
平欧杂交榛13 个品种(系)抗寒性比较1)
2014-03-06 03:20韩俊威史彦江宋锋惠董玉芝哈迪尔依沙克
东北林业大学学报 2014年12期
韩俊威 史彦江 宋锋惠 董玉芝 哈迪尔·依沙克 韩 强
(新疆林业科学院经济林研究所,乌鲁木齐,830052) (新疆农业大学) (新疆林业科学院经济林研究所) (新疆农业大学)
20 世纪初期,我国主要种植平榛(C.heterophylla)和欧洲榛(C.avellana),且存在平榛产量低、口感差和欧洲榛抗逆性差等问题,远远不能满足市场需求,为有效解决此项问题,我国学者经过多年的科学研究,于20 世纪90年代末成功选育出杂交品种——平欧杂交榛(Corylus.heterophylla Fisch.×Corylus.avellana L.)。它是由我国原产的平榛(C.heterophylla)与引入的欧洲榛(C.avellana)种间远缘杂交选育出的优良品种(系),它们集中了欧洲榛与平榛的优良基因,即具有平榛的抗寒适应性强及优良风味的特点,又具有欧洲榛的大果、丰产、出仁率高的特点[2]。近几年平欧杂交榛已在东北、华北等区域形成一定的发展规模,是未来我国大果榛子栽培的首选品种,具有非常大的发展前景[3]。已鉴定的平欧杂交榛品种(系),适宜栽培范围在北纬32°~42°,即沈阳以南到长江以北地区栽培,冬季抗寒-35 ℃低温[2-4],但在北纬42°以北的地区栽培会出现冻害、抽条等现象[5],影响树体生长发育和产量。自2001年新疆林科院引进杂交榛,榛子种植面积稳中有升,产量持续增加,而北疆地区地处北纬43° ~49°,存在冬季寒冷干燥,灾害天气发生频繁等诸多问题,影响榛子产业的良性健康发展,但该区域具有光、热等资源优势,使得该区域发展榛子有很大潜力,因此,选择较为抗寒的品种对该区域的榛子发展具有重要意义。
低温是限制植物生长和分布的一种非生物胁迫因素[6],低温寒害是农林业生产中一种严重的自然灾害,据统计,世界每年因此造成的损失达2 000 亿美元[7],因此,研究植物抗寒性,不仅在理论上具有重要意义,而且在生产上也具有广泛的应用价值。关于植物抗寒性的研究有不少报道,通过测定相对电导率并结合logistic 方程推算出半致死温度的方法,是研究植物抗寒性最有效的手段之一,已应用于枣[8]、苹果[9]、果梅[10]、葡萄[11]、梨[12]等树种的抗寒能力鉴定上;同时,通过研究渗透调节物质游离脯氨酸[13]、丙二醛[14]和保护酶系统[15]等指标与植物抗寒性的关系来鉴定其差异性的方法,也都已广泛应用;也有研究者认为通过观察枝条恢复发芽能力的方法,可以客观反映植物的实际抗寒水平[16]。目前对不同树种的抗寒性研究很多,但对平欧杂交榛在新疆的抗寒适应性方面的研究报道却较少。
