
Bio-Argo浮标观测贫营养水体浮游植物的光适应特性*
2014-03-18 09:22邢小罡丘仲锋
激光生物学报 2014年6期
邢小罡,丘仲锋,王 林
(1.中国海洋大学 物理海洋重点实验室,山东 青岛266003;2.南京信息工程大学 海洋科学学院,江苏 南京210044;3.国家海洋环境监测中心 遥感室,辽宁 大连116023)
海洋浮游植物的光合作用,不仅是海洋生态系统中最重要的过程,而且在大时空尺度上调节着大气中CO2的含量以及影响着整个地球的气候变化,在全球碳循环中同样扮演着非常重要的作用。在过去的20多年里,全球海洋初级生产力的估算及其时空变化,一直是推动海洋光学研究以及水色遥感发展的重要动力,也是海洋生态学、海洋生物地球化学以及全球气候变化研究中关注的重点[1-5]。
由于叶绿素a是浮游植物细胞内参与光吸收与光合作用的主要色素,因此,叶绿素a浓度([Chla])一直以来都被看作表征浮游植物生物量的关键性指标以及估算初级生产力的主要参量[3,5,6],也因此成为海洋光学观测的基本参数与水色遥感反演的基本产品[7,8]。然而,浮游植物生理学的相关研究已证明,单位浮游植物细胞内的叶绿素a含量会随着光照、温度、营养盐、藻种类型以及其他环境因子而发生变化[9-11]。特别是在环境光照的变化过程中,叶绿素a浓度与浮游植物生物量(通常用C含量来表征)的比值(Chla:C)会发生显著的生理学响应,表现为随光照的负相关关系,称为光适应(Photoadaptation)或 光 驯 化(Photoacclimation)[10-12]。于 此 同 时,海洋生物光学的研究也发现,一些浮游植物的固有光学参量(Inherent Optical Properties,IOPs)--如颗粒物后向散射系数(bbp)和颗粒物光束衰减系数(cp)--与颗粒有机碳(particulate organic carbon,POC)的丰度在局地海区存在很高的相关性[13-15],并已应用到卫星数据估算POC[16,17]。上述研究不仅证明了这些颗粒物的固有光学量可以作为比叶绿素a浓度更好的生物量指标(大洋水体中,浮游植物碳是POC的主要贡献者),其与叶绿素a的比值还可以作为浮游植物生理学的指标(一阶近似的情况下正相关于Chla:Cphy)[18-20]。
在这些研究的背景下,Behrenfeld等通过现场数据模拟了营养盐充足的情况下Chla:Cphy与光照的关系,即Chla:Cphy=A+Bexp(-CIg),其中A、B、C是回归系数,均为正值,Ig称为生长光照)。然而,与该模型相关的几个问题尚未证实。首先,Behrenfeld等基于第2次和第3次大西洋经向断面(Atlantic Meridional Transect)航次AMT-2、AMT-3的观测数据确定系数C为3[21],但其回归分析中使用的仅是营养盐充足的表层数据。在贫营养水体(营养盐不足)中浮游植物的光适应是否也满足此方程尚不明确。此外,AMT航次是在整个大西洋经向上进行的断面观测,每次历时仅为1个月左右,浮游植物的光适应性随光照的变化来源于混合层深度的海区差异(动力条件不同)和表层光照的纬度差异(太阳高度不同)。然而在局地海区中,浮游植物的光适应性则是受混合层与光照的季节变化共同作用导致的,Chla:Cphy与光照在时间尺度上的变化关系是否仍然满足其在空间尺度上建立的模型,也需要更多的现场观测数据加以验证或修正。
然而,受观测平台的限制,目前可用于验证局地海区(特别是贫营养海区)浮游植物光适应性的现场数据非常稀少。船基走航和锚系浮标是目前进行现场观测的主要平台。船基走航虽然可以满足一定空间覆盖的需要,但难以给出完整的季节变化;锚系浮标虽然可以满足任何时间尺度上的观测需要,但一般只能设立在近岸海区,远离人类活动的大洋水体观测非常稀少,且一般没有混合层深度的观测数据。最近,一种新型的海洋自动观测平台Bio-Argo浮标的出现,为海洋光学和生物地球化学的现场观测带来了革命性的技术飞跃[23,24]。它可以在局地海区进行长期的自动化观测(1-2年),且具有很高的垂向分辨率(1 m)。此外,在Bio-Argo浮标上物理参数与生物光学参数是同步测量的,因此特别适用于局地海区生物光学与生物地球化学季节变化过程的分析以及生态-物理耦合过程的研究[25-28]。本文利用2008-2010年在南、北太平洋副热带海区以及西北地中海的4台Bio-Argo浮标观测数据,分析上述三个贫营养水体中混合层内叶绿素a浓度与颗粒物光束衰减系数的季节变化规律及其比值(Chla:cp)与生长光照之间的关系,以期揭示贫营养水体中浮游植物的光适应规律以及不同海区的局地性差异。
1 材料与方法
1.1 浮标数据
