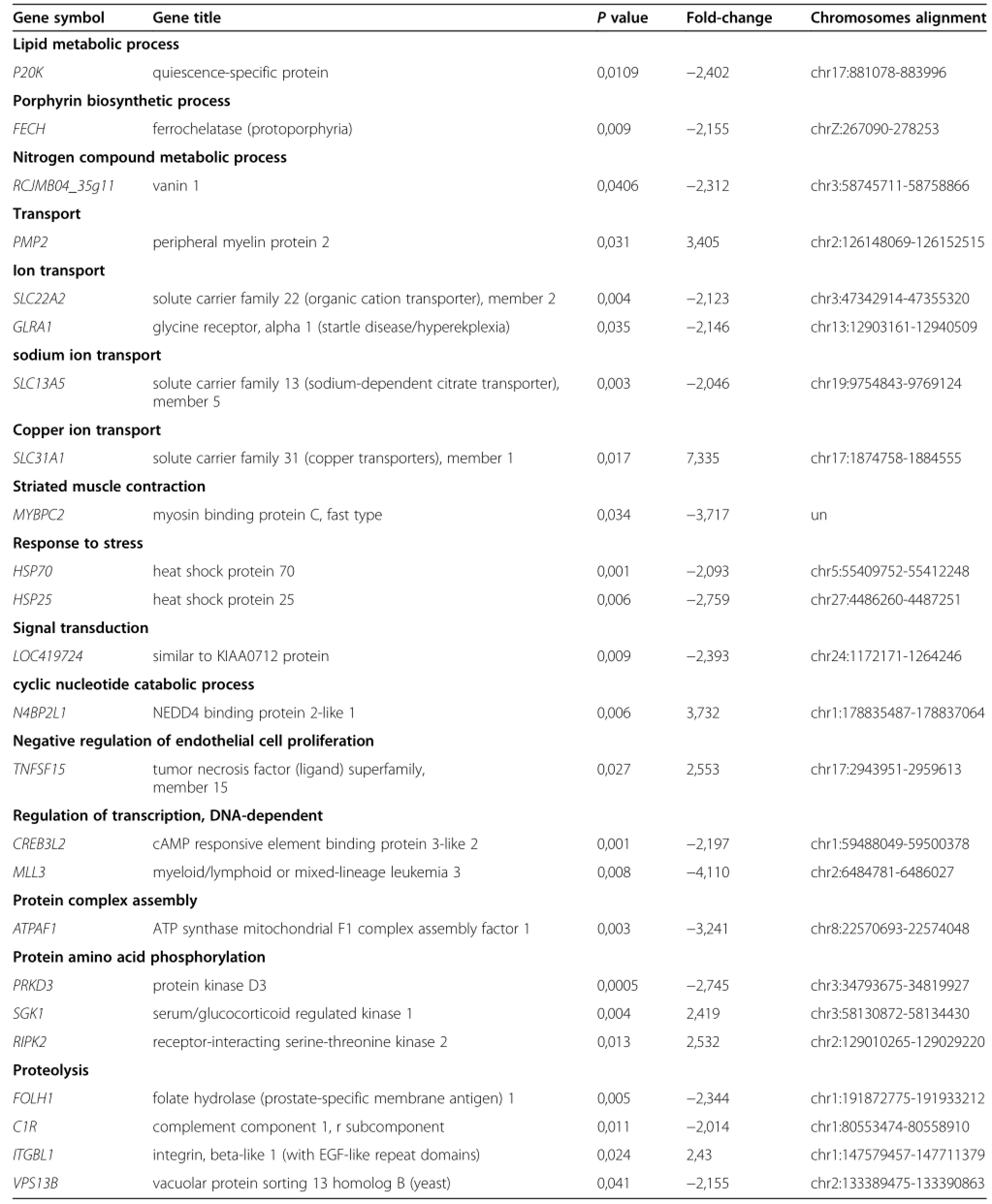
Table 3 Differentially expressed genes in hypothalamus of WRR and XH chickens
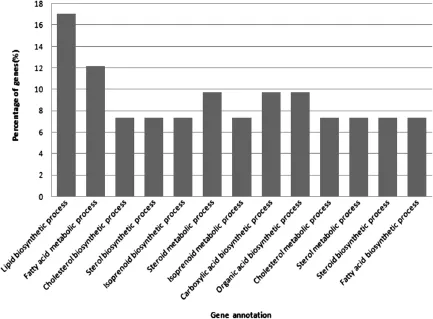
Figure 2The functional distribution of gene clusters of liver biological process.
The liver is the main site for fatty acid biosynthesis and the fatty acids are then transported to the adipose tissue for storage.The tasks are accomplished through the generation of triglycerides by the liver from fatty acids and L-a-glycerophosphate, packaged into very low density lipoproteins(VLDL), and then,secreted into the blood.The triglycerides in VLDL are processed by the adipose tissue and finally deposited in the centralvacuole ofthe adipocyte.It was suggested that several mechanisms regulate intracellular non-esterified fatty acids composition,including fatty acid transport,acyl CoA synthetases,fatty acid elongases,desaturases,neutral and polar lipid lipases and fatty acid oxidation.Most of these mechanisms are regulated by PPAR alpha or SREBP-1c.Together,these mechanisms control hepatic lipid composition and affect whole-body lipid composition [27].LPL catalyzes the hydrolysis of plasma lipoproteins,which is a rate-limiting step in
the transportation of lipids into peripheral tissues [28,29].The LPL gene expression in fast growing chicken was 2.5-fold greater than that in the slow growing type at the developmental stage in this study.In mammals,increased LPL activity is strongly associated with fat deposition and obesity,and these are regulated by both insulin and glucocorticoids according to Fried et al.[30].The major site of lipogenesis in birds,however,is the liver rather than the adipose tissue[31].The role of fatty acid-binding protein in the intramuscular trafficking of long-chain fatty acids within intramuscular adipocytes has been studied and found to be related to intramuscular levels in different species[32,33].