本试验以13 个平欧杂交榛品种(系)为研究对象,通过对抗寒生理指标的测定、分析和比较,研究各品种(系)间的差异性,综合评价其抗寒性,筛选出抗寒性较强的品种(系),为杂交榛在伊犁河谷、天山北坡等不同生态区域的生产推广提供理论依据。
1 材料与方法
供试的13 个平欧杂交榛品种(系)于2001年从辽宁林科院引进,种植于新疆伊犁州霍城县大西沟乡榛子资源汇集圃。于2013年1月上旬采集枝条,每个品系随机选取5 株树,选取植株中上部、充分成熟、无病虫害、粗度接近、长度为30 ~40 cm 的1年生枝条作为试材,所选枝条的朝向和部位尽量一致,用凡士林涂抹剪口,按品系贴上标签装入聚乙烯袋中,带回实验室,在冰箱中0 ~4 ℃储存。13 个平欧杂交榛品种(系)的原编号分别为:80-13、85-152、82-11、84-36、82-15、80-4、85-49、80-43、84-254、B21、84-310、84-226 和83-33。
1.1 试验处理
将采回的1年生枝条剪成30 cm 的长度,先用自来水冲洗,除去表面污垢,再用去离子水冲洗3遍,用吸水纸吸干水分。每个品系分成6 份,用干净纱布包裹放入塑料袋中,置于可控超低温冰箱中进行人工低温处理。共设置-15、-20、-25、-30、-35、-40 ℃6 个梯度,以-15 ℃为对照,冷冻时降温幅度和解冻时的升温幅度均为4 ℃/h,到达设定温度后维持12 h,然后解冻。进行有关生理指标测定时,每处理均重复3 次。
1.2 试验方法
相对电导率测定:将低温处理过的枝条剪成2~3 cm 长的小段,避开芽眼,称1 g 放入三角瓶中,加入20 mL 去离子水,在室温下浸泡12 h,用DDS-307 型电导率仪测定浸提液的初电导率C1,然后将三角瓶置于沸水中煮沸15 min,然后将浸提液定容至20 mL,待浸提液冷却至室温时,摇匀,测定终电导率C2,计算相对电导率。相对电导率=(C1/C2)×100%。
半致死温度的测定:相对电导率拟合Logistic 方程为y=k/(1 +ae-bx),其中y 为实测相对电导率,x为处理温度,k 为细胞伤害率的饱和容量,a、b 为方程常数。在实际应用中,常令y' =ln[(k-y)/y],y'=ln(a-bx),即电解质外渗率y 转换成y'后,与处理温度x 之间的关系可以用线性方程表示,故可以按一般的直线相关法求出a 和b。在数学上,拐点即dy2/dx2=0 时的x 值,可获得曲线的拐点温度x=ln(a/b),即为半致死温度值,并求出方程的拟合度[17]。