图1显示了4台浮标数据所观测的三个海区,方框表示浮标在观测期内运动的范围。其中,南太平洋副热带海区SPG(South Pacific Gyre)的2台浮标(浮标编号PAC_SO_B04、PAC_SO_B07),从2008年12月3日开始观测分别运行至2010年3月6日和1月30日;北太平洋副热带海区NPG(North Pacific Gyre)的1台浮标(浮标编号PAC_NO_B08),有效观测数据从2008年8月15日至2010年8月10日;西北地中海NWM(NorthWestern Mediterranean sea)的1台浮标(浮标编号MED_NW_B02),有效观测数据从2008年5月11日至2010年2月8日。
4台浮标上均携带有美国Wetlabs公司生产的Eco-Puck传感器(用于观测叶绿素a荧光、黄色物质荧光与532 nm后向散射系数)和C-Rover透射计(用于观测660 nm光束衰减系数)以及加拿大Satlantic公司生产的OC4辐射计(用于观测412、490、555 nm下行辐照度)。
叶绿素荧光数据首先经过偏移校正(offset correction),偏移量取为每次剖面观测的最小值(一般出现在350-400 m),再通过基于光照的校正方法确定校正斜率[29],最终得到校正后的叶绿素a浓度([Chla],单位mg m-3)。660 nm光束衰减系数的处理也需要进行偏移校正,同时去除了纯海水的衰减系数(常数)和(由于颗粒物沉降在传感器表面而产生的)仪器观测误差,得到颗粒物衰减系数(cp(660),单位m-1)。此外,混合层深度(Mixed Layer Depth,MLD)通过密度剖面计算。为去除海洋表层(<10 m)密度的昼夜波动,以10 m处的密度值作为参考值,密度增加的阈值选择为0.03 kg m-3[30]。
1.2 卫星数据与推导数据
与浮标观测位置时间同步的海表面日有效光合辐照度(Photosynthetically Available Radiance,PAR,单位mol quanta m-2d-1)来自于美国NASA提供的Aqua卫星中分辨率成像光谱仪MODIS(MODerate resolution Imaging Spectroradiometer)遥感观测的8天平均PAR数据,分辨率9 km×9 km。同步卫星数据选择为浮标观测时所在位置±0.1°区间内所有卫星观测值(一般4-9个)的平均值,用PAR(0)表示。考虑到日照时长对浮游植物光适应性的影响,一般将日有效光合辐照度的单位转化为mol quanta m-2h-1(除以当天的日照小时数,单位h/d)[21,22]。
混合层内的生长光照参照Behrenfeld等[22]提出的:
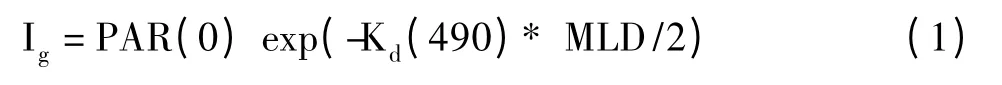
其中,Kd(490)称为490 nm的光衰减系数,通过叶绿素a浓度与Kd(490)的经验关系[31]估算得到:
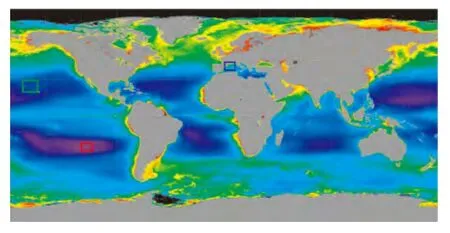
图1 用于本文研究的Bio-Argo浮标在三个贫营养水体的观测区域(SPG、NPG分别代表南北太平洋副热带环流区,NWM代表西北地中海)Fig.1 Observation regions of the Bio-Argo float deployed in three oligotrophic waters(SPG,NPG represent the South and North Pacific subtropical Gyre,respectively.NWM represents the NorthWestern Mediterranean Sea)

这里的叶绿素a浓度取为混合层内的平均值。
1.3 光适应模型
本文在对浮游植物的光适应性进行参数化分析的过程中,采用了两种回归模型:

前一种模型是根据Behrenfeld等基于大西洋的现场观测数据提出的光适应关系(即公式4中的C被确定为3)[21-22],回归时直接采用线性回归方法,确定系数A和B。第二个模型则假定系数C是可变的,方程变为非线性,本文采用优化算法得到三个系数。为表述方便,后文将公式(3)称为3Ig模型,将公式(4)称为CIg模型。
2 结果与分析
2.1 混合层深度与叶绿素a极大值的季节变化
图2显示了4台浮标在三个贫营养海区观测的混合层深度MLD与叶绿素极大值(Deep Chlorophylla Maximum,DCM)所在的深度zDCM。三个海区的MLD都显示出类似的季节性变化,冬季加深、夏季变浅,其中SPG和NPG的最大MLD都超过120 m,NWM的MLD在冬季也超过100 m。对于叶绿素极大值的深度,zDCM似乎与MLD的变化过程相反:冬季变浅、夏季加深。其中,SPG的zDCM最深,最深超过180 m,最浅也达到150 m;NPG的zDCM次之,在90-120 m之间波动;NWM最浅,最深仅有80 m,但其变化的幅度非常剧烈。需要说明的是,在NPG与NWM海区,计算的zDCM在冬季会出现在混合层内(即zDCM<MLD),这一计算结果并非说明叶绿素a浓度在混合层内存在极大值,而是表明混合层在冬季的加深会破坏DCM现象。