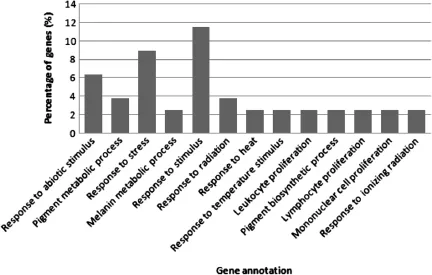
Figure 3The functional distribution of gene clusters of hypothalamus biological process.
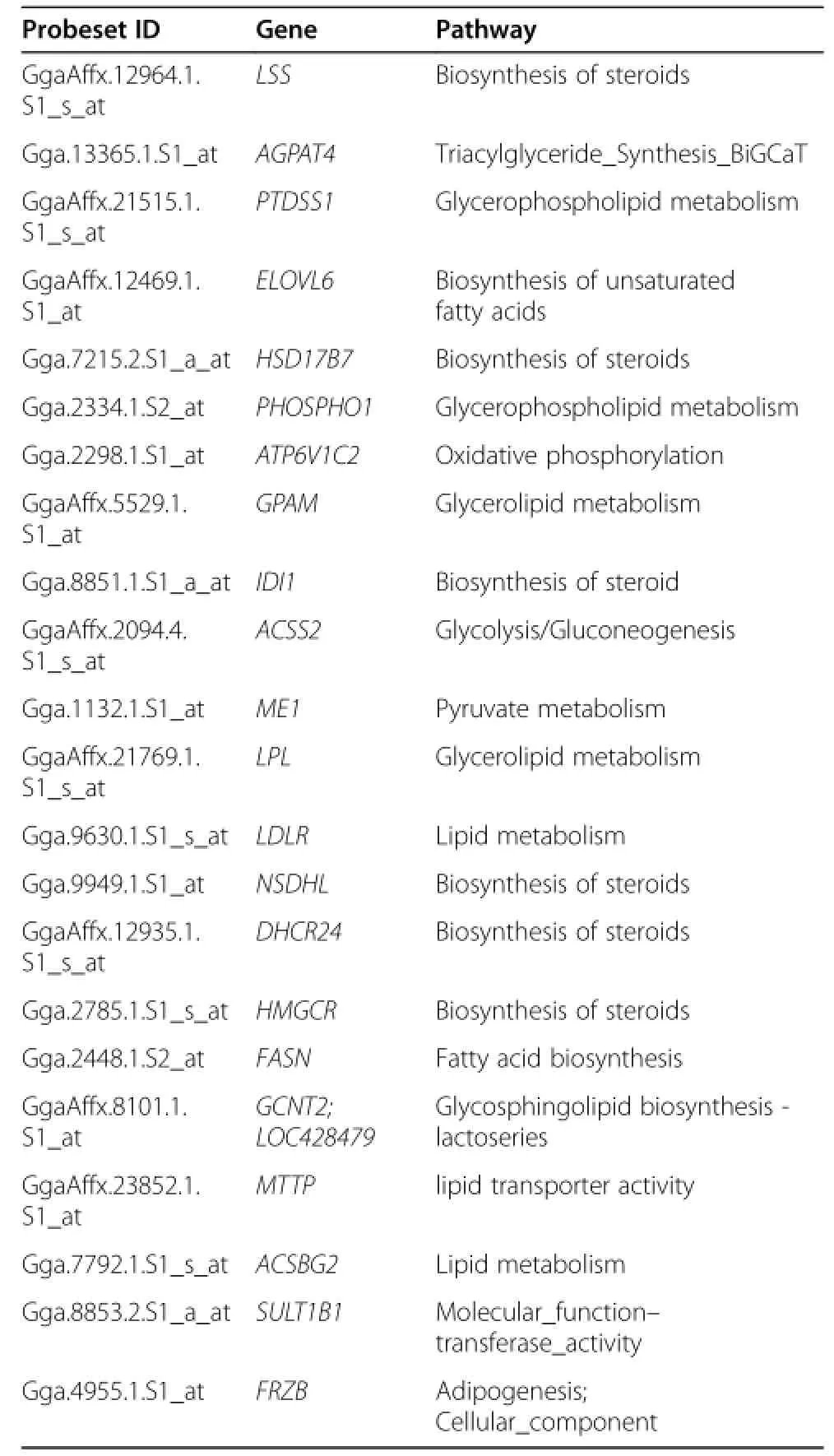
Table 4 Pathways of the fat-deposition-related genes in the liver of WRR and XH chickens
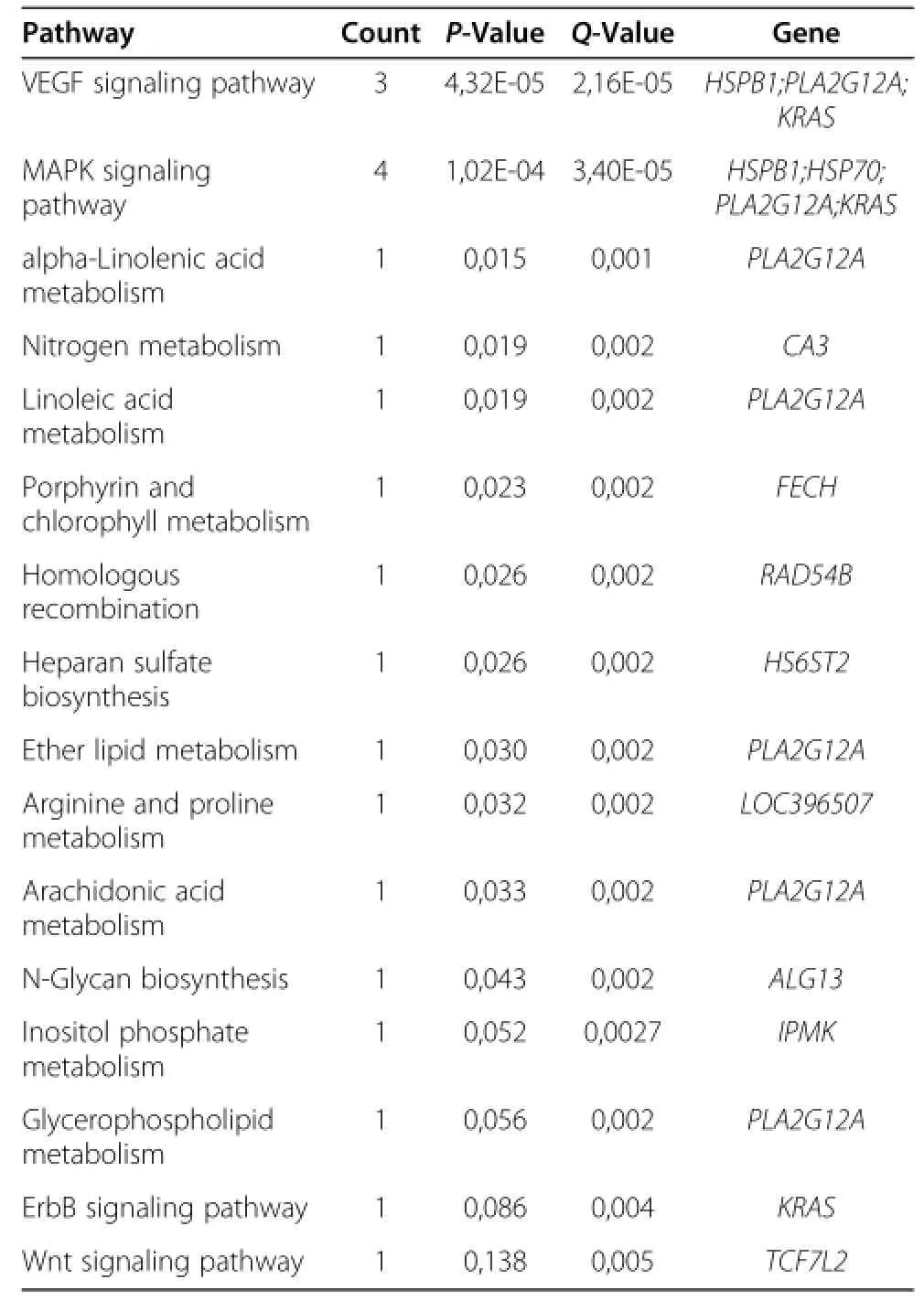
Table 5 Pathway of the fat-deposition-related genes expressed in hypothalamus tissue of WRR and XH chickens
Fatty acid synthesis(FAS)occurs during periods of energy surplus and concomitantly its gene expression is down-regulated during starvation in the liver[34], which is the major site of lipogenesis in avian species[35-37].The regulation of hypothalamic fatty acid synthesis gene expression in response to starvation is similar to that of liver fatty acid synthesis.In birds,like in humans,fatty acid synthesis primarily occurs in the liver.Demeure et al.[38]reported that chicken FASN gene is directly the target of liver cross receptor(LxR)alpha and therefore,expands the role of LxR alpha as a regulator of lipid metabolism. FASN and GPAM are two enzymes that play central roles in de novo lipogenesis.The G4928024A of the FASN gene is significantly associated with fat band width,abdominal fat percentage,and abdominal fat percentage.
The DDT gene was observed down-regulated in both tissues when fast growing WRR chickens were compared with slow growing XH chickens.This genehas function in melanization which can play a role in the pigmentation of abdominal fat.It also,has a high correlation with the accumulation of melanin in the skin of the shanks.Melanization of abdominal fascia is not harmful but it may cause severe economic losses to the producer.It was surprising to observe that the FDPS,LSS,HMGCR,NSDHL,DHCR24, IDI1,and HSD17B7 were up-regulated in fast growing WRR chickens.These genes are considered as the ones which has some functions in cholesterol biosynthesis.The glycolytic genes(ACSS2),carbohydrate metabolic and fatty acid biosynthesis were also up-regulated in the WRR chickens.It is suggested that the genes related to cholesterol biosynthesis,carbohydrate metabolic and fatty acid biosynthesis may have influence on fat development.
