
丝栗栲、苦槠和青冈幼苗叶片功能性状对增温和施氮的响应1)
2014-03-06 03:20王致远赵广东
东北林业大学学报 2014年12期
王致远 赵广东 王 兵
(北京市通州区园林绿化局,北京,101101) (国家林业局森林生态环境重点实验室(中国林业科学研究院森林生态环境与保护研究所))
邓宗富 夏 晨 罗嘉东 王财英
(中国林业科学研究院亚热带林业实验中心) (江西省赣州市南康区林业技术推广站)
植物与环境的关系一直是植物生态学研究的重点,植物对环境的适应体现在植物的生活史、形态、物候等许多方面,何种植物适合何种环境,不同的植物有着不同的表现特征[1]。因此,许多植物生态学家致力于寻找一些重要的植物性状,以体现植物生长以及植物对环境的适应。而植物叶片是植物进化过程中对环境变化较敏感且可塑性较大的器官,其结构和功能特征最能体现环境因子的影响或植物对环境的适应[2]。叶片作为植物光合作用的主要场所,通常被作为表征植物资源利用、生物量和生态系统功能的性状,不仅能够解释其对环境的适应机理,而且能够预测生态系统对环境变化的重要影响[3]。
随着全球变暖的加剧,植物生长对温度的响应研究越来越受到关注。研究认为,增温一般能延长植物的生长季,将有利于净初级生产力的增加[4];增温导致群落地理格局向高寒区扩展[5],改变了植物地上和地下部分生物量的分配模式[6]。而在全球氮沉降不断增加的情况下,施氮可能会对植物的形态特征和生理特征产生显著影响,进而影响全球生态系统碳平衡和养分循环[7]。适量的氮有利于植物的生长,并且不同树种对氮沉降响应规律有所不同。李廷亮等对冬小麦(Triticum aestivum)的光合特性、产量和氮素利用的研究表明在0 ~270 kg·hm-2施氮量范围内,随施氮量的增加,旗叶的净光合速率和叶绿素含量增加,气孔导度增大,胞间二氧化碳浓度降低,旗叶蒸腾速率显著提高;但施氮量超过180 kg·hm-2时,除蒸腾速率外其它光合指标均无显著变化[8];Brown 等对3 种松树(Picea sitchensis,Thuja plicata and Tsuga heterophylla)研究发现当叶片氮质量分数小于21 mg·g-1时,所有树种的光合速率均随叶片氮质量分数增加而增加,但当叶片氮质量分数超过这一值时,光合速率反而下降[9];李德军等对3 种南亚热带树苗的研究表明氮处理没有对木荷(Schima superba)和黄果厚壳桂(Cryp tocarya concinna)幼苗的气孔导度产生明显影响,但显著促进了锥栗(C.chinensis)幼苗的气孔导度[10]。因此,研究不同环境对叶片性状功能的影响能够对深入理解植物对环境适应机理有重要意义。
本研究以中亚热带常绿阔叶林的主要优势树种丝栗栲(C.fargesii)、苦槠(C.sclerophylla)和青冈(C.glauca)幼苗为研究对象,通过MSR -2420 红外辐射增温装置和人工氮输入,设置出增温和不同施氮梯度,试图探讨其干质量面积、叶氮质量分数、叶氮面积质量及叶片干质量与饱和鲜质量比4 种功能性状指标对增温(W)、施氮(N1 和N2)及其交互作用(WN1、WN2)的响应规律,以期为未来全球变暖和氮沉降增加情景下中亚热带常绿阔叶林植物叶片功能性状的研究提供基础数据和科学依据。