值得注意的是,DCM在冬季的消失也表明,深层的营养盐会在混合作用下向表层补充。一些现场观测数据显示,zDCM的深度与营养盐跃层(nutricline)的深度存在一致性[32-33]。因此,zDCM与MLD的关系,也在一定程度上指示了营养盐向表层的补充情况。例如,在SPG海区,全年的zDCM均深于MLD,说明叶绿素a极大值的现象在该海区是常年存在的,且营养盐难以补充到表层,导致SPG成为全球最低的贫营养水体与极低叶绿素a浓度水体[34]。此外,虽然NPG与NWM在冬季都存在充分混合(wellmixed)期,但NWM在冬季的混合时间更长(约4个月),因而营养盐在冬季的补充将更丰富;而NPG每年仅有约1个月的充分混合期。
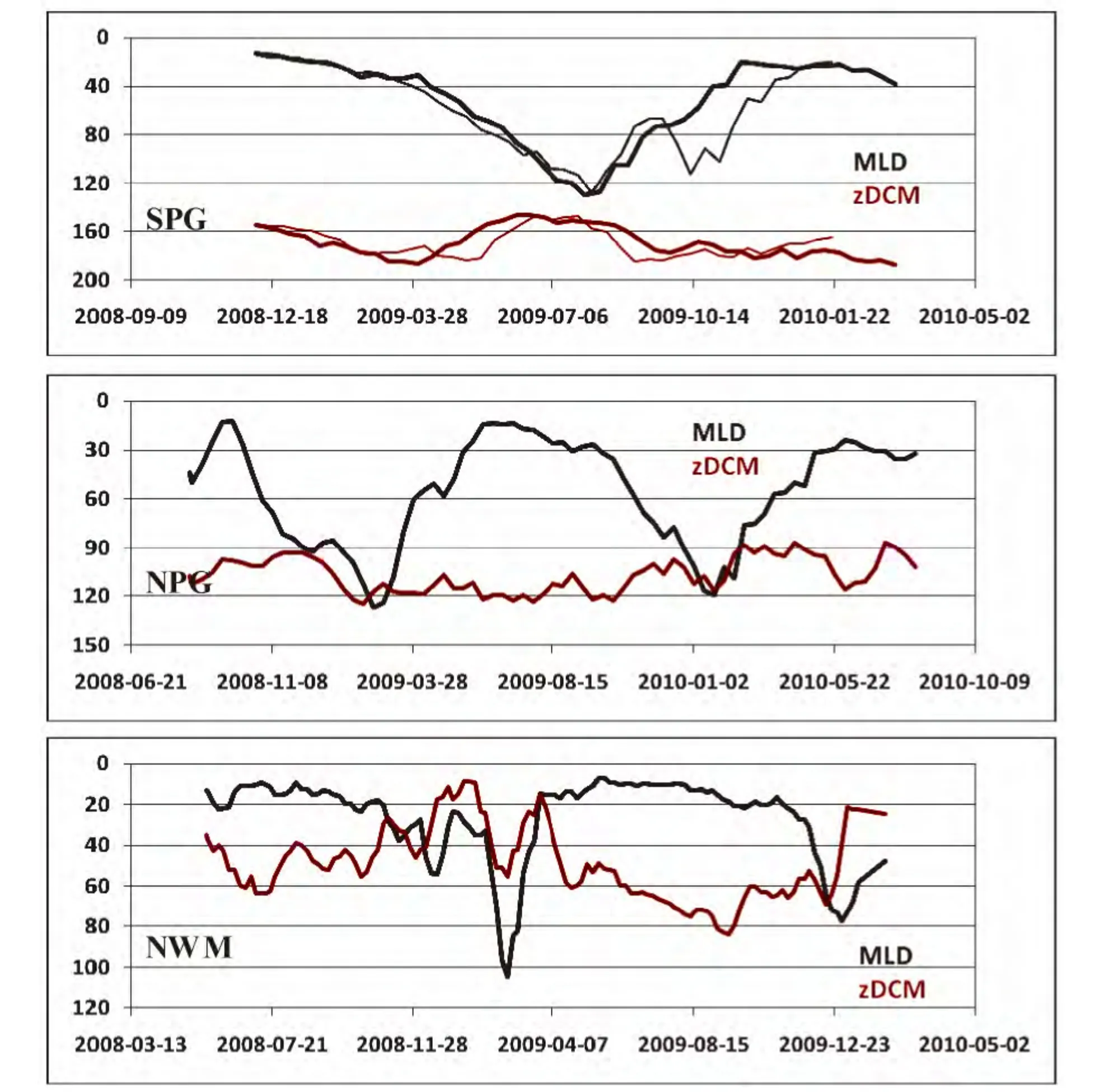
图2 三个贫营养海区观测的混合层深度(MLD)与叶绿素a极大值深度(zDCM)的时间序列,SPG图中粗线代表B04浮标、细线代表B07浮标观测数据Fig.2 Time series of the mixed layer depth(MLD)and the depth of deep chlorophyll-a concentration(zDCM)observed in three oligotrophic waters.In the SPG panel,the heavy and thin lines represent the data recorded by the B04 and B07 float,respectively
2.2 叶绿素a与光束衰减系数的季节变化
图3显示了三个贫营养海区观测的混合层内叶绿素a浓度[Chla]与光束衰减系数cp(660)的季节变化。对于SPG与NPG海区,[Chla]与cp(660)呈现出不同的季节变化趋势,cp(660)的季节性变化很小,而[Chla]则有显著的季节特征,冬季增高,夏季降低。SPG海区,cp(660)甚至呈现出微弱的与[Chla]相反的变化趋势:冬季稍低、夏季稍高。这与Behrenfeld等[22]通过卫星数据观测的颗粒物后向散射系数的结果非常一致:在副热带环流区的典型贫营养海区中,相比于叶绿素信号,浮游植物生物量的季节性变化非常微弱,甚至存在负相关关系。Siegel等[20]也基于卫星数据指出,在一些长期的或季节性的上升流区(如东赤道太平洋、东边界流、阿拉伯海等),浮游植物生物量一般与叶绿素共变。对于这些区域,叶绿素是很好的评价浮游植物生物量的指标;然而在西北太平洋和南大西洋以及南太平洋副热带环流区,叶绿素与浮游植物生物量间甚至存在反向的变化关系,说明叶绿素在这些海区的季节性波动实际上代表了浮游植物的生理学变化。
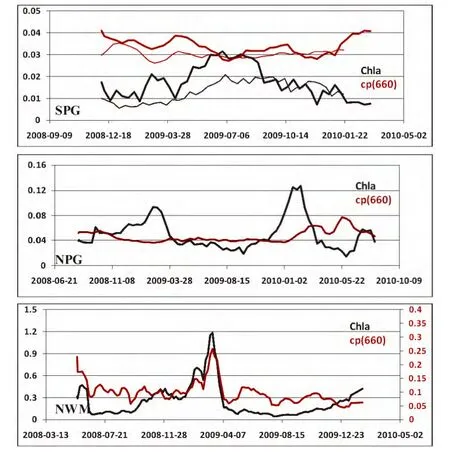
图3 三个贫营养海区观测的表层叶绿素a浓度([Chla])与光束衰减系数(cp(660))的时间序列Fig.3 Time series of the surface chlorophyll-a concentration([Chla])and the beam attenuation coefficient(cp(660))observed in three oligotrophic waters
在NWM海区,叶绿素a与衰减系数则呈现相似的季节形态。由于冬季强烈的混合作用,营养盐被输送到混合层,从而促进了冬季和春季的表层藻华。值得注意的是,[Chla]的增长要明显早于cp(660),叶绿素a浓度从秋季(9月)开始升高,而浮游植物直到冬季(12月)才出现爆发,说明在NWM海区浮游植物的光适应性仍然非常显著。出现上述现象的原因,本文推测主要由于秋季混合层的加深,浮游植物的生长光照降低,促进了浮游植物细胞内叶绿素a的增加(光适应),而此时由于混合作用尚未影响到营养盐跃层,表层的营养盐水平变化不明显,浮游植物的生物量也未显著增加。直到冬季,当混合作用将足够的营养盐带入表层后,浮游植物的生物量才出现了明显的增长。
综上所述,本文讨论的3个海区叶绿素a与颗粒物衰减系数都呈现出不同程度的非耦合关系。而造成这种非耦合关系的主要原因是浮游植物在随光照的季节性变化过程中不断调节着自身细胞内叶绿素a含量。

图4 三个贫营养海区观测的叶绿素a浓度和光束衰减系数的比值(Chla:cp)与生长光照(Ig)的时间序列Fig.4 Time series of the ratio of chlorophyll-a concentration to the beam attenuation coefficient(Chla:cp)and the growth irradiance(Ig)observed in three oligotrophic waters