恢复生长试验:将低温处理过的枝条,在温室(20 ~25 ℃)的花盆内进行扦插,以V(蛭石)∶ V(珍珠岩)=1∶ 1 为基质,观察发芽情况,1 个月后统计萌芽率。
游离脯氨酸质量分数测定采用酸性茚三酮法。丙二醛质量摩尔分数测定采用硫代巴比妥酸法。过氧化物酶活性测定采用愈创木酚法。
1.3 数据处理
用Excel2007 进行数据处理,SPSS19.0 软件进行方差分析等。运用隶属函数值法进行综合评价[18],计算公式:Uij= (Xij- Xjmin)/(Xjmax- Xjmin)。式中:Uij表示i 种类j 指标的抗寒隶属函数值;Xij表示i 种类j 指标的测定值;Xjmin表示所有种类j 指标的最小值;Xjmax表示所有种类j 指标的最大值;i 表示某个品种;j 表示某项指标。
2 结果与分析
2.1 不同低温处理后杂交榛相对电导率的变化
从图1中看出,在经过一系列低温处理后,杂交榛的相对电导率与处理温度间呈“S”型曲线,各个品种(系)的相对电导率在处理温度高于-25 ℃时,呈缓慢上升趋势;在-25 ~-35 ℃,表现为急剧上升状态;当处理温度低于-35 ℃时,上升缓慢且平缓。相对电导率急剧增大,说明枝条组织已经受到严重伤害,膜透性增加,细胞内电解质大量外渗。13个品种(系)在相同低温处理条件下的相对电导率差异较大,而且随着处理温度的降低,相对电导率的上升速度并不均匀,增加幅度在85.6% ~141.2%,这可能与品种(系)间的抗寒差异性有关。相对电导率与处理温度间的相关系数为-0.965 ~-0.988,呈极显著负相关(P <0.01),这与前人在其它树种上的研究结果基本一致[8]。
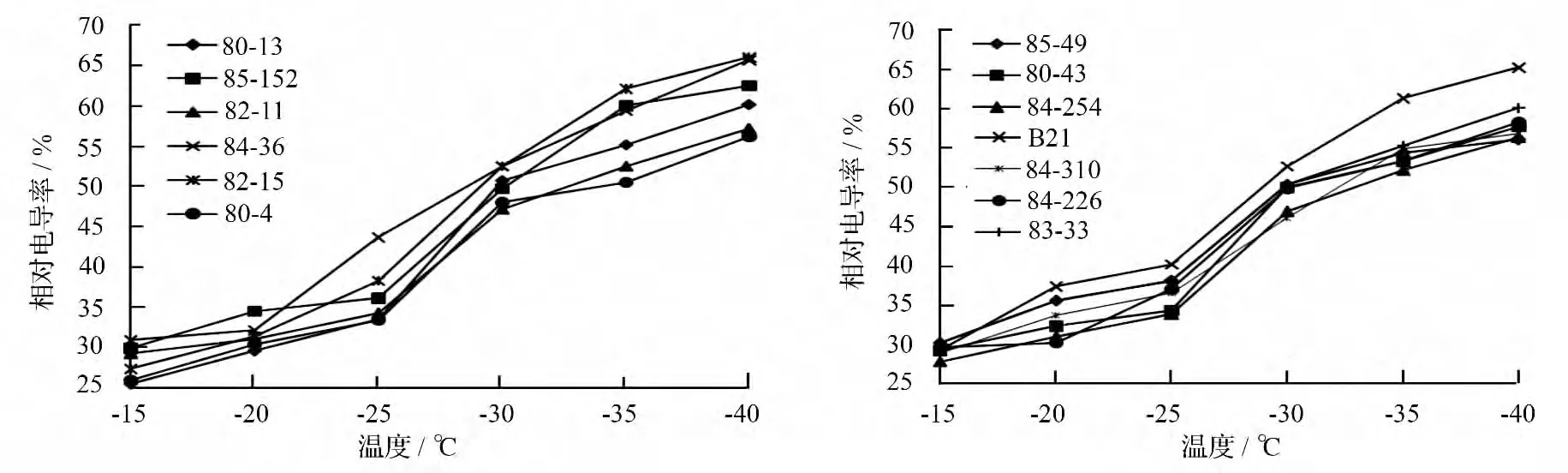
图1 低温处理后杂交榛枝条相对电导率的变化
2.2 半致死温度的比较
以相对电导率拟合Logistic 方程求得的半致死温度可反映植物在低温下保持膜系统稳定性和完整性的能力,进而反映植物的抗寒能力[17]。根据材料在不同低温处理下的相对电导率求得的Logistic 方程、拟合度及半致死温度(见表1),13 个平欧杂交榛品种(系)的半致死温度为-27.21 ~-33.99 ℃;拟合度都大于0.900,均达极显著水平(P <0.01);抗寒性不同的品种(系)间的半致死温度存在差异,半致死温度与抗寒力间呈负相关趋势,半致死温度越低,抗寒力则越强,反之则越弱。抗寒性较强的是84-254、84-310,半致死温度低于-32 ℃;其次是85-152、82-11、80-43 和83-33,半致死温度低于-30 ℃;最后是84-36、82-15、85-49、84-226、80-13、80-4 和B21,半致死温度低于-27 ℃。