1 研究区概况
研究地位于江西省分宜县大岗山国家级森林生态站,地理位置114°30' ~114°45'E,27°30' ~27°50'N,海拔约167.8 m,处于长江中下游低山丘陵区。该地区属亚热带湿润气候,雨量充沛、光照充足,非常适合林木生长。年平均气温15.8 ℃,年积温5 355 ℃,极端最高温39.9 ℃,极端最低温零下8.3 ℃,年平均日照时间1 656.9 h,年蒸发量504 mm,年降水量1 590.9 mm(其中4—6月份降雨量占全年降雨量的45%),年太阳辐射总量约487 kJ/cm2,年均相对湿度80%,无霜期265 d。土壤主要为红壤、黄壤类型,成土母质为砂页岩,土层厚度70~100 cm。地带性植被为天然常绿阔叶混交林,代表种是青冈栎(C.glauca)、丝栗栲(C.fargesii)、苦槠(C.sclerophylla)、刺栲(C.hystrix)、木荷(S.superba)、甜槠(C.eyrei)等。
2 材料与方法
2.1 材料
2010年秋季,在江西大岗山典型常绿阔叶林内人工采集丝栗栲、苦槠和青冈种子。丝栗栲母树、苦槠母树和青冈母树的生长条件相似,相距约为25 m。种子经过一个冬天的低温沙藏,于2011年春季在温室播种,在温室条件下生长一段时间后,将幼苗分别移栽至塑料钵中,每钵1 株,用砂土(m(土)∶m(沙)=3∶ 1)培养。3月初待幼苗长至5 cm 高左右,选择株高及生长状况基本一致的幼苗分别移至6 个大小一致的栽培小区内。每个栽培小区内,丝栗栲、苦槠、青冈幼苗随机栽植和分布。
2.2 栽培小区设置
江西大岗山国家级森林生态站站区选取周围无高大乔木或灌木遮挡的空旷地。2010年3月前修建2 个栽培池,其长、宽、深分别为4.30、3.80、1.20 m。再将每个栽培池等分成宽度均为1.0 m 的3 个栽培小区。每个栽培小区内的土壤均来自种子采集区,并尽量保持与种子采集区土壤层次相同顺序,以减少人为因素对土壤的扰动。
2.3 增温处理与环境监测
其中一个栽培池安装上MSR -2420 红外辐射增温装置,该装置悬挂在样地中央上方,通过可以散发红外线辐射的灯管实现模拟气候变暖。红外线辐射装置由美国伯利Kalglo 电子公司生产(Kglo Electronics,Bethlehem,PA,USA),型号MSR-2420,规格165 cm×15 cm。辐射器距地面垂直距离2.25 m,安放于3 m ×4 m 小区的中间部位,灯的最大功率是2 000 W,使用时调至8 档。从2011年3月份开始加热,全年365 d 不间断。每个对照区的中间部位,距地面垂直距离2.25 m 安装与红外线辐射器大小、形状相同的“假灯”以降低或消除辐射器遮阴或其它因素造成的实验误差。
2011年3月份在每个栽培池中间冠层中部处安装HC2 -S3 型空气温湿度传感器,同时在每个栽培小区5、20 cm 土壤处均安装EC-5 型土壤水分传感器和TR219 型土壤温度传感器以测量不同处理下不同深度土壤温湿度实时变化值。采样频率均为10 min 1 次。
2.4 施氮处理