2.3 叶绿素a与光束衰减系数的比值与生长光照的季节变化
图4显示出了三个海区的[Chla]与cp(660)的比值以及对应的生长光照Ig的季节变化过程。如上文所述,cp(660)通常作为POC的一个替代性参数,而由于在大洋水体中,浮游植物碳是POC的主要来源。因此,在一定程度上,Chla:cp可以表征浮游植物叶绿素a与碳比值(Chla:C),后者经常作为一个重要的浮游植物生理学参量以及光适应性指标[10,21]。可以看出,Chla:cp与生长光照Ig呈现出非常好的耦合关系。Chla:cp在夏季最低,冬季最高,与生长光照的变化过程刚好相反。
特别说明的是,由于在Ig的计算过程中需要[Chla]估算的Kd(490)。因此,本研究特别分析了计算的Ig与三个主要输入量(PAR(0)、MLD、Kd(490))的相关性,以保证Chla:cp与Ig的强耦合关系并非由于Ig与[Chla]的相关性导致的。根据公式(1),PAR(0)与Ig通过线性回归,MLD、Kd(490)与Ig通过指数函数回归确定其相关性。在SPG,三种输入量与Ig的相关系数r2分别为0.868、0.931和0.332;NPG海区得到的r2分别为0.772、0.924和0.595;NWM海区的三个输入量与Ig的r2分别为0.872、0.876和0.403。可以看出,虽然Kd(490)与Ig也存在一定程度的弱相关关系,但生长光照主要受表层光照和混合层深度的影响,特别是混合层深度的影响最大。
图5显示的是Chla:cp与Ig的散点图,并分别根据3Ig模型与CIg模型回归分析得到的两者关系曲线(回归公式与相关性r2见表1)。除SPG外,NPG与NWM海区两者的相关性都非常高,特别是CIg模型模拟的结果都在0.8左右。由于CIg模型比3Ig模型多一个可变的参数,因而得到的相关性更高。两种模型的回归参数在3个海区存在同样的差别,即3Ig模型的回归系数均高于CIg模型。从模型的数学本质上来说,回归系数A代表了高光条件下的最低比值(Chla:cp)min,回归系数B代表了高光与低光条件下比值的差,而A+B则代表了低光条件下的最高比值(Chla:cp)max。因此,上述差异表明,3Ig模型回归的最低比值与最高比值均高于CIg模型,这一结果通过图5显示的回归曲线也可以看出。
此外,比较不同海区的关系还可以发现,光适应性的区域性差异非常明显。在同等光照条件下,Chla:cp在NWM最高,SPG最低(NWM>NPG>SPG)。此外,对于CIg模型中回归的系数C,则刚好相反,SPG海 区 最 高(1.74),NWM海 区 最 低(0.81)。回归系数C与3的差异,也导致了两条回归曲线在NWM海区的差异性比较大,而在SPG海区则相对比较接近。简单来说,两者关系的区域性差异以及两种模型的结果差异,反映的是不同海区的营养盐和温度的限制程度[22]。
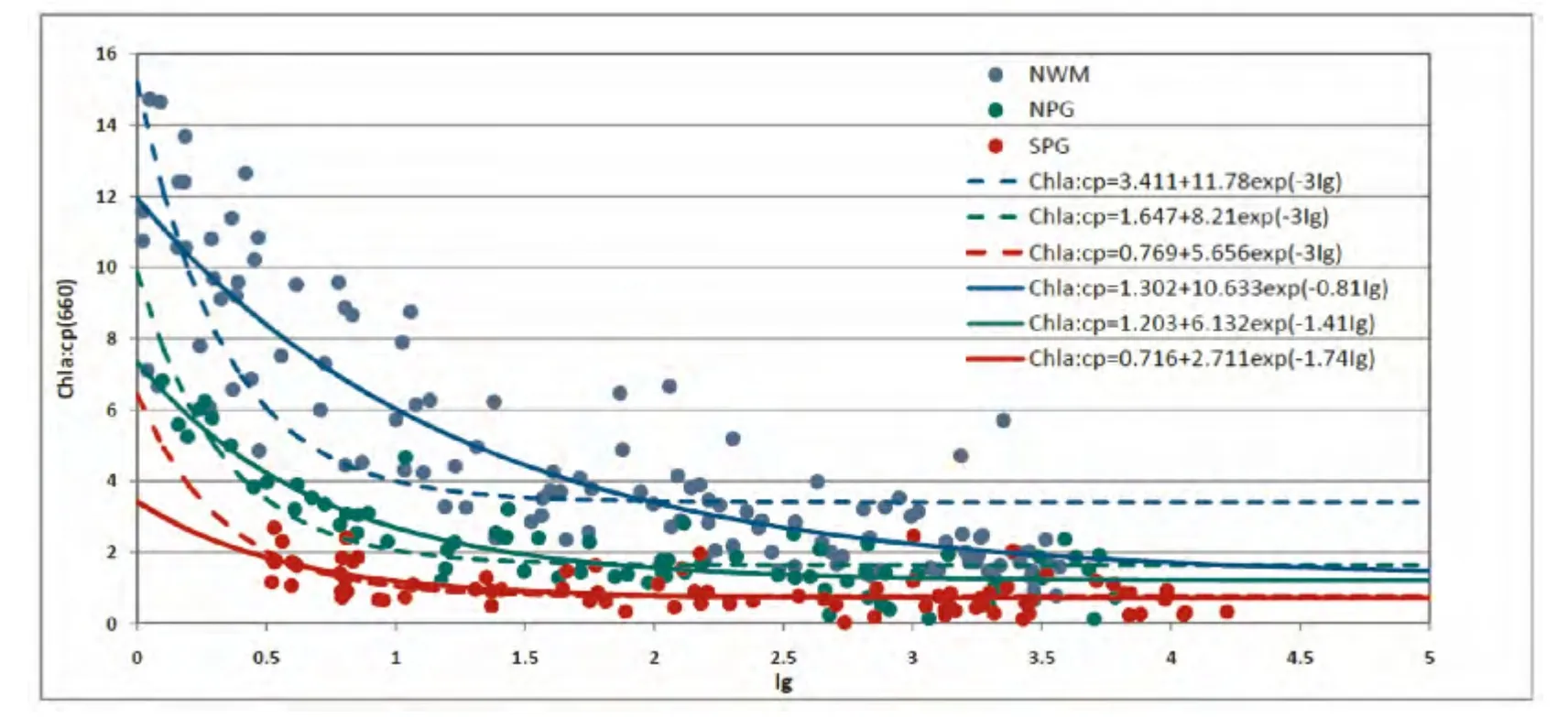
图5 三个贫营养海区观测的叶绿素a浓度和光束衰减系数的比值(Chla:cp)与生长光照(Ig)的散点图及其回归关系FIg.5 Scatter plot between the ratio of chlorophyll-a concentration to the beam attenuation coefficient(Chla:cp)and the growth irradiance(Ig)observed in three oligotrophic waters,as well as the regressed relationships

表1 现场观测的Chla:cp与Ig的回归分析结果
3 讨论
3.1 浮标结果与卫星估算结果的比较
由于光束衰减系数目前无法通过水色遥感反演,Behrenfeld等将颗粒物后向散射系数(bbp(440))作为浮游植物碳的替代性参数,并通过卫星数据进行了全球海洋的光适应性研究[22]。基于卫星数据的研究,目前主要存在两个问题:(1)从观测数据和米散射理论分析来看,光束衰减系数更能代表浮游植物的信号[35],而颗粒物后向散射系数似乎对小型浮游植物或浮游植物以外的小颗粒物(微生物)的信号响应更明显[36]。因此,卫星观测的Chla:bbp可能除生理学信息外,还携带了颗粒物粒径的信息,这可能对浮游植物的生理学研究造成一定的困难。(2)由于卫星无法实时监测混合层深度,因此生长光照的计算存在很大的难度(生长光照与混合层深度的相关性非常高,见2.3节)。Behrenfeld等人使用FNMOC(海军舰队数字化气象和海洋中心)模型模拟的混合层深度进行生长光照的估算[22],可能带来一定的误差。因此,本文在这里将现场浮标数据与卫星估算的结果进行比较,可以在一定程度上对卫星遥感的结果进行验证。由于所使用的数据并不相同(本文使用cp,Behrenfeld等使用bbp并将其转化为浮游植物碳[22]),因而本文仅定性地比较海区的相对差别,而无法直接定量比较回归系数的数值。