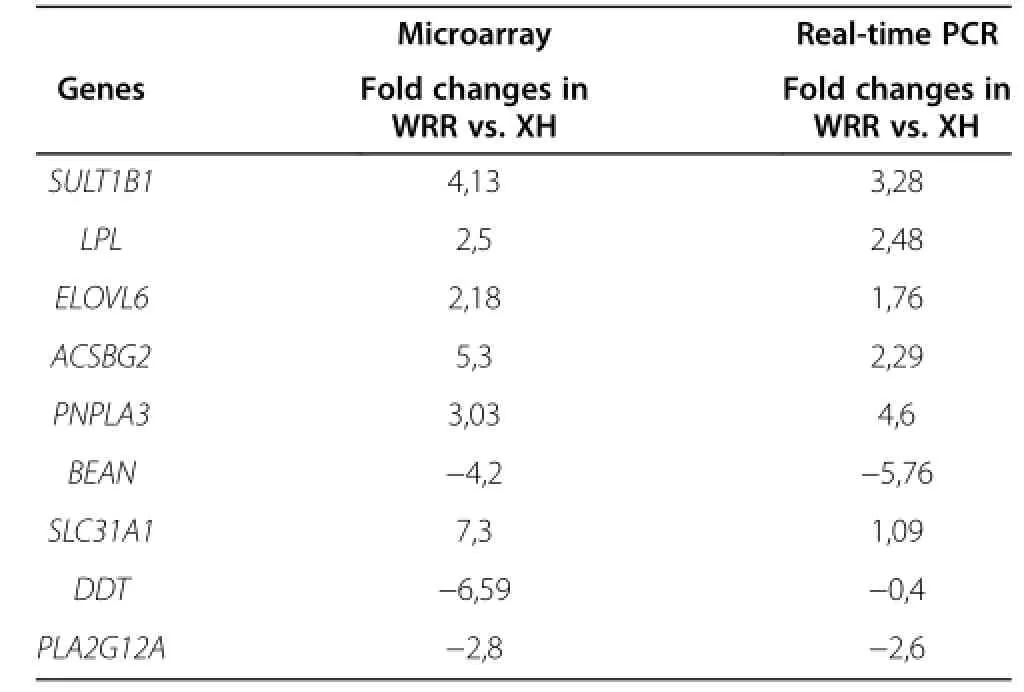
Table 6 Comparison of liver tissue gene expression levels between microarray and qRT-PCR
This study also showed that the genes related to proline biosynthetic process,member 2 of pyrroline-5-carboxylate reductase family,member A1 of aldehyde dehydrogenase 18 families,and oxidation reduction, CYP1A4,CYP1A1,AKR1B1,CYP4V2,DDO,and similar to aldose reductase,were differently expressed between the WRR and XH chickens.The CYP2H1,CYP2C45, CYP2C18,MICAL1 and CYP3A37 genes were significantly different in between the WRR and XH chickens. In this study,many lipid-related genes were identified, ACSBG2,FASN,LPL,GPAM,and FDPS.The cicardian clock gene(ARNTL)was observed,it plays a role in glucose,lipid metabolism and adipogenesis[39-41]. Moreover,a network of 11 genes,LPL,ACSBG2, AACS,FASN,LSS,FDPS,SULT1B1,HMGCR,DPP4, FUT8,and PLAU,was observed.Parallel expression patterns of these functionally relevant genes provided strong evidence for their coordinated involvement in lipid biosynthesis,cholesterol biosynthesis and fatty acid degradation in chickens.In chickens,the ACSBG2 gene has been found to play a significant role in lipid metabolism.The present study confirmed this conclusion.

Figure 4Data presented indicate the different mRNA gene relative expressions(as fold changes)of FG and SG male chicken relative to different fat tissues.Error bars represent the standard errors.Positive values imply genes were up-regulated in fast-growing chicken compared to slow-growing ones.
Table 7 The localization of differentially expressed fat-related genes

Table 7 The localization of differentially expressed fat-related genes(Continued)
In order to support the results of the microarray study,all the genes used for the mRNA assay were found to have good relationship with fat-related genes as their functions related to lipid metabolism, cholesterol biosynthesis and fatty acid metabolism. Interestingly,the SULT1B1,PNPLA3,GPAM,ELOVL6, LPL,FASN,ACSBG2,FDPS,and FRZB genes were preferentially expressed in 4 fatty tissues of abdominal fat,subcutaneous fat,breast muscle and pituitary gland when WRR werecompared with XH chickens. However,PNPLA3 mRNA level was higher in all tissues in WRR except pituitary tissue,where it expressed lower levels in XH chicken.PNPLA3,also referred to as adiponutrin,was originally identified as a highly adipose-specific transcript that rapidly responds to nutritional status[42].The microarray assay demonstrated that there was a 3 times higher expression of the PNPLA3 gene in liver tissue at 8 wk of age in WRR than in XH chickens.It could be concluded that the PNPLA3 gene is involved in fat deposition.
The microarray data showed that the SULT1B1 is abundantly expressed in liver tissue with 7 fold change in WRR with XH chickens.The gene was reported to be expressed in liver and other numerous extrahepatic tissues[43].FDPS is an important intermediate in cholesterol and sterol biosynthesis,a substrate for protein farnesylation and geranylgeranylation,and a ligand or agonist for certain hormone receptors and growth receptors.In this study,the FDPS was found to belong to the cholesterol biosythetic group.The FDPS mRNA level was higher in subcutaneous fat and pituitary tissue of WRR female chicken against XH counterpart.
The GPAM gene plays a vital role in the regulation of cellular triacylglycerol and phospholipid levels[44,45]. In this study,adipose tissues such as abdominal fat and subcutaneous fat were found to have the highest levels of GPAM mRNA expression whereas it was rarely detectable in the liver in the microarray assay.The FRZB gene(also known as SFRP3)is a member of the secreted frizzled receptor family of soluble proteins which binds to and antagonises Wnt receptor[46]. Wnts are secreted lipid-modified signaling proteins that influence multiple processes in the development of animals.The FRZB was shown to play a major role in adipogenesis in the microarray analysis of WRR and XH at 8 wk of age.ELOVL6 is involved in de novo lipogenesis and is regulated by dietary,hormonal and developmental factors[47].In this study,ELOVL6 mRNA level was higher in all tissues of XH chickens than of WRR chickens.