2011年3月份对栽培池做施氮处理。在每个栽培池的3 个栽培小区内都设有同样的施氮梯度,依次为0、60、120 kg·hm-2·a-1。施氮处理条件下的3 个栽培小区依次记为CK、N1 和N2,增温和施氮共同处理条件下的3 个栽培小区依次记为W、WN1 和WN2。施肥频率为每2 个月1 次,每次将NH4NO3氮肥溶解于20 kg 的水里,用喷雾器均匀喷洒到栽培小区的土壤中,对照组喷洒等量的水。定期给幼苗松土,除草。
2.5 植物性状选择
本研究选取4 个关键性的叶片性状:①干质量面积(比叶面积),即叶片面积与其干质量之比,它是重要的结构性状之一,能够反映植物对不同环境的适应,使其成为比较生态学研究的首选指标[11]。有研究表明干质量面积与叶片的最大光合速率以及植物潜在的生长速率呈正相关关系,与叶片的寿命呈负相关关系,并且在环境条件越好的地方其值一般也越大[7]。②叶氮质量分数,以干质量计。植物体的氮素质量分数可以反映土壤氮素供应情况,常作为施肥指标[12]。③叶氮面积质量(单位面积叶氮含量),将叶氮质量分数通过干质量面积换算成叶氮面积质量,换算公式为:叶氮面积质量=叶氮质量分数/干质量面积[13]。④叶片干质量与饱和鲜质量比(干物质含量),为叶片干物质质量和叶片饱和鲜质量的比值,,反映了植物获取资源的能力[14]。
2.6 取样与分析
2012年10月份在6 个栽培小区里选样,从每个栽培小区里的每种树中上部选取10 片生长良好、没有病虫害的成熟叶片。取好后将其装入自封袋里,贴好标签。将样品拿回实验室,直接用LI -3000C 便携式叶面积仪(LI - COR Inc.,Lincoln,USA)测量每片叶片的叶面积,做好标记。然后将样品浸泡在水里,每隔5 h 测1 次叶片质量,直至质量不再增加为止,然后取出叶片,迅速用吸水纸吸走叶表面的水分,用电子天平称取其饱和质量。再将样品在65 ℃的烘箱里烘干至恒质量,测量其干质量。
2012年10月,在每个栽培小区内每种树选3株,其生长状况基本一致,每株树的中上部选取20片无病虫害,生长良好的成熟叶片,将叶片样品在105 ℃下杀青30 min,65 ℃的烘箱里烘干至恒质量,称取其干质量。然后将其磨碎过1 mm 筛备用,用凯氏定氮法测定叶片全氮质量分数。
2.7 数据分析
采用Excel 2010 软件进行数据的简单处理和图表绘制,并用SPSS17.0 软件进行方差分析以及增温施氮的交互作用分析,采用Duncan 方法检验不同处理间的差异。
3 结果与分析
3.1 红外辐射增温装置对空气、土壤温湿度的影响
3.1.1 空气温湿度
2012年1月—2013年1月,红外辐射增温装置明显提高了增温区的平均空气温度,增温平均幅度为1.22 ℃,并且增温区的空气温度变化与对照区基本一致,均在7—8月份达到最大值,1—2月份为最小值。由于空气温度的升高,空气相对湿度呈现出明显的下降趋势。与对照区相比,增温区的平均空气相对湿度下降了7.07%,并且在夏秋季节下降幅度大于春冬季节(图1)。

图1 红外辐射增温装置对试验区空气温湿度影响
3.1.2 土壤温湿度
2012年1月—2013年1月,红外增温辐射装置对土壤温度的影响和对空气温度的影响类似,增温区5、20 cm 处土壤平均温度均高于对照区,增幅分别为1.05、0.65 ℃,增温区和对照区温度变化基本一致,夏季达到最大值,冬季则值最小;并且无论是增温区还是对照区,20 cm 处土壤平均温度均大于5 cm 处土壤平均温度。与对照区相比,增温使得不同土层深度土壤平均体积含水量均有所下降,其中5 cm 处土壤平均体积含水量下降了7.02%,20 cm 处下降了5.52%;并且无论在增温区还是在对照区,5 cm 的土壤平均体积含水量均大于20 cm 处土壤平均体积含水量(图2)。