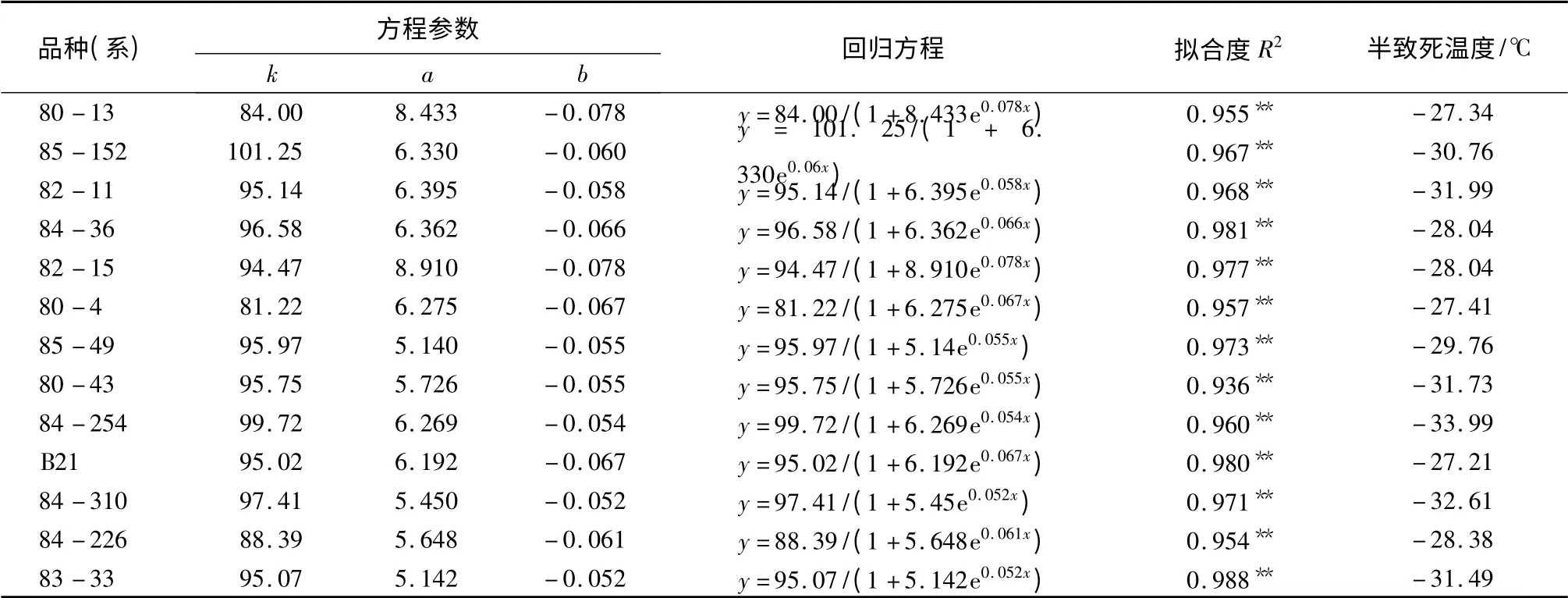
表1 平欧杂交榛枝条相对电导率Logistic 方程参数及半致死温度
2.3 不同低温处理后丙二醛质量摩尔浓度的变化
如表2所示,13 个杂交榛品种(系)的丙二醛(MDA)质量摩尔浓度随着温度的降低,其变化趋势整体呈增长状态,在-15 ℃时,84-254 的MDA 质量摩尔浓度最小,为2.69 μmol/g;85-49 的MDA质量摩尔浓度最大,为6.12 μmol/g。在-20 ~-40℃时,各品种(系)较对照(-15 ℃)均出现不同程度的升高。MDA 是由于植物受冻害后细胞膜透性遭到破坏,进而引发膜脂过氧化作用而产生的物质,在受冻害时,MDA 质量摩尔浓度持续增大,但因品种差异,各自达到高峰时的温度不同[19],达到峰值时的温度越低,则抗寒性较强;反之,则较弱。经方差分析可知,平欧杂交榛的MDA 质量摩尔浓度在不同品种(系)间和温度间差异均达极显著水平(P<0.01),可见低温胁迫和品种差异是影响杂交榛MDA 质量摩尔浓度的两大因素。本研究中83-33和84-254 的MDA 质量摩尔浓度达到最大值时所对应的温度最低,为-40 ℃,比其余品种(系)的耐寒性强;85-152、82-11、80-43、84-36、B21 和84-310 都是在温度降到-35 ℃时MDA 质量摩尔浓度达到最高;其余5 个品种(系)是在温度降到-30℃时MDA 质量摩尔浓度达到最高,可见耐寒性最弱。
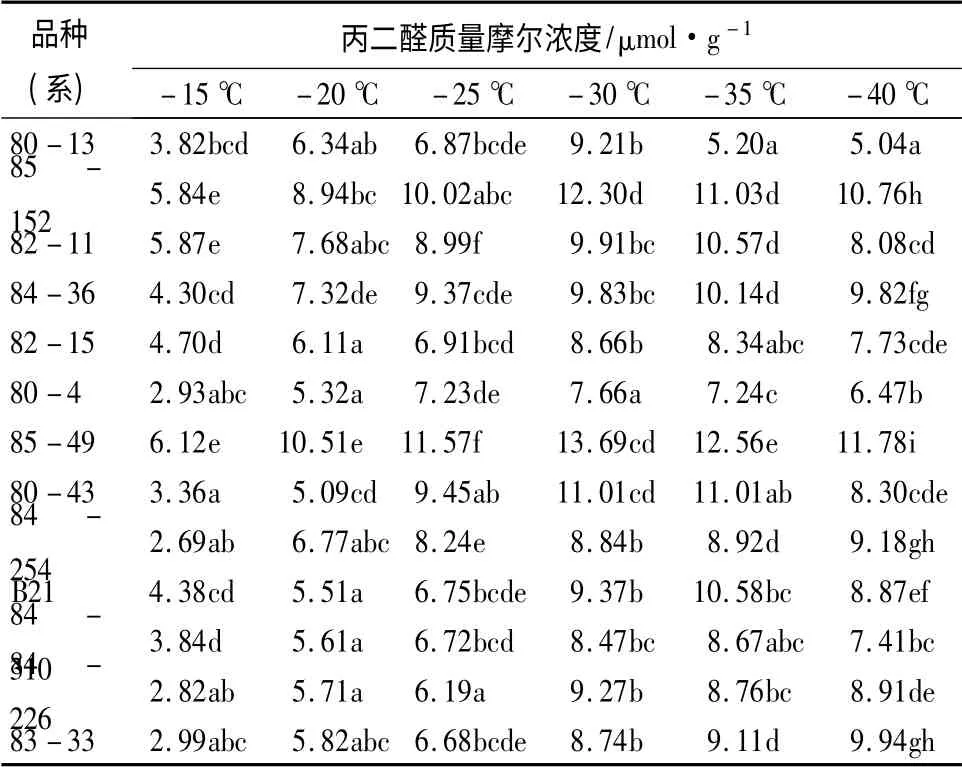
表2 不同低温处理后杂交榛枝条丙二醛质量摩尔浓度的变化
2.4 不同低温处理后游离脯氨酸质量分数的变化
如表3所示,13 个杂交榛品种(系)的游离脯氨酸质量分数随着温度的降低,其变化趋势基本呈逐渐增长状态,在-15 ℃时,85-152 和84-254 的游离脯氨酸质量分数最大,为38.86 μg/g 和38.09 μg/g;80-4 的游离脯氨酸质量分数最小,为27.49 μg/g。在-20 ~-40 ℃,各品种(系)较对照(-15℃)均出现不同程度的升高。游离脯氨酸是植物逆境下的产物,一般认为,低温胁迫往往伴随着游离脯氨酸质量分数的增加,游离脯氨酸质量分数越高,表明抗寒性越强;反之,则抗寒性越弱[20]。经方差分析可知,平欧杂交榛的游离脯氨酸质量分数在不同品种(系)间和温度间差异均达极显著水平(P <0.01)。由表3可知,在不同处理低温时,85-152、82-11、84-36、85-49 和84-254 的游离脯氨酸质量分数均相对较高,则抗寒性相对较强;80-13、80-4和84-226 的游离脯氨酸质量分数均相对最低,则抗寒性相对较弱;其余5 个品种(系)的游离脯氨酸质量分数较为接近,介于上述两者之间,则抗寒性居中。