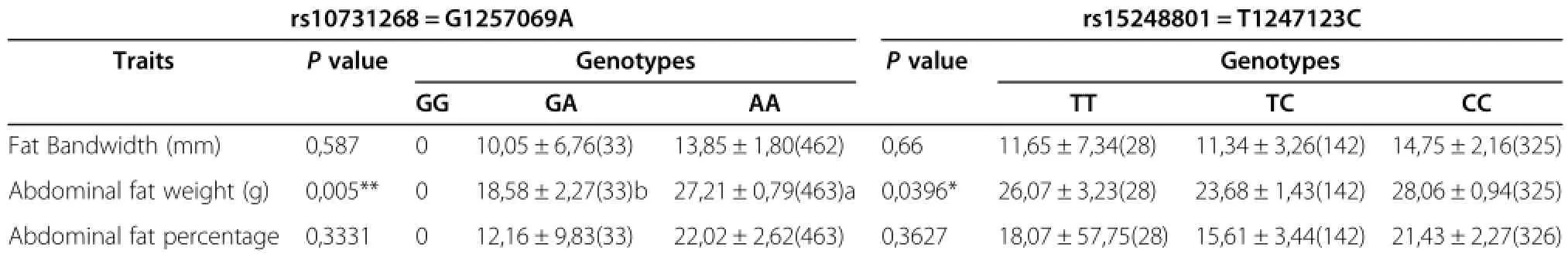
Table 8 Association of the G127069A,T1247123C in the ACSBG2 gene with chicken fat traits
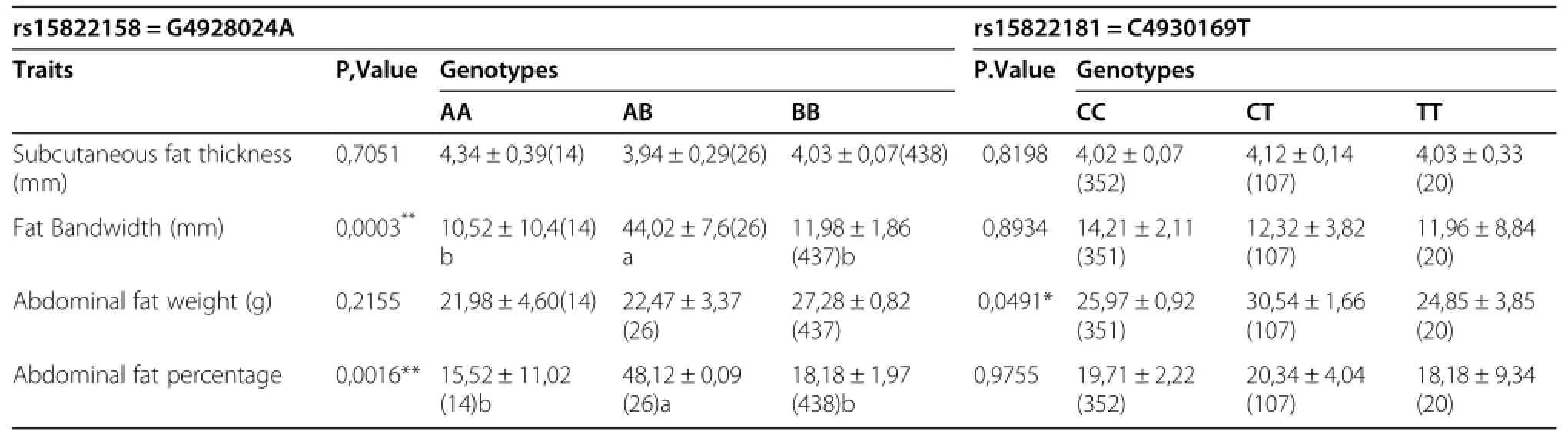
Table 9 Association of the G4928024A,C4930169T in the FASN gene with chicken fat traits
LPL is a glycoprotein enzyme that is produced in several tissues of mammals such as adipose tissue,skeletal muscle, heart,macrophages and lactating mammary gland,but not in the liver of adults[48,49].In chickens,LPL hydrolyzes lipids in lipoproteins,such as those found in chylomicrons and very low-density lipoproteins(VLDL)into three free fatty acid molecules and one glycerol molecule[29,50-52]. In studying the deposition of fat in the abdominal fat pads of chicken,it has become clearer that LPL-catalyzed hydrolysis of triacylglycerol in adipose tissue is a ratelimiting step in fat accumulation[28].Therefore,the transport and incorporation of exogenous lipids,i.e.plasma VLDL lipoprotein and portomicron,are essential for the deposition of cytoplasmic triglycerides in abdominal adipose tissue.These are characteristics of lipid metabolism in avian species since lipogenic activity is much greater in the liver than in adipose tissue[28,53,54].This study showed that the LPL gene expression was significantly higher in fast-growing chickens than in slow-growing chickens.
The association study provides direct evidence of genes related to fat deposition.In our association study, the A59539099G of the ELOVL6 gene was significantly associated with subcutaneous fat.The A8378815G of the DDT gene was associated with fat band width.The C4930169T of the FASN gene was also found to be associated with abdominal fat weight.G1257069A and T1247123C of the ACSBG2 gene were significantly associated with fat traits.The above results further confirmed that the ELOVL6,DDT,FASN,and ACSBG2 genes are related to chicken fat deposition.
Conclusion
The differential genes expressions in fast and slow growing chickens show differences in fat developmental stage which is supported by lipid-related genes identified and characterized in these two types of chicken.The findings indicate that the variation of the ACSBG2,FASN,ELOVL 6,and DDT genes were significantly associated with fat deposition.

Table 10 Association of the A59539099G in the ELOVL 6 and the A8378815G in the DDT gene on chicken fat traits
Abbreviations
WRR∶White recessive rock;XH∶Xinghua;RMA∶Robust multi-array; MAS∶Molecule annotation system;KEGG∶Kyoto encyclopedia of genes and genomes;AgriGO∶GO analysis toolkit and database for agricultural community;DAVID∶Database for annotation,visualization and integrated discovery.
Competing interests
The authors declare that they have no competing interests.
Authors’contributions
HCA is a correspondence author,conducted all the experiments and written and approved the final manuscript.WP participated in data analysis and approved the final manuscript.SX participated in data collection,laboratory experiment and approved final manuscript.JX participated in data collection, laboratory experiment and approved final manuscript.ZR participated in data collection,laboratory experiment and approved final manuscript.SL corried out the data analysis and approved final manuscript.ZX guided in gene expression analysis and approved final manuscript.