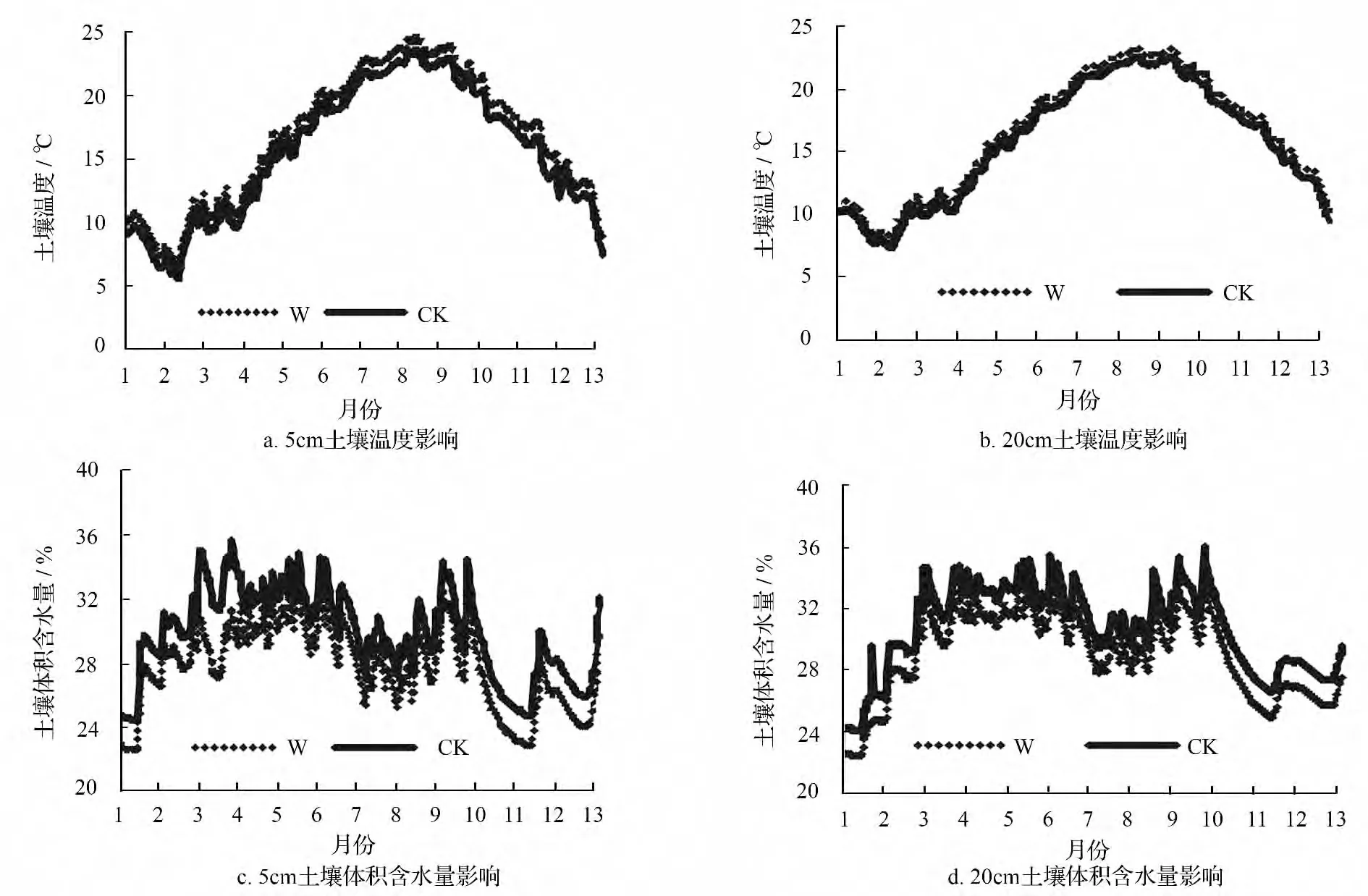
图2 红外辐射增温装置对试验区5、20 cm 深度土壤温度和体积含水量的影响
3.2 叶片干质量面积对增温和施氮的响应
对照条件下,3 种植物叶片的干质量面积表现为丝栗栲最大,苦槠次之。CK、W、N1、N2 和WN2 条件下,苦槠叶片的干质量面积均不同程度大于青冈。增温导致丝栗栲、苦槠和青冈叶片的干质量面积分别降低了4.6%、17.4%和10.8%。N1 处理时,丝栗栲、苦槠和青冈叶片干质量面积分别降低了0.3%、16.1%和12.3%。N2 处理时,丝栗栲、苦槠和青冈叶片干质量面积分别降低了0.5%、9.2%和10.0%。
总体来说,增温和施氮均引起3 种植物叶片干质量面积的降低,但降低程度随植物种类和施氮量的不同而不同。增温和施氮交互作用均导致3 种植物叶片的干质量面积减小,并且对苦槠和青冈叶片的交互作用明显(表1)。
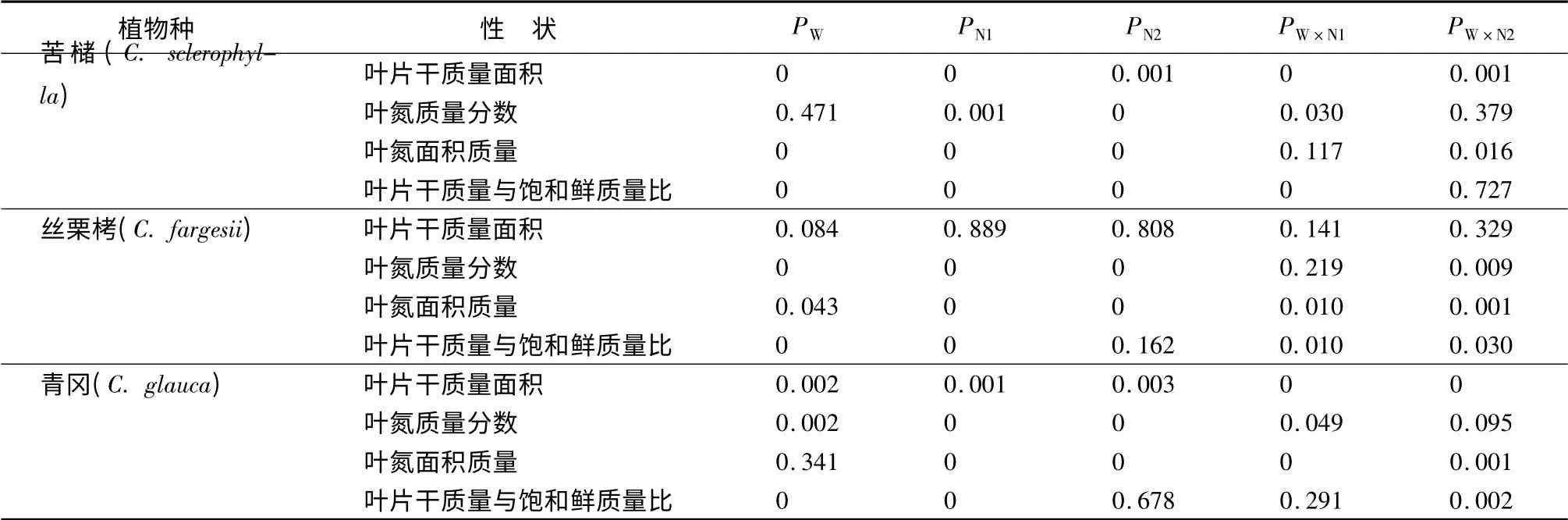
表1 不同处理的方差分析
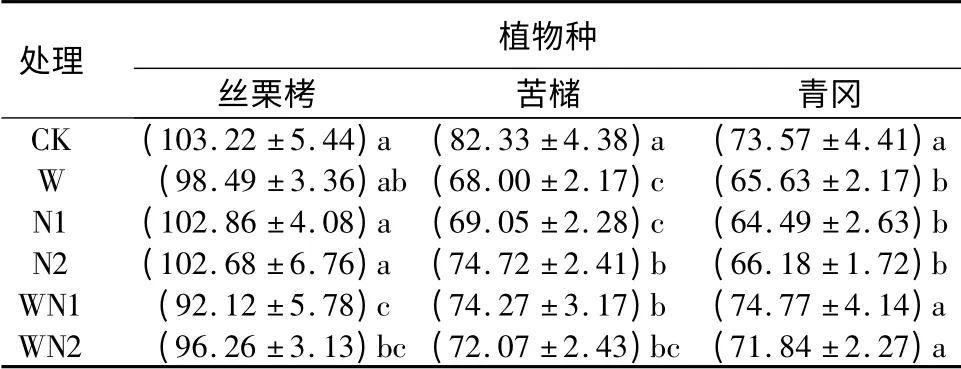
表2 不同处理对3 种植物幼苗叶片干质量面积的影响cm2·g -1
3.3 3 种植物叶片叶氮质量分数对增温和施氮的响应
对照条件下,丝栗栲、苦槠和青冈叶片叶氮质量分数分别为(13.45 ±0.48)、(12.61 ±0.81)、(13.25 ±0.33)mg·g-1。增温导致丝栗栲、苦槠和青冈叶片叶氮质量分数分别显著降低了13.4%、2.9%和8.9%。N1 处理时,丝栗栲、苦槠和青冈叶片干质量面积分别显著增加了48.3%、16.4%和14.8%。N2 处理时,丝栗栲、苦槠和青冈叶片干质量面积分别显著增加了47.1%、31.2%和16.9%。增温和施氮交互作用均引起丝栗栲、苦槠和青冈叶片叶氮质量分数的显著增加。WN1 条件下,丝栗栲、苦槠和青冈叶片叶氮质量分数分别增加了40.1%、28.3%和13.8%;WN2 条件下其增加值则分别为47.6%、22.7%和13.7%。
可见,增温对丝栗栲、苦槠和青冈叶片叶氮质量分数有明显的负效应,施氮则有明显的正效应。增温和施氮有明显交互效应,并且其正效应与施氮相一致,而且N1、N2、WN1 和WN2 对丝栗栲、苦槠和青冈叶片叶氮质量分数的影响程度差异不明显。
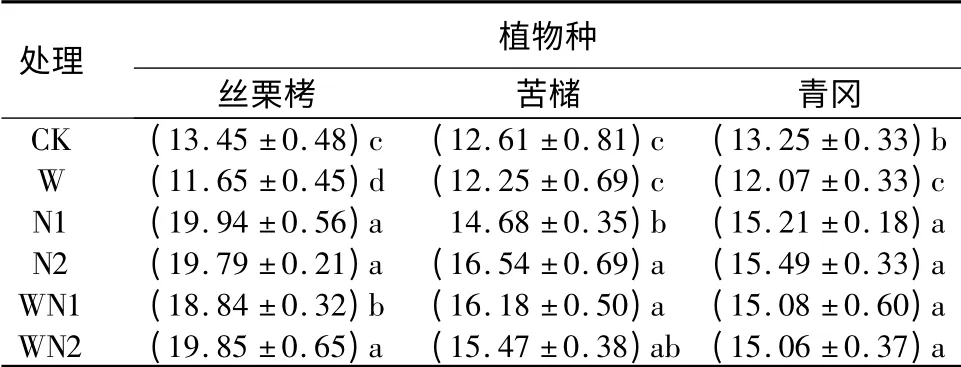
表3 不同处理对3 种植物幼苗叶片叶氮质量分数的影响mg·g -1
3.4 3 种植物叶片叶氮面积质量对增温和施氮的响应
增温条件下,丝栗栲叶片的叶氮面积质量明显降低,苦槠叶片明显增加,而青冈叶片则基本没有变化。单独施氮条件下,丝栗栲、苦槠和青冈叶片的叶氮面积质量均显著增加。N1 条件下时,3 种植物叶片的叶氮面积质量分别显著增加了46.2%、31.3%和33.3%;N2 条件下的对应值则分别为46.2%、37.5%和33.3%,可见施氮对3 个树种叶片的叶氮面积质量均有明显的正效应,并且N1 和N2 条件下差异不明显。除WN1 条件下的苦槠外,增温和施氮存在明显的交互作用(表1)。增温和施氮交互作用对丝栗栲叶片叶氮面积质量的增加效应大于单独施氮,苦槠表现为基本没有差异,而对青冈叶片叶氮面积质量的增加效应则比单独施氮小。总体而言,施氮有利于3 种植物幼苗叶片的叶氮面积质量增加,而其对增温、增温和施氮的交互作用响应模式则在不同植物间存在差异。