2.5 不同低温处理后过氧化物酶活性的变化
如表4所示,13 个杂交榛品种(系)的过氧化物酶(POD)活性随着温度的降低呈下降趋势,在-15℃,84-254 的POD 活性最大,为657.79 μg/g;84-36 的POD 活性最小,为172.50 μg/g。在-20 ~-40 ℃,各品种(系)较对照(-15 ℃)均出现不同程度的降低。植株可通过提高POD 活性以减少低温胁迫产生的超氧化物自由基的伤害,保证植物细胞的正常机能,可以通过POD 活性的高低来判断植物的抗寒性[21],一般认为,POD 活性高,则抗寒性较强;反之,抗寒性较弱。经方差分析可知,平欧杂交榛的POD 活性在不同品种(系)间和温度间差异均达极显著水平(P <0.01),可知在不同处理低温时,POD 活性均相对较大的是85-152、80-43、84-254和84-310,则其抗寒性较强;82-11、84-36、80-4和83-33 的POD 活性在不同低温内均相对较低,则抗寒性较弱;其余5 个品种(系)的POD 活性介于上述两者之间,则抗寒性居中。
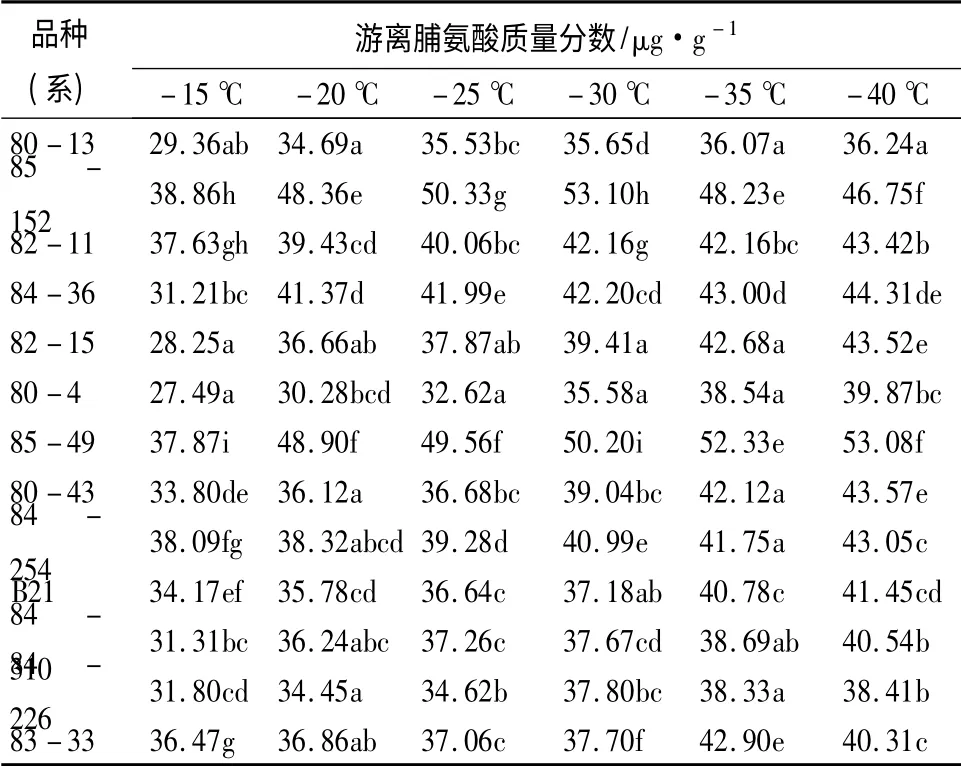
表3 不同低温处理后杂交榛枝条游离脯氨酸质量分数的变化
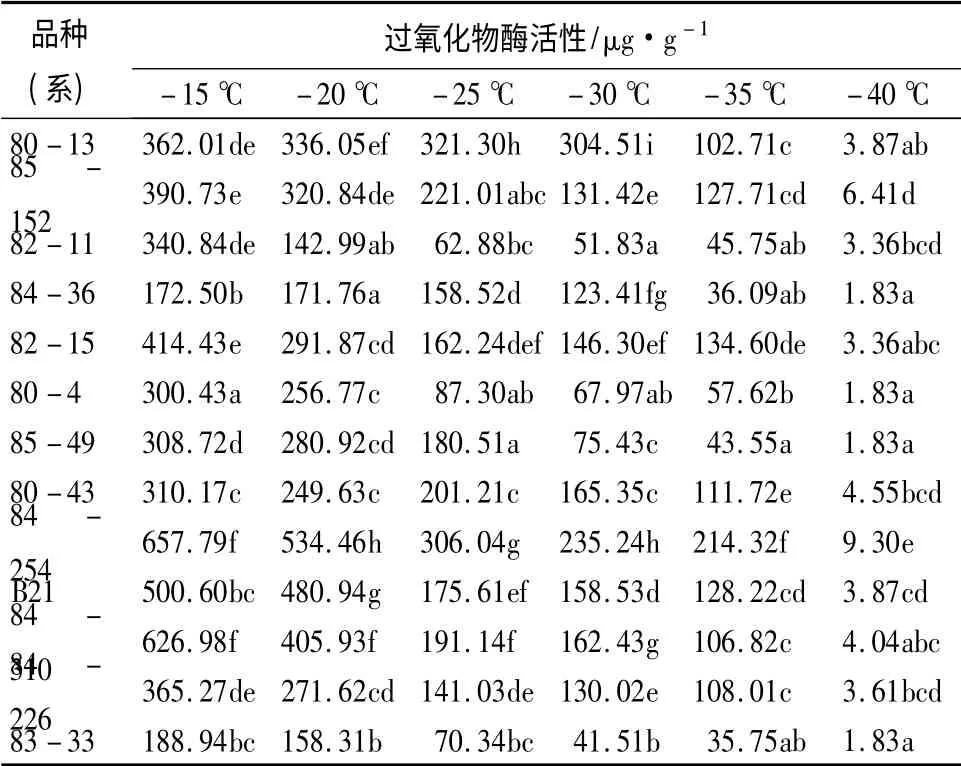
表4 不同低温处理后杂交榛枝条过氧化物酶活性的变化
2.6 不同低温处理后萌芽率的变化
萌芽率是评价树木受冻害程度或能否存活的最直观方法,能够客观反映品种的实际抗寒水平。萌芽率高,则自身抵御冻害能力较强;反之,则较弱。如表5所示,13 个杂交榛品种(系)的萌芽率随着温度的降低呈下降趋势,在-15 ~-20 ℃的萌芽率几乎达100%,差异不显著。在-25 ~-40 ℃低温诱导后各品种(系)间萌芽率差异显著,84-254 的萌芽率均相对较高,则抗寒性较强;80-13 的萌芽率均相对较低,其抗寒性较弱;其余11 个品种(系)的萌芽率相对集中,其抵御冻害能力较为接近,抗寒性居中。
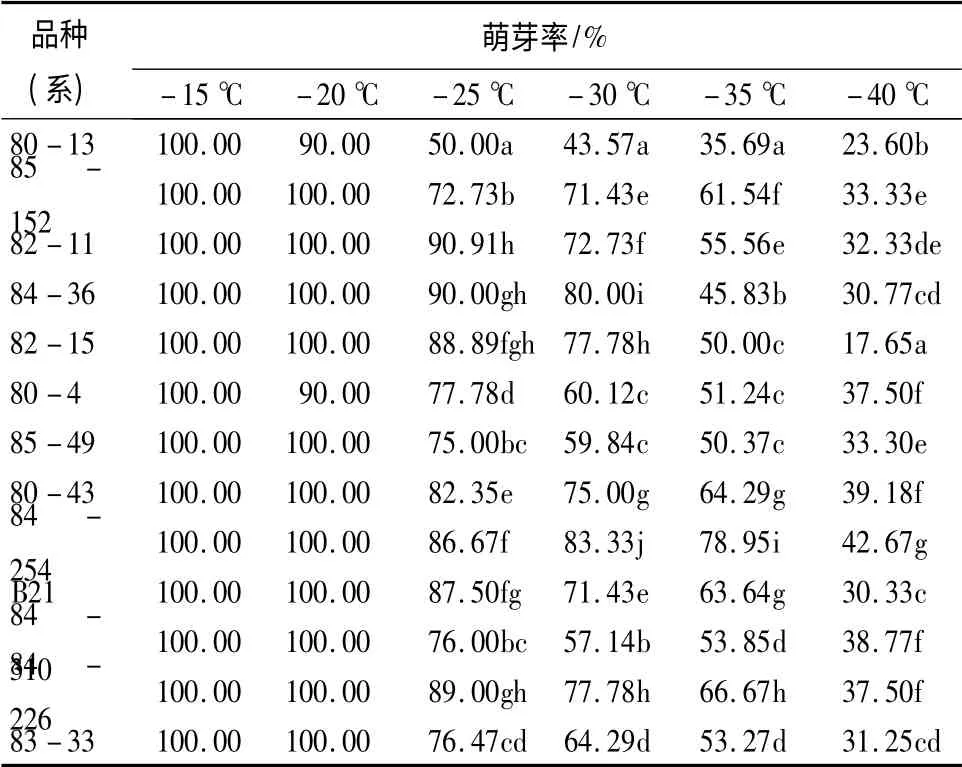
表5 不同低温处理后杂交榛枝条萌芽率的变化
2.7 相关性分析