Acknowledgments
Funds for this work were partly provided by The Ministry of Higher Education Student Financing Agency of Rwanda,China Scholarship Council and South China Agricultural University,Guangzhou,China.Further support was provided by the Major State Basic Research Development Program, China(project no.2006CB102107),and the National High Technology Research and Development Program of China(863 Program,project no. 2007AA10Z163).
Author details
1Rwanda Agriculture Board,Research Department,P.O Box 5016,Kigali, Rwanda.2Council for Scientific and Industrial Research(CSIR),Animal Research Institute,P.O.Box AH 20,Accra,Achimota,Ghana.3Department of Animal Genetics,Breeding and Reproduction,College of Animal Science, South China Agricultural University,Guangzhou,Guangdong 510642,China.
Received∶26 March 2013 Accepted∶30 October 2013
Published∶9 November 2013
1. Ricard FH,Rouvier R∶Etude de la composition anatomique du poulet.1. Variabilite de la repartition des differentes parties corporelle chez de coquelets“Bresse-Pile”.Anri Zoorerh 1967,16∶23.
2. Proudman W,Mellen J,Anderson DL∶Utilization of feed in fast-and slowgrowing lines of chickens.Poult Sci 1970,49∶961-972.
3. Leclercq B,Blum JC,Boyer JP∶Selecting broilers for low or high abdominal fat∶initial observations.Br Poult Sci 1980,21∶107-113.
4. Becker WA∶Genotypic and phenotypic relations of abdominal fat in chickens. Kansas City,Missouri∶Presented at the 27th Annual National Breeder’s roundtable;1978.
5. Huan-Xian C,Ran-Ran L,Gui-Ping Z,Mai-Qing Z,Ji-Lan C,Jie W∶Identification of differentially expressed genes and pathways for intramuscular fat deposition in pectoralis major tissues of fast-and slow-growing chickens. BMC Genomics 2012,13∶213.
6. Dunnington EA,Siegel PB∶Long-term divergent selection for eight-week body weight in white Plymouth rock chickens.Poultry Sci 1996,75∶1168-1179.
7. Zhao SM,Ma HT,Zou SX,Chen WH∶Effects of in ovo administration of DHEA on lipid metabolism and hepatic lipogenetic genes expression in broiler chickens during embryonic development.Lipids 2007,42∶749-757.
8. Zhao X,Mo DL,Li AN,Gong W,Xiao SQ,Zhang Y,Qin LM,Niu YN,Guo YX, Liu XH,Cong PQ,He ZY,Wang C,Li JQ,Chen YS∶Comparative analyses by sequencing of transcriptomes during skeletal muscle development between pig breeds differing in muscle growth rate and fatness.PLoS One 2011,6(5)∶e19774.1-18.
9. Havenstein GB,Ferket PR,Qureshi MA∶Carcass composition and yield of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets.Poult Sci 2003,82∶1509-1518.
10.Jakobsson A,Westerberg R,Jacobsson A∶Fatty acid elongases in mammals∶their regulation and roles in metabolism.Prog Lipid Res 2005,45∶237-249.
11.Li WJ,Li HB,Chen JL,Zhao GP,Zheng MQ,Wen J∶Gene expression of heart and adipocyte-fatty acid-binding protein and correlation with intramuscular fat in Chinese chickens.Anim Biotechnol 2008,19(3)∶189-193.
12.Carlborg O,Kerje S,Schütz K,Jacobsson L,Jensen P,Andersson L∶A global search reveals epistatic interaction between QTL for early growth in the chicken.Genome Res 2003,13(3)∶413-421.
13.Zhao SM,Ma HT,Zou SX,Chen WH,Zhao RQ∶Hepatic lipogenesis gene expression in broiler chicken with different fat deposition during embryonic development.J Vet Med 2007,54∶1-6.
14.Cogburn LA,Wang X,Carre W,Rejto L,Aggrey SE,Duclos MJ,Simon J, Porter TE∶Functional genomics in chickens∶development of integrated systems microarrays for transcriptional profiling and discovery of regulatory pathways.Comp Funct Genomics 2004,5∶253-261.
15.Richards MP,Poch SM,Coon CN,Rosebrough RW,Ashwell CM,McMurtry JP∶Feed restriction significantly alters lipogenic gene expression in broiler breeder chickens.J Nutr 2003,133∶707-715.
16.Goodridge AG,Ball EG∶Lipogenesis in the pigeon∶in vivo studies.Am J Physiol 1967,213∶245-249.
17.O’Hea EK,Leveille GA∶Lipogenesis in isolated adipose tissue of the domestic chick(gallus domesticus).Comp Biochem Physiol 1968,26∶111-120.
18.Cai Y,Song Z,Zhang X,Wang X,Jiao H,Lin H∶Increased de novo lipogenesis in liver contributes to the augmented fat deposition in dexamethasone exposed broiler chickens(Gallus gallus domesticus). Comp Biochem Physiol C Toxicol Pharmacol 2009,150(2)∶164-169.
19.Wang HB,Li H,Wang QG,Zhang XY,Wang SZ,Wang YX,Wang XP∶Profiling of chicken adipose tissue gene expression by genome array. BMC Genomics 2007,27(8)∶193-207.
20.NRC∶Nutrient requirements of poultry.9th edition.Washington,DC∶National Academy Press;1994.
21.Lei MM,Nie QH,Peng X,Zhang DX,Zhang XQ∶Single nucleotide polymorphisms of the chicken insulin-like factor binding protein 2 gene associated with chicken growth and carcass traits.Poult Sci 2005,84(8)∶1191-1198.
22.Dodson MV,Hausman GJ,Guan L,Du M,Rasmussen TP,Poulos SP,Mir P, Bergen WG,Fernyhough ME,McFarland DC,Rhoads RP,Soret B,Reecy JM, Velleman SG,Jiang Z∶Lipid metabolism,adipocyte depot physiology and utilization of meat animals as experimental models for metabolic research.Int J Biol Sci 2010,6∶691-699.