Behrenfeld等[22]基于3Ig模型得到的系数如表2所示,SP-L0海区(对应于本文的SPG)的Chla:Cmin与Chla:Cmax分别为0.0037和0.016,在全球28个大洋海区中都接近最低值;NP-L1海区(对应本文的NPG)的Chla:Cmin与Chla:Cmax分别为0.0053和0.031,均高于SPG;虽然他们未对地中海进行研究,但考虑到其海区特点大致相当于同纬度的北大西洋,NA-L2海区的Chla:Cmin与Chla:Cmax分别为0.0058和0.021。与图5比较可以看出,卫星数据与浮标数据表现出的海区差异比较相似,特别是Chla:Cmin的大小同样按照SPG<NPG<NWM顺序排列。但Chla:Cmax的顺序稍有不同,NWM>NPG的海区特点未有体现。导致这一差异的原因可能是NWM的区域特殊性。对于整个地中海,NWM比其他海区有更高的叶绿素a浓度但颗粒物的丰度(bbp)则差别不大[37],说明NWM比其他地中海区域拥有更高Chla:C水平,相应地Chla:Cmin与Chla:Cmax应该也更高。虽然无法直接比较回归系数,但可以通过比值法(即低光最高值/高光最低值)分析,消除cp与bbp的差别(假设两者都随浮游植物碳满足y=Ax的关系),因此可比较浮标观测的Chla:cp与卫星观测的Chla:bbp的回归结果,通过比较比值发现,在NPG与NWM海区浮标与卫星数据的结果非常接近,SPG的比值浮标结果偏高,可能是由于数据较少,Chla:C变化不大,相关性不高导致的(浮标与卫星数据都存在这一问题)。
综上所述,卫星与现场观测数据得到的结论基本一致,本文发现的海区差异在卫星数据中也有明显的体现。
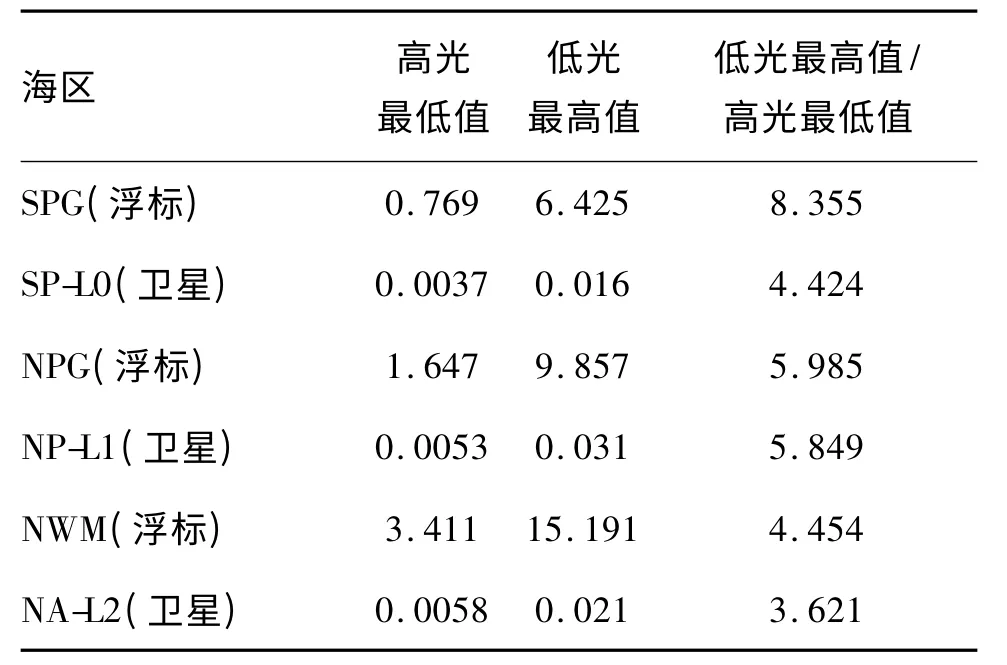
表2 浮标与卫星观测数据回归分析的高光最低值与低光最高值及其比值(对于浮标数据是Chla:cp;对于卫星数据是Chla:C)
3.2 光适应方程的生理学解释
关于上述的海区差异,Behrenfeld等根据前人的实验室和现场数据也指出,Chla:Cmin受营养盐限制的影响,营养盐限制越强,Chla:Cmin越低;Chla:Cmax则受温度限制的影响,高温对应着高的Chla:Cmax,低温将对叶绿素a产生限制导致Chla:Cmax的降低[22]。因此,方程(3)可以写为

这里用f(N)表示Chla:Cmin是营养盐的函数,g(T)表示Chla:Cmax是温度的函数。根据公式(5),我们可以推导出两个基本结论:
1)理论上存在一个不受营养盐和温度限制的最大Chla:C与光照的响应曲线,Behrenfeld等[22]基于此,将所有卫星数据的上廓线(包括了99%的数据点)找出:

所有受到营养盐或温度限制的点都将落在廓线之下,限制越大的海区,其观测点距离廓线越远。根据本文发现的海区差异推论,SPG所受的营养盐和温度的限制都最大,而NWM受到的限制最小。这一结论对于营养盐限制比较适用(从2.1节显示的MLD与zDCM的关系可以看出,SPG全年处于营养盐限制,而NWM在冬季和春季表层营养盐得到补充)。但对温度而言,NWM的年平均温度最低,Chla:Cmax应该较低,这一差异同上文所述,NWM海区存在一定的特殊性,Chla:C整体偏高,可能是由于藻种类型的差别导致的。
此外,由于公式(5)中营养盐和温度的限制是分离的,因此可以通过比较低光最高值与高光最低值的比值来评价哪种限制占主导。由于非限制模型(公式(6))中两者的比值是2.045(0.045/0.022),如果仅存在温度限制,将导致比值小于2.045(低光最高值下降);反之,如果仅存在营养盐限制,比值将高于2.045(高光最低值下降)。换言之,温度限制在降低这一比值,而营养盐限制在调高这一比值。因此,不同海区两者的比值就代表了营养盐与温度限制的比例(即哪种限制占主导)。比较表2的比值关系,可以看出,所有三个海区都是营养盐限制占主导因素,且SPG与NPG要高于NWM。