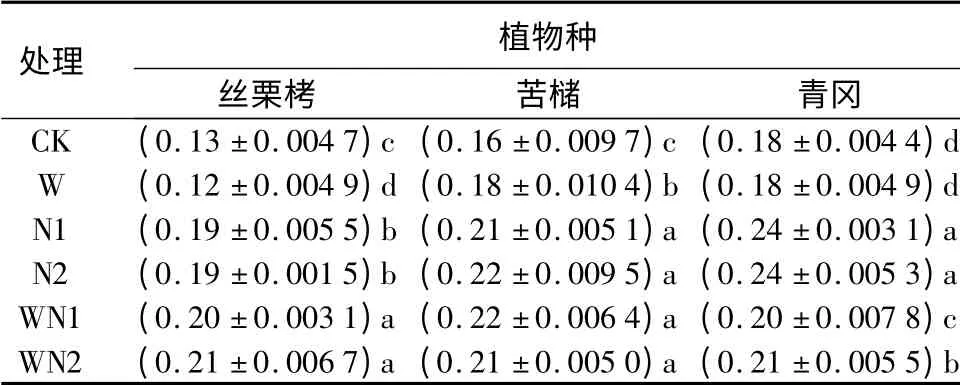
表4 不同处理对3 种植物幼苗叶片叶氮面积质量的影响mg·cm -2
3.5 3 种植物叶片干质量与饱和鲜质量比对增温和施氮的响应
对照条件下,3 种植物叶片干质量与饱和鲜质量比表现为青冈最大,苦槠次之。增温条件下,3 种植物叶片干质量与饱和鲜质量比分别显著增加了4.9%、4.4%和4.1%。N1 条件下时,3 种植物叶片干质量与饱和鲜质量比分别显著增加了4.9%、13.3%和2.0%;N2 条件下的对应值则分别为0、4.4%和0,可见N1 处理对叶片干质量与饱和鲜质量比的促进程度明显大于N2。对丝栗栲而言,增温、施氮以及增温和施氮交互作用对其叶片干质量与饱和鲜质量比的作用基本一致;苦槠表现为N1的作用最大,W 和N2、WN1 和WN2 的作用基本一致;青冈则为WN1 和WN2 作用明显大于增温、施氮。总体而言,增温、施氮以及增温和施氮对丝栗栲、苦槠和青冈叶片干质量与饱和鲜质量比均有明显的正效应,且增温和施氮间存在一定的交互作用(表1)。
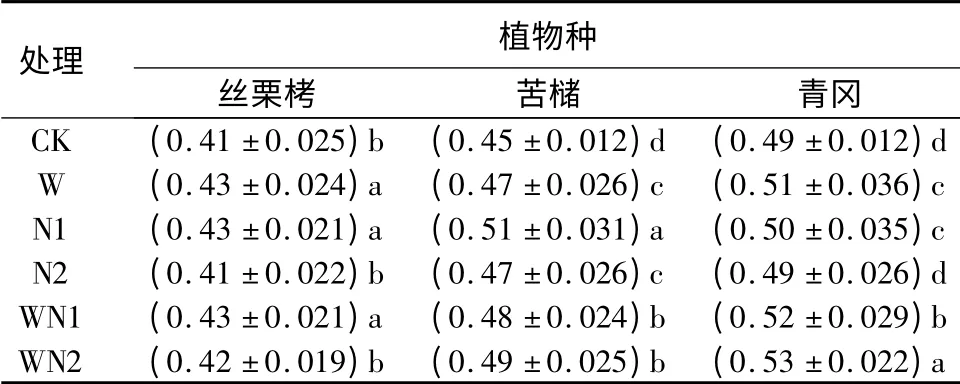
表5 不同处理对3 种植物幼苗叶片干质量与饱和鲜质量比的影响 g·g -1
4 讨论与结论
4.1 红外辐射增温装置增温效应
随着全球变暖的加剧,温度升高对植物生长乃至生态系统的影响备受关注,因此,许多研究人员致力于研究植物对模拟增温的响应。目前,国内外有许多用于科研的增温装置,都各有其优缺点和应用范围[15]。本研究采用的红外线辐射器主动加热方法,是野外使用最广泛的增温方式[16]。2012年1月—2013年1月期间,增温导致增温区平均空气温度比对照升高了1.22 ℃,平均空气相对湿度降低了7.07%。这与卫云燕等的研究中增温导致空气温度增加了1.62 ℃,空气相对湿度降低了9.94%相一致[17],其它也有类似的研究结果[18]。而与对照组比较,增温导致土壤温度增加幅度和土壤体积含水量减小幅度随土壤深度增加都有所减小,其中,5、20 cm 处土壤温度分别增加了1.05、0.65 ℃,5、20 cm处土壤体积含水量分别降低了7.02%、5.52%,这与羊留冬等对峨眉冷杉(Abies fabri)幼苗的研究结果一致[19]。这说明增温装置对表层土壤环境影响更为显著,可能是由于表层土壤与外界环境的物质和能量交换更为密切,更易受外界环境的干扰。
4.2 不同处理对不同树种性状的影响差异