23.Duclos MJ,Wang X,Carre W,Rejto L,Simon J,Cogburn LA∶Nutritional regulation of global gene expression in chicken liver during fasting and refeeding.San Diego,CA∶Plant Animal Genom XII Conf;2004.
24.Bourneuf E,Herault F,Chicault C,Carre W,Assaf S,Monnier A,Mottier S, Lagarrigue S,Douaire M,Mosser J,et al∶Microarray analysis of differential gene expression in the liver of lean and fat chickens.Gene 2006, 372∶162-170.
25.Ashrafi K,Chang FY,Watts JL,Fraser AG,Kamath RS,Julie A,Gary R∶Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes.Nature 2003,421∶268-272.
26.Collin A,Swennen Q,Skiba-Cassy S,Buyse J,Chartrin P,Le Bihan-Duval E,Crochet S,Duclos MJ,Joubert R,Decuypere E,Tesseraud S∶Regulation of fatty acid oxidation in chicken(Gallus gallus)∶Interactions between genotype and diet composition.Comp Biochem Physiol 2009,Part B 153∶171-177.
27.Jump DB,Botolin D,Wang Y,Xu J,Christian B,Demeure O∶Fatty acid regulation of hepatic gene transcription.J Nutr 2005,135(11)∶2503-2506.
28.Sato K,Akiba Y,Chida Y,Takahashi K∶Lipoprotein hydrolysis and fat accumulation in Chicken adipose tissues are reduced by chronic administration of lipoprotein lipase monoclonal antibodies.Poult Sci 1999,78∶1286-1291.
29.Sato K,Sook SH,Kamada T∶Tissue distribution of lipase genes related to triglyceride metabolism in laying hens(Gallus gallus).Comp Biochem Physiol 2010,155(1)∶62-66.
30.Fried SK,Russell CD,Grauso NL,Brolin RE∶Lipoprotein lipase regulation by insulin and glucocorticoid in subcutaneous and omental adipose tissues of obese women and men.J Clin Invest 1993,92∶2191-2198.
31.Leveille GA∶In vitro hepatic lipogenesis in the hen and chick.Comp Biochem Physiol 1969,28∶431-435.
32.Benistant C,Duchamps C,Cohen-Adad F,Rouanet JL,Barre H∶Increased in vitro fatty acid supply and cellular transport capacities in cold-acclimated ducklings (Cairina moschata).Am J Physiol 1998,275∶R683-R690.
33.Saez G,Davail S,Gentès G,Hocquette JF,Jourdan T,Degrace P,Baéza E∶Gene expression and protein content in relation to intramuscular fat content in Muscovy and Peki ducks.Poult Sci 2009,88∶2382-2391.
34.Back DW,Goldman MJ,Fisch JE,Ochs RS,Goodridge AG∶The fatty acid synthase gene in avian liver.Two mRNAs are expressed and regulated in parallel by feeding,primarily at the level of transcription.J Biol Chem 1986,261∶4190-4197.
35.Goodridge AG,Ball EG∶The Effect of Prolactin on Lipogenesis in the Pigeon.In Vitro Studies.Biochemistry 1967,6(8)∶2335-2343.
36.Leveille GA,O’Hea EK,Chakrabarty K∶In vivo lipogenesis in the domestic chicken.Proc Soc Exp Biol Med 1968,128∶398-401.
37.Leveille GA,Romsos DR,Yeh YY,O’Hea EK∶Lipid biosynthesis in the chick. A consideration of site of synthesis,influence of diet and possible regulating mechanisms.Poult Sci 1975,54∶1075-1093.
38.Demeure O,Duby C,Desert C,Assaf S,Hazard D,Guillou H,Lagarrigue S∶Liver X receptor regulates fatty acid synthase expression in chicken. Poult Sci 2009,88∶2628-2635.
39.Shimba S,Ishii N,Ohta Y,Ohno T,Watabe Y,Hayashi M,et al∶Brain and muscle arnt-like protein-1(BMAL1),a component of the molecular clock, regulates adipogenesis.Proc Natl Acad Sci U S A 2005,102∶12071-12076.
40.Gómez-Santos C,Gómez-Abellán P,Madrid JA,Hernández-Morante JJ,Lujan JA,Ordovas JM,Garaulet M∶Circadian rhythm of clock genes in human adipose explants.Obesity 2009,17(8)∶1481-5.30.
41.Wu X,Zvonic S,Floyd ZE,Kilroy G,Goh BC,Hernandez TL,et al∶Induction of circadian gene expression in human subcutaneous adipose-derived stem cells.Obesity 2009,15(11)∶2560-2570.
42.Baulande S,Lasnier F,Lucas M,Pairault J∶Adiponutrin,a transmembrane protein corresponding to a novel dietary and obesity-linked mRNA specifically expressed in the adipose lineage.J Biol Chem 2001,276∶33336-33344.
43.Dooley TP,Haldeman-Cahill R,Joiner J,Wilborn TW∶Expression profiling of human sulfotransferase and sulfatase gene superfamilies in epithelial tissues and cultured cells.Biochem Biophys Res Commun 2000,277∶236-245.
44.Yazdi M,Ahnmark A,William-Olsson L,Snaith M,Turner N,Osla F,Wedin M, Asztély AK,Elmgren A,Bohlooly-Y M,Schreyer S,Lindén D∶The role of mitochondrial glycerol-3 phosphate acyltransferase-1 in regulating lipid and glucose homeostasis in high-fat diet fed mice.Biochem Biophys Res Commun 2008,369∶1065-1070.
45.Palou M,Priego T,Sánchez J,Villegas E,Rodríguez AM,Palou A,Picó C∶Sequential changes in the expression of genes involved in lipid metabolism in adipose tissue and liver in response to fasting.Pflugers Arch 2008,456(5)∶825-836.