如表6所示,对13 个杂交榛品种(系)的丙二醛质量摩尔浓度、游离脯氨酸质量分数、过氧化物酶、萌芽率与相对电导率进行相关性分析。结果表明,丙二醛质量摩尔浓度、游离脯氨酸质量分数与对电导率呈极显著正相关(P <0.01),相关系数达0.655 和0.517;过氧化物酶、萌芽率与相对电导率呈极显著负相关(P <0.01),相关系数达-0.759和-0.889。测定相对电导率是研究植物抗寒性最有效的手段之一,本研究中的4 个抗寒指标与相对电导率的相关性均达极显著相关(P <0.01),表明除相对电导率和半致死温度作为传统的比较可靠指标外,丙二醛质量摩尔浓度、游离脯氨酸质量分数、过氧化物酶和萌芽率也可作为鉴定平欧杂交榛抗寒性的相关指标。
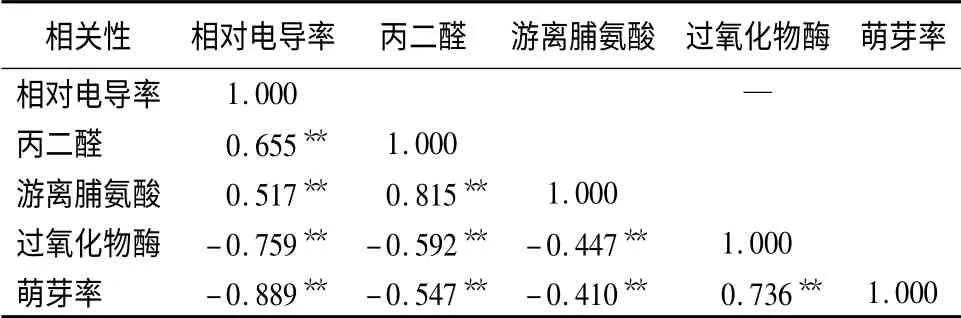
表6 4 个指标与相对电导率的相关性分析
2.8 平欧杂交榛抗寒性综合评价
由以上分析可知,丙二醛质量摩尔浓度等4 个指标在不同品种(系)间差异极显著,且与相对电导率呈极显著相关,可以有效评价平欧杂交榛抗寒性。以相对电导率、丙二醛质量摩尔浓度、游离脯氨酸质量分数、过氧化物酶活性和萌芽率5 个指标为依据,计算各指标的隶属函数值,并进行综合评价,结果见表7。抗寒性由大到小排列顺序为:84-254、85-152、84-310、84-226、80-43、B21、85-49、82-15、80-13、82-11、80-4、84-36、83-33。
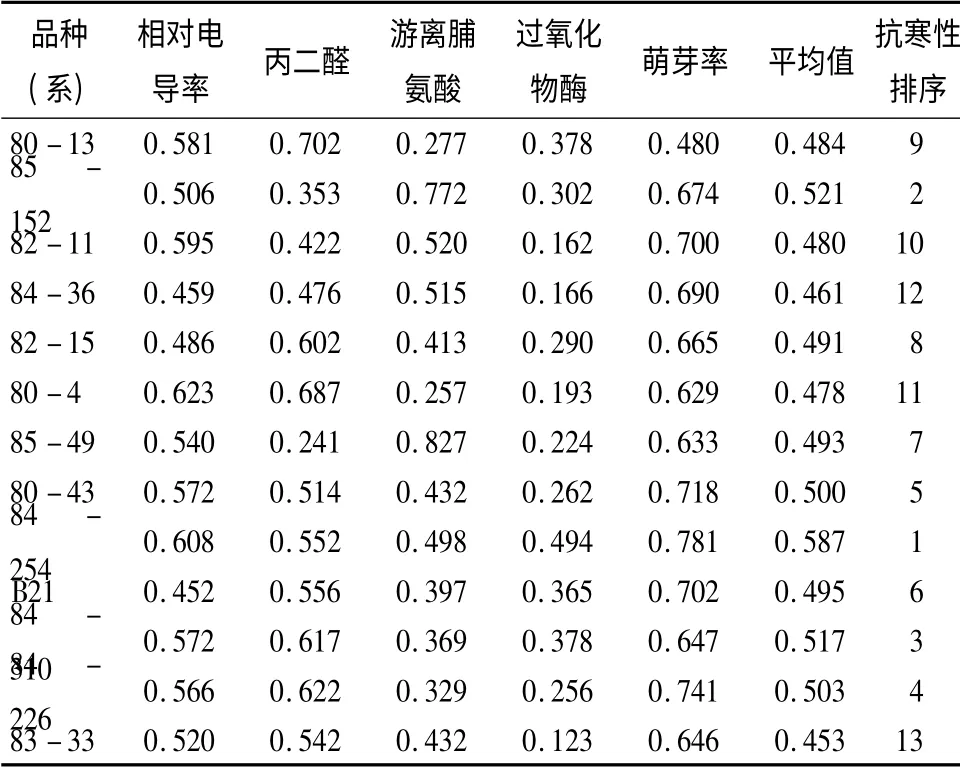
表7 低温胁迫条件下平欧杂交榛隶属函数值
3 结论与讨论
植物受到低温侵害时,细胞的质膜透性发生改变,膜脂过氧化程度加剧,活性氧的代谢平衡受到破坏,活性氧的增加对植物起到伤害作用。同时,植物体内的渗透调节物质和保护酶系统保护植物不受或受到较小伤害[22]。植物的生理过程是错综复杂的,受多种因素影响,孤立地用某一指标表示这一复杂的抗寒生理过程,不能真实反映植物的抗寒水平[23]。本文通过研究筛选出有效抗寒指标,利用隶属函数分析对材料进行综合评价,此法不仅给我们提供了一条在多指标测定基础上对材料特性进行综合评价的途径,而且能克服只利用少数指标进行评价的不足,以揭示不同品种间对低温反应特性的实质,从而提高抗寒性鉴定的准确性。这一方法已运用在杨树[18]抗寒性的研究中,因此,此法应用于平欧杂交榛抗寒品种筛选更具科学性和可靠性。
Lyons[24]认为,植物受到低温影响时,细胞的质膜透性会明显增加,电解质发生不同程度外渗,导致电导率不同程度的加大,可作为抗寒性的生理指标。测定植物电解质渗出率是不同树种常用的鉴定抗寒力水平的方法[8-10],本文通过研究电解质渗出率与丙二醛(MDA)、游离脯氨酸(Pro)、过氧化物酶(POD)、萌芽率的相关性,确定了除电解质渗出率以外的4 个指标也可作为鉴定平欧杂交榛的有效指标,多个指标综合分析其抗寒性,可以增加结果的精确性。