可以看出,总体上温度的升高使得这3 种植物的干质量面积和叶氮质量分数都减小,叶片干质量与饱和鲜质量比明显增加,叶氮面积质量不同树种响应不同,苦槠明显增加,丝栗栲明显减小,而青冈基本没有变化。受温度升高的影响,干质量面积减小,这与羊留冬等对贡嘎山峨眉冷杉(A.fabri)幼苗生长的研究一致[19],但与其它研究的结论不一致[20-23],可能原因是温度升高叶面积增大,叶片的生物量也增大[6],但是其增长速率要比干质量面积的增长速率大。干质量面积的减小有利于植物能够更好地适应贫瘠的环境。已有研究表明温度升高会使得叶氮质量分数降低[24],可能原因是温度升高降低了叶片中的蛋白质和淀粉的含量,使得光合速率增加,所需的酶量也增加,从而使得氮向叶片中的分配增加[25],但是有些针叶树种的研究也有相反的结论[26-27],因此,这就说明叶氮质量分数对温度的响应存在种间差异。叶氮面积质量是由叶氮质量分数和干质量面积计算得到,本实验的研究结果表明增温使得叶氮质量分数和干质量面积都减小,叶氮面积质量的变化因树种而异。目前长期模拟增温对常绿阔叶幼苗的叶片干质量与饱和鲜质量比研究还比较少,一些研究结果表明温度的升高,阔叶乔木和落叶乔木的叶片干质量与饱和鲜质量比就会减小[28-29],而本研究结果则与其相反,这可能是由于温度增长1 ℃范围内时有利于这3 个树种的生长,其营养消耗相应增加,植物为了保证自身的生长,从而提高了资源利用效率以适应环境的变化。
氮素作为植物生长的重要组成元素,已经被人们熟知。近些年来,随着人为因素的干扰使得大气氮沉降过剩,远远超过了森林在生长季对氮素的正常需求(约5 ~8 kg·hm-2·a-1)[30]。本实验点位于江西省分宜县大岗山境内,随雨水输入到马尾松林和杉木林的氮分别为60.6、57.0 kg·hm-2·a-1[31],所施氮肥换算成氮沉降率,其值远远超过了大气氮沉降。在这种情况下,3 种植物幼苗的叶片性状有着不同的响应。以往的研究表明施一定量的氮肥导致叶片的干质量面积增加[7,32-33],而本研究结果是N1 和N2 情况下苦槠和青冈的干质量面积都减小,并且这两种植物受N1 的影响大于N2,而丝栗栲基本没有变化。干质量面积的减小可能是由于在富氮营养的条件下,植物幼苗生长受抑制而导致,同时也说明了苦槠和青冈受N1 和N2 的影响比丝栗栲更明显。叶氮质量分数、叶氮面积质量、叶片干质量与饱和鲜质量比对N1 和N2 的响应基本都是一致的,施氮都导致其明显增加,这与一些相关研究结果一致[34-36],并且叶片干质量与饱和鲜质量比受N1 的影响要大于N2,叶氮质量分数和叶氮面积质量在N1 和N2 的处理下,处理之间不同树种响应模式不同,但和对照相比都是增加。总的来说,施氮对这3 种植物的叶片氮质量分数和叶片干质量与饱和鲜质量比都有促进作用,进而也说明施氮有利于植物对养分的吸收和干物质的积累。
不同的植物叶片的性状对增温和施氮的响应模式不同,增温和施氮的交互作用在不同植物叶片性状上也有所不同(表1)。增温和施肥一般都是能够促进植物生长和生物量积累,温度升高导致土壤微生物比较活跃,而施氮导致肥土壤所需有效养分含量增加,进而影响土壤的一些基本特性[17]。因此,不同植物也就表现出不用的响应模式。本研究发现在增温和施氮交互作用下基本都使这3 种植物的干质量面积减小,并且对于苦槠和青冈有明显的交互作用。交互作用使得丝栗栲的干质量面积减小值比单个因子大,而青冈则相反,WN1 和WN2 之间没有明显差异;对叶氮质量分数的影响和单独施氮的影响基本一致;对叶氮面积质量的影响,就苦槠而言,和单独施氮基本一致,而对于丝栗栲,交互作用要大于单独施氮作用,WN2 大于WN1,而青冈则是交互作用大于单独增温作用,小于单独施氮作用,并且WN2 大于WN1;交互作用都使得叶片干质量与饱和鲜质量比增加,交互作用之间以及与单个因子之间又有不同的差异。可见,不同植物因研究区的气候条件、树种生物学特性、土壤性质等的不同而表现出不同的差异。当然,本研究由于受树种密度、增温的均匀性、施氮量、土壤性质等的影响,对于外推氮沉降和全球变暖的结果可能有一定的影响。因此,相关研究还需要进一步的深化。