2)如果所有的数据点受到的温度或营养盐的限制水平始终保持不变,CIg模型与3Ig模型的模拟结果将趋于一致(即回归系数C=3)。换言之,如果营养盐或温度的限制在季节性上存在差异,将导致CIg模型的模拟结果偏离3Ig模型,即回归系数C将小于3。而且,这样的季节性差异越大,C越小。基于这一推论,我们可以根据回归结果推测,NWM海区浮游植物所受的营养盐或温度的限制在季节上变化最显著(C=0.81),而SPG海区营养盐和温度限制的变化率最小(C=1.74),这一推测同样可以从现场数据中得到印证。从营养盐的限制上可以看出,SPG全年处于营养盐限制,季节变化最小(虽然受到的限制最大),NWM在冬季和春季表层营养盐充足,夏季存在限制,因此季节变化最大。而温度的变化同样支持上述结论,各海区温度的季节变化为:NWM(10°C)>NPG(6°C)>SPG(5°C)。这就解释了回归系数C的生理学意义,它表征了营养盐或温度限制在季节上的变化幅度。
此外,3Ig模型与CIg模型回归结果的差别说明,3Ig模型是一种“评估”模型,用于评估该区域水体总体的营养盐与温度限制水平。将C限制为3(营养盐与温度非限制情况),即将Chla:C随光照限制的响应去除,仅留下营养盐与温度限制的响应。但其反演的参数Chla:Cmin和Chla:Cmax会同时高估真实的高光最低值和低光最高值(见表1和图5)。因为,在低光条件下,仅存在温度限制,营养盐限制很小;而在高光条件下,仅存在营养盐限制,温度限制又很小[22]。因此,如果对总体进行评估时,将同时低估两者的限制水平,导致3Ig回归曲线更向上并接近于非限制曲线(公式(6))。而CIg模型的结果则非常依赖于曲线两端,因此其回归参数所反映的情况是Chla:Cmin基本受高光情况的影响,而Chla:Cmax则基本受低光情况的影响,同时体现季节变化上的两种限制水平达到最大的情况,而系数C则是描述了这两种限制在季节变化上有多大差别。
综上所述,3Ig模型首先将光照限制去除,从而描述总体的温度和营养盐限制水平,但会同时高估真实的Chla:C最大和最小值,更适用于区域性分析;CIg模型的结果难以分离三种限制水平,但可以真实描述Chla:C的季节变化情况,体现了冬夏两季的实际差别,更适用于季节性分析。
4 结论
本文利用4台Bio-Argo浮标在三个贫营养海区的观测数据,分析了叶绿素a与光束衰减系数的季节变化,及其两者的比值与光照的响应关系,并通过两种回归模型获得了表征营养盐与温度限制程度的回归系数。分析结果发现,浮游植物的光适应性特征在局地的季节变化上也是普遍存在的,并存在一定的海区差异。SPG海区受营养盐的限制最大,NPG次之,NWM最小。主要由于三个海区的冬季混合时间(MLD>zDCM的时间)不同所致,营养盐的补充存在明显的区域性差异。CIg模型回归的系数C代表了营养盐或温度限制在季节上的变化幅度,对于变化幅度最大的NWM海区,C最小,而对于全年表层温度以及营养盐变化都不大的海区,C将趋近于3。这有可能为卫星数据分析提供一个新的参数,考察不同海区C的变化也有助于研究浮游植物的生理环境的季节性变化。值得注意的是,本研究发现NWM海区的Chla:C特征可能存在局地特殊性,对于这一现象可以通过卫星数据进一步加以验证与讨论。
致谢:感谢法国滨海自由城海洋学实验室(Laboratoire d’Océanographie de Villefranche)的Hervé CLAUSTRE教授为本研究提供的Bio-Argo浮标数据。
[1] MOREL A.Optical modeling of the upper ocean in relation to its biogenous matter content[J].Journal of Geophysical Research,1988,93(C9):10749-10768.
[2] ANTOINE D,ANDRÉJ M,MOREL A.Oceanic primary production:2.Estimation at global scale from satellite(coastal zone color scanner)chlorophyll[J].Global Biogeochem Cycles,1996,10(1):57-69.
[3] BEHRENFELD M J,FALKOWSKI P G.Photosynthetic rates derived from satellite-based chlorophyll concentration[J].Limnol Oceanogr,1997,42(1):1-20.
[4] CAMPBELL J,ANTOINE D,ARMSTRONGR,et al.Comparison of algorithms for estimating primary productivity from surface chlorophyll,temperature and irradiance[J].Global Biogeochem Cycles,2002,16(3):1035-1049.