Peter[25]提出,将电解质渗出率达到50%时的温度视为半致死温度。但后续的研究结果[10]表明,利用电导法测定一系列温度下植物组织的电导率并配以Logistic 方程回归分析,以拐点温度表示植物组织的半致死温度更为准确。本研究通过测定发现84-254、84-310、85-152、82-11、80-43 和80-33 的半致死温度低于-30 ℃,其余7 个品种(系)的半致死温度则高于-30 ℃,说明前者的抗寒力强于后者,适应低温能力较强,能够在不同低温区域适当生长。半致死温度越低,说明抗低温能力较强;反之,则较弱。这与葡萄[11]、洋梨[12]抗寒性的研究结果相一致。
本研究结果表明,13 个平欧杂交榛品种(系)的MDA 质量摩尔浓度变化规律整体呈增长趋势,各自达到波峰时的温度不同,这可能由于自身遗传抗寒特性不同引起的,与玉米中MDA 作为衰老指标[19]中的研究观点一致;游离脯氨酸质量分数的变化规律不完全一致,这可能是低温胁迫后枝条的游离脯氨酸参与了蛋白质的合成或者被氧化的原因,大多数学者对此研究不尽相同[20],有待进一步研究;POD 活性的变化规律相同,随温度降低呈下降趋势,这与前人[21]的研究结果一致。萌芽率的变化趋势与半致死温度的变化相接近,表明二者可以有效结合,分析不同品种间抗寒性差异。结合本文研究可知,13 个平欧杂交榛品种(系)对以上5 项指标的抗寒性反应不同,且用某一项指标单一评价杂交榛抗寒性时的结果存在差异,不能统一分析品种(系)间的抗寒差异性。因此,运用隶属函数法综合有效指标进行全面系统评价。
用相对电导率、MDA、游离脯氨酸、POD 和萌芽率等指标,运用隶属函数值法综合分析平欧杂交榛抗寒性,可知其抗寒性排序为:84-254、85-152、84-310、84-226、80-43、B21、85-49、82-15、80-13、82-11、80-4、84-36、83-33。84-254、85-152 和84-310 这3 个品种(系)在低温下仍有一定生存力,可以适应复杂低温环境;80-4、84-36 和83-33 能适应的低温环境有限。新疆北疆地区冬季气温普遍较低,每年寒潮侵入的时间、强度及持续时间,都是造成不同程度冻害的原因,严重影响新疆榛子产业的持续健康高效发展。因此,鉴定不同品种间抗寒性,有选择的栽种抗寒性强的品种(系),可以有效解决低温侵害带来的损失。
[1] 张宇和.中国果树志:板栗榛子卷[M].北京:中国林业出版社,2005.