[1] Garnier E,Laurent G,Bellmann A,et al.Consistency of species ranking based on functional leaf traits[J].New Phytologist,2001,152(1):69 -83.
[2] 董莉莉,刘世荣,史作民,等.中国南北样带栲属植物叶功能性状及其与环境因子的关系[J].林业科学研究,2009,22(4):463 -469.
[3] 韦兰英,上官周平.黄土高原不同退耕年限坡地植物干质量面积与养分含量的关系[J].生态学报,2008,28(6):2526 -2535.
[4] Suzuki S,Kudo G.Short-term effects of simulated environmental change on phenology,leaf traits,and shoot growth of alpine on a temperate mountain,northern Japan[J].Global Change Biology,1997,3(1):108 -115.
[5] Grabherr G,Gottfried M,Pauli H.Climate effects on mountain plants[J].Nature,1994,369:448 -450.
[6] 喻志强,赵广东,王兵,等.增温和施氮对丝栗栲和苦槠幼苗生长状况的影响[J].江西农业大学学报,2013,35(1):102 -107.
[7] 刘莹.施氮对油松幼苗生长特征的影响[D].西安:西北农林科技大学,2011.
[8] 李廷亮,谢英荷,洪坚平,等.施氮量对晋南旱地冬小麦光合特性、产量及氮素利用的影响[J].作物学报,2013,39(4):704 -711.
[9] Brown K R,Thompson W A,Weetman G F.Effects of N addition rates on the productivity of Picea sitchensis,Thuja plicata and Tsuga heterophylla seedlings[J].Trees,1996,10(3):189 -197.
[10] 李德军,莫江明,方运霆,等.模拟氮沉降对三种南亚热带树苗生长和光合作用的影响[J].生态学报,2004,24(5):876 -882.
[11] Wilson P J,Thompson K,Hodgson J G.Specific leaf area and leaf dry matter content as alternative predictors of plant strategies[J].New Phytologist,1999,143(1):155 -162.