46.Marsit CJ,Houseman EA,Christensen BC,Gagne L,Wrensch MR,et al∶Identification of methylated genes associated with aggressive bladder cancer.PLoS One 2010,5(8)∶e12334.
47.Kan S,Abe H,Kono T,Yamazaki M,Nakashima K,Kamada T,Akiba Y∶Changes in peroxisome proliferator-activated receptor gamma gene expression of chicken abdominal adipose tissue with different age,sex and genotype.Anim Sci J 2009,80∶322-327.
48.Hoenig M,McGoldrick JB,De Beer M,Demacker PNM,Ferguson DC∶Activity and tissue-specific expression of lipases and tumor-necrosis factor a in lean and obese cats.Domest Anim Endocrinol 2006,30∶333-344.
49.Albalat A,Saera-Vila A,Capilla E,Gutierrez J,Perez-Sanchez J,Navarro I∶Insulin regulation of lipoprotein lipase(LPL)activity and expression in gilthead Sea bream(sparus aurata).Comp Biochem Physiol B 2007,148∶151-159.
50.Sato K,Akiba Y∶Lipoprotein lipase mRNA expression in abdominal adipose tissue is little modified by age and nutritional state in broiler chickens.Poultry Sci 2002,81∶846-885.
51.Nie Q,Fang M,Xie L,Shi J,Zhang X∶cDNA cloning,characterization,and variation analysis of chicken adipose triglyceride lipase(ATGL)gene. Mol Cell Biochem 2009,320∶67-74.
52.D’André Hirwa C,Yan W,Wallace P,Nie Q,Luo C,Li H,Shen X,Sun L,Tang J,Li W,Zhu X,Yang G,Zhang X∶Effects of the thyroid hormone responsive spot 14 {alpha}gene on chicken growth and fat traits.Poult Sc.2010,89(9)∶1981-1991.
53.Griffin HD,Hermier D∶Plasma lipoprotein metabolism and fattening on poultry.In Leanness in domestic birds.Edited by Leclercq B,Whitehead CC. London∶Butterworths;1988∶175-201.
54.Griffin HD,Guo K,Windsor D,Butterwith SC∶Adipose tissue lipogenesis and fat deposition in leaner broiler chickens.J Nutr 1996,122∶363-368.
doi∶10.1186/2049-1891-4-43
Cite this article as:Claire D’Andre et al.∶Identification and characterization of genes that control fat deposition in chickens.Journal of Animal Science and Biotechnology 2013 4∶43.
Submit your next manuscript to BioMed Central and take full advantage of:
· Convenient online submission
· Thorough peer review
· No space constraints or color fi gure charges
· Immediate publication on acceptance
· Inclusion in PubMed, CAS, Scopus and Google Scholar
· Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
*Correspondence∶chirwa02@yahoo.fr
1Rwanda Agriculture Board,Research Department,P.O Box 5016,Kigali, Rwanda
3Department of Animal Genetics,Breeding and Reproduction,College of Animal Science,South China Agricultural University,Guangzhou,Guangdong 510642,China
Full list of author information is available at the end of the article
©2013 Claire D’Andre et al.;licensee BioMed Central Ltd.This is an open access article distributed under the terms of the Creative Commons Attribution License(http∶//creativecommons.org/licenses/by/2.0),which permits unrestricted use, distribution,and reproduction in any medium,provided the original work is properly cited.
Results:Four hundred genes in the liver tissues and 220 genes hypothalamus tissues,respectively,were identified to be differentially expressed in fast-growing chickens and slow-growing chickens.Expression levels of genes for lipid metabolism(SULT1B1,ACSBG2,PNPLA3,LPL,AOAH)carbohydrate metabolism(MGAT4B,XYLB,GBE1,PGM1, HKDC1)cholesttrol biosynthesis(FDPS,LSS,HMGCR,NSDHL,DHCR24,IDI1,ME1)HSD17B7 and other reaction or processes(CYP1A4,CYP1A1,AKR1B1,CYP4V2,DDO)were higher in the fast-growing White Recessive Rock chickens than in the slow-growing Xinghua chickens.On the other hand,expression levels of genes associated with multicellular organism development,immune response,DNA integration,melanin biosynthetic process,muscle organ development and oxidation-reduction(FRZB,DMD,FUT8,CYP2C45,DHRSX,and CYP2C18)and with glycol-metabolism (GCNT2,ELOVL 6,and FASN),were higher in the XH chickens than in the fast-growing chickens.RT-PCR validated high expression levels of nine out of 12 genes in fat tissues.The G1257069A and T1247123C of the ACSBG2 gene were significantly associated with abdominal fat weight.The G4928024A of the FASN gene were significantly associated with fat bandwidth,and abdominal fat percentage.The C4930169T of the FASN gene was associated with abdominal fat weight while the A59539099G of the ELOVL 6 was significantly associated with subcutaneous fat.The A8378815G of the DDT was associated with fat band width.
Conclusion:The differences in fat deposition were reflected with differential gene expressions in fast and slow growing chickens.
 Journal of Animal Science and Biotechnology2014年1期
Journal of Animal Science and Biotechnology2014年1期
- Journal of Animal Science and Biotechnology的其它文章
- GMOs in animal agriculture∶time to consider both costs and benefits in regulatory evaluations
- Dual potential of microalgae as a sustainable biofuel feedstock and animal feed
- Haplotype phasing after joint estimation of recombination and linkage disequilibrium in breeding populations
- Comparative analysis of proteomic profiles between endometrial caruncular and intercaruncular areas in ewes during the peri-implantation period
- Changes in feed intake,nutrient digestion,plasma metabolites,and oxidative stress parameters in dairy cows with subacute ruminal acidosis and its regulation with pelleted beet pulp
- Estimation of endogenous protein and amino acid ileal losses in weaned piglets by regression analysis using diets with graded levels of casein