[5]BEHRENFELD M J,WORTHINGTON K,SHERRELL R M,et al.Controls on productivity across the tropical Pacific ocean revealed through nutrient stress diagnostics[J].Nature,2006,442(7106):1025-1028.
[6] BOYCE D G,LEWIS M R,WORM B.Global phytoplankton decline over the past century[J].Nature,2010,466(7306):591-596.
[7] O’REILLY J E,MARITORENA S,MITCHELL B G,et al.Ocean color chlorophyll algorithms for SeaWiFS[J].J Geophys Res,1998,103(C11):24937-24953.
[8] MARITORENA S,SIEGEL D A,PETERSON A R.Optimization of a semianalytical ocean color model for global-scale applications[J].Appl Opt,2002,41(15):2705-2714.
[9] LAWS E A,BANNISTER T T.Nutrient-and light-limited growth ofThalassiosira fluviatilisin continuous culture,with implications for phytoplankton growth in the ocean[J].Limnol Oceanogr,1980,25(3):457-473.
[10] GEIDER R J.Light and temperature dependence of the carbon to chlorophyll ratio in microalgae and cyanobacteria:implications for physiology and growth of phytoplankton[J].New Phytol,1987,106(1):1-34.
[11] FALKOWSKI PG,LA ROCHE J.Acclimation to spectral irradiance in algae[J].JPhycol,1991,27(1):8-14.
[12] LEWIS M R,CULLEN J J,PLATT T.Relationships between vertical mixing and photoadaptation of phytoplankton:similarity criteria[J].Marine Ecology Progress Series,1984,15:141-149.
[13] CLAUSTRE H,MOREL A,BABIN M,et al.Variability in particle attenuation and chlorophyll fluorescence in the Tropical Pacific:Scales,patterns,and biogeochemical implications[J].JGeophys Res,1999,104(C2):3401-3422.
[14] BISHOP JK B,DAVISR E,SHERMAN J T.Robotic observation of dust storm enhancement of carbon biomass in the North Pacific[J].Science,2002,298(5594):817-821.
[15] STRAMSKI D,REYNOLDSR A,BABIN M,et al.Relationships between the surface concentration of particulate organic carbon and optical properties in the eastern South Pacific and eastern Atlantic Oceans[J]. Biogeosciences,2008,5:171-201.
[16] LOISEL H,BOSC E,STRAMSKI D,et al.Seasonal variability of the backscattering coefficients in the Mediterranean Sea based on Satellite SeaWIFS imagery[J].Geophys Res Lett,2001,28(22):4203-4206.