[2] 梁维坚.大果榛子育种与栽培[M].北京:中国林业出版社,2002:1-2.
[3] 侯志霞,原牡丹,刘雪梅,等.我国榛子生产研究概况[J].经济林研究,2008,26(2):123-126.
[4] 叶国盛.高抗寒杂交大果榛子[J].特种经济动植物,2006(1):29.
[5] 马庆华,王贵禧,梁维坚,等.我国榛属植物种质资源的研究、利用与创新[J].果树学报,2013,30(1):159-164.
[6] 彭筱娜,易自力,蒋建雄.植物抗寒性研究进展[J].生物技术通报,2007(4):15-18.
[7] 卢存福.第七界国际植物抗寒会议概况[J].植物学通报,2004,21(5):617.
[8] 王长柱,高京草,李新岗,等.西北地区枣树主栽品种抗寒性研究[J].果树学报,2011,28(5):898-902.
[9] 时朝,王亚芝,刘国杰.应用Logistic 方程确定五种苹果枝条的半致死温度的研究[J].北方园艺,2013(2):36-38.
[10] 高志红,章镇,韩振海.果梅种质枝条抗寒性鉴定[J].果树学报,2005,22(6):709-711.
[11] 张倩,刘崇怀,郭大龙,等.5 个葡萄种群的低温半致死温度与其抗寒适应性的关系[J].西北农林科技大学学报:自然科学版,2013,41(5):149-154.
[12] 李俊才,刘成,王家珍,等.洋梨枝条的低温半致死温度[J].果树学报,2007,24(4):529-532.
[13] 赵一鹤,李建宾,杨时宇,等.低温胁迫下3 个甜角品种幼苗的抗寒性研究[J].林业科学研究,2012,25(2):150-156.
[14] 冯建灿,张玉洁,杨天柱.低温胁迫对喜树幼苗SOD 活性、MDA 和脯氨酸含量的影响[J].林业科学研究,2002,15(2):197-202.
[15] Bowler C,Van Montagn C,Inze D.Superoxide dismutase and stress tolerance[J].Ann Rev Plant Physiol Plant Mol Biol,1992,43:83-116.
[16] 郑元,杨途熙,魏安智,等.低温胁迫对仁用杏几个抗寒生理指标的影响[J].西北农林科技大学学报:自然科学版,2008,36(1):164-167.
[17] 朱根海,刘祖祺,朱培仁.应用Logistic 方程确定植物组织低温半致死温度的研究[J].南京农业大学学报,1986,9(3):11-16.
[18] 史清华,高建社,王军,等.5 个杨树无性系抗寒性的测定与评价[J].西北植物学报,2003,23(11):1937-1941.
[19] 陈贵,胡文玉,谢浦缔,等.提取植物体内MDA 的溶剂及MDA作为衰老指标的探讨[J].植物生理学通讯,1991,27(1):44-46.
[20] 汤章诚.逆境条件下植物脯氨酸的积累及其可能的意义[J].植物生理学通讯,1984,20(1):15-17.
[21] 聂庆娟,孟朝,梁海永,等.低温胁迫对4 种常绿阔叶植物膜脂过氧化及保护酶活性的影响[J].植物研究,2007,27(5):578-581.
[22] 和红云,田丽萍,薛琳.植物抗寒性生理生化研究进展[J].天津农业科学,2007,13(2):10-13.
[23] 王荣富.植物抗寒指标的种类及其应用[J].植物生理学通讯,1987(3):49-55.
[24] Lyons J M.Chilling injury in plants[J].Annu Rev Plant Physioll,1973,24:445-446.
[25] Peter L,Steponkus P L.Role of the plasma membrane in freeing injury and cold acclimation annu[J].Annu Rev Plant Physiol,1984,35:543-584.
猜你喜欢
智慧农业导刊(2022年7期)2022-04-21
今日农业(2021年20期)2021-11-26
烟台果树(2021年2期)2021-07-21
内蒙古林业调查设计(2021年1期)2021-06-29
河北果树(2020年4期)2020-11-26
河北果树(2020年1期)2020-02-09
新疆农业科学(2019年8期)2019-10-21
江苏农业科学(2017年10期)2017-07-21
广西林业科学(2016年2期)2016-03-20
中国果业信息(2013年7期)2013-01-22