[12] 黎莉,冯树丹,李蕊沁,等.三江湿地典型植被的叶片氮含量与比叶面积关系的研究[J].哈尔滨师范大学自然科学学报,2010,26(1):94 -97.
[13] 吕金枝,苗艳明,张慧芳,等.山西霍山不同功能型植物叶性特征的比较研究[J].武汉植物学研究,2010,28(4):460 -465.
[14] 万里强,李向林,石永红,等.PEG 胁迫下4 个黑麦草品种生理生化指标响应与比较研究[J].草叶学报,2010,19(1):83-88.
[15] 侯彦会,周广胜,许振柱.基于红外增温的草地生态系统响应全球变暖的研究进展[J].植物生态学报,2013,37(12):1153 -1167.
[16] Kimball B A.Theory and performance of an infrared heater for ecosystem warming[J].Global Change Biology,2005(11):2041-2056.
[17] 卫云燕,尹华军,刘庆,等.夜间增温和施肥对川西亚高山针叶林两种树苗根际效应的影响[J].生态学报,2011,31(3):698-708.
[18] Wan S,Luo Y,Wallace L L.Changes in microclimate induced by experimental warming and clipping in tallgrass prairie[J].Global Change Biology,2002(8):754 -768.
[19] 羊留冬,杨燕,王根绪,等.短期增温对贡嘎山峨眉冷杉幼苗生长及其CNP 化学计量学特征的影响[J].生态学报,2011,31(13):3668 -3676.
[20] 杨兵,王进闯,张远彬.长期模拟增温对岷江冷杉幼苗生长与生物量分配的影响[J].生态学报,2010,30(21):5994 -6000.
[21] 徐振锋,胡庭,张力,等.王开运.模拟增温对川西亚高山林线交错带绵穗柳生长、叶物候和叶性状的影响[J].应用生态学报,2009,20(1):7 -12.
[22] Loveys B R,Scheurwater I,Pon T L,et al.Growth temperature influences the underlying components of relative growth rate:an investigation using inherently fast-and slow-growing plant species[J].Plant,Cell and Environment,2002,25(8):975 -987.
[23] Tjoelker M G,Reich P B,Oleksyn J.Changes in leaf nitrogen and carbohydrates underlie temperature and CO2acclimation of dark respiration in fiveboreal tree species[J].Plant,Cell and Environment,1999,22(7):767 -778.
[24] Berry J,Björkman O.Photosynthetic response and adaptation to temperature in higher mountain plants[J].Annual Review of Plant Physiology and Plant Molecular Biology,1980,31:491 -543.
[25] Dewar R C,Medlyn B E,McMutrie R E.Acclimation of the respiration/photosynthesis ratio to temperature:in - sights from a model[J].Global Change Biology,1999,5(5):615 -622.
[26] Seppl K,Wang K Y.Effects of long-term CO2and temperature on crown nitrogen distribution and daily photosynthetic performance of Scots pine[J].Forest Ecology and Management,1997,99(3):309 -326.
[27] Zhao C Z,Liu Q.Growth and photosynthetic responses of two coniferous species to experimental warming and nitrogen fertilization[J].Canadian Journal of Forest Research,2009,39(1):1 -11.
[28] 张业.油松天然林林内主要植物种的叶功能性状研究[D].北京:北京林业大学,2012.
[29] 冯秋红,史作民,董莉莉,等.南北样带落叶乔木功能性状及其与气象因子的关系[J].中国农业气象.2009,30(1):79 -81.
[30] 郝吉明,谢邵东,段雷.酸沉降临界负荷及其应用[M].北京:清华大学出版社,2001.
[31] 魏样,同延安,段敏,等.陕北典型农区大气干湿氮沉降季节变化[J].应用生态学报,2010,21(1):255 -259.
[32] Jonannes M K,Kurt R.Specific leaf area along a nitrogen fertilization gradient[J].American midland naturalist,2000,144(2):265 -272.
[33] 黎蕾,蔡传涛,刘贵周.光强和施氮量对催吐萝芙木叶片生长及光合作用的影响[J].武汉植物学研究,2010,28(2):206 -212.
[34] Valera-Burgos J,Zunzunegui M,Díaz-Barradas M C.Do leaf traits and nitrogen supply affect decomposability rates of three Mediterranean species growing under different competition levels?[J].Pedobiologia,2013,56(3):113 -119.
[35] Guillard K,Allinson D W.Effects of Nitrogen Fertilization on a Chinese Cabbage Hybrid[J].Agronomy Journal,1988,80(1):21 -26.
[36] 郝敏.施肥对番茄功能特征与光合速率及果实产量关系的影响[D].兰州:兰州大学,2012.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
今日农业(2021年10期)2021-11-27
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
农业环境科学学报(2017年2期)2017-03-20
西藏科技(2016年9期)2016-09-26
江苏农业科学(2016年6期)2016-07-25
中国火炬(2015年12期)2015-07-31
农家顾问(2014年9期)2014-10-20
中国火炬(2013年3期)2013-07-24