[17] VANTREPOTTE V,LOISEL H,MÉLIN F,et al.Global particulate matter pool temporal variability over the SeaWiFSperiod(1997-2007)[J]. Geophys Res Lett,2011,38(2):L02605.
[18] BEHRENFELD M J,BOSSE.The beam attenuation to chlorophyll ratio:an optical index of phytoplankton photoacclimation in the surface ocean?[J].Deep Sea Res Part I,2003,50(12):1537-1549.
[19] BEHRENFELD M J,BOSSE.Beam attenuation to chlorophyll concentration as alternative indices of phytoplankton biomass[J].JMar Res,2006,64(3):431-451.
[20] SIEGEL D A,BEHRENFELD M J,MARITORENA S,et al.Regional to global assessments of phytoplankton dynamics from the SeaWiFS mission[J].Remote Sensing of the Environment,2013,135:77-91.
[21] BEHRENFELD M J,MARANON E,SIEGEL D A,et al.A photoacclimation and nutrient based model of light-saturated photosynthesis for quantifying ocean primary production[J].Mar Ecol Prog Ser,2002,228:103-117.
[22] BEHRENFELD M J,BOSS E,SIEGEL D A,et al.Carbon based ocean productivity and phytoplankton physiology from space [J]. Global Biogeochem Cycles,2005,19(1):GB1006.
[23] CLAUSTRE H.IOCCG report No.11:Bio-Optical sensors on Argo floats[M].Dartmouth:International Ocean-Color Coordinating Group,2011.
[24] 邢小罡,赵冬至,CLAUSTRE H,等.一种新的海洋生物地球化学自主观测平台:Bio-Argo浮标[J].海洋环境科学,2012,31(5):733-739.XING Xiaogang,ZHAODongzhi,CLAUSTRE H,et al.A new autonomous observation platform of marine biogeochemistry:Bio-Argo floats[J].Marine Environmental Science,2012,31(5):733-739.
[25] 邢小罡,赵冬至,CLAUSTRE H,等.地中海黄色物质次表层极大值的季节变化观测[J].海洋学报,2013,35(2):35-42.XING Xiaogang,ZHAO Dongzhi,CLAUSTRE H,et al.Observations on seasonal variations of colored dissolved organic matter(CDOM)subsurface maximum in the Mediterrranean Sea[J].Acta Oceanologica Sinica,2013,35(2):35-42.
[26] 邢小罡,赵冬至,王林,等.南太平洋副热带环流区次表层叶绿素极大值的季节变化特征[J].高技术通讯,2013,23(10):1038-1047.XING Xiaogang,ZHAO Dongzhi,WANG Lin,et al.Seasonal variation characterisitics of subsurface chlorophyll maximum in the South Pacific subtropical gyre[J].Chinese High Technology Letters,2013,23(10):1038-1047.
[27] 邢小罡,邱国强,王海黎.Bio-Argo浮标观测北大西洋色素与颗粒物的季节分布[J].高技术通讯,2014,24(1):55-64.XINGXiaogang,QIU Guangqiang,WANGHaili.Seasonal distributions of pigment and particle in the North Atlantic observed by a Bio-Argo float[J].Chinese High Technology Letters,2014,24(1):55-64.
[28] XINGX,CLAUSTRE H,WANG H,et al.Seasonal dynamics in colored dissolved organic matter in the Mediterranean Sea:patterns and drivers[J].Deep-Sea Res Part I,2014,83:93-101.
[29] XING X,MOREL A,CLAUSTRE H,et al.Combined processing and mutual interpretation of radiometry and fluorimetry from autonomous profiling Bio-Argo floats:Chlorophyll a retrieval[J].J Geophys Res,2011,116(C6):C06020.
[30] DE BOYER MONTEGUT C,MADEC G,FISCHER A S,et al.Mixed layer depth over the global ocean:an examination of profile data and a profile-based climatology[J].J Geophys Res,2004,109(C12):C12003.
[31] MOREL A,HUOT Y,GENTILI B,et al.Examining the consistency of products derived from various ocean color sensors in open ocean(Case1)waters in the perspective of a multi-sensor approach[J].Remote Sens Environ,2007,111(1):69-88.
[32] LETELIER R M,KARL D M,ABBOTT M R,et al.Light driven seasonal patterns of chlorophyll and nitrate in the lower euphotic zone of the North Pacific Subtropical Gyre[J].Limnol Oceanogr,2004,49(2):508-519.
[33] MCLAUGHLIN F A,CARMACK E C.Deepening of the nutricline and chlorophyll maximum in the Canada Basin interior,2003-2009[J].Geophys Res Let,2010,37(24):L24602.
[34] MOREL A,CLAUSTRE H,GENTILI B.The most oligotrophic subtropical zones of the global ocean:similarities and differences in terms of chlorophyll and yellow substance[J].Biogeosciences Discuss,2010,7(4):5047-5079.
[35] BISHOP J K B.Transmissometer measurement of POC[J].Deep-Sea Res I,1999,46(2):353-369.
[36] STRAMSKI D,BOSSE,BOGUCKI D,et al.The role of seawater constituents in light backscattering in the ocean[J].Prog Oceanogr,2004,61(1):27-55.
[37] LOISEL H,BOSCE,STRAMSKID,et al.Seasonal variability of the backscattering coefficient in the Mediterranean Sea based on satellite SeaWiFS imagery[J].Geophys Res Lett,2001,28(22):4203-4206.
猜你喜欢
海洋信息技术与应用(2021年3期)2022-01-17
无损检测(2018年11期)2018-11-28
建筑热能通风空调(2018年5期)2018-07-09
现代测绘(2018年5期)2018-02-18
水利技术监督(2017年6期)2017-12-19
长江大学学报(自科版)(2017年19期)2017-10-14
中国测试(2016年3期)2016-10-17
海洋渔业(2016年6期)2016-04-16
中国工程咨询(2016年9期)2016-02-13
广东海洋大学学报(2015年3期)2015-12-22
