
南方泡桐二倍体及其同源四倍体AFLP 和MSAP 的差异1)
2014-03-05 09:59张晓申范国强邓敏捷董焱鹏赵振利
东北林业大学学报 2014年2期
张晓申 范国强 邓敏捷 董焱鹏 赵振利
(河南农业大学,郑州,450002)
多倍化是植物进化过程中的一种普遍现象。自然界大约70%的被子植物在进化史中经历过一次以上的多倍化过程[1-2]。植物染色体数增加后,基因的剂量和互作效应往往会导致植物一系列的生理生化功能的改变,其最明显特征表现为器官和生物量的增大以及适应生物和非生物胁迫能力的提高[3-4]。研究表明,拟南芥[5]、小麦[6-7]、水稻[8]、棉花[9]和黄瓜[10]多倍化后,DNA 甲基化水平和甲基化模式都发生了变化。然而,这些研究多集中在农作物和1年生草本植物[11-12],而有关木本植物的研究则相对较少[13-14]。泡桐(Paulownia)是我国重要的速生用材和绿化树种之一,然而,泡桐生产中存在的丛枝病发生严重和“低干大冠”等问题,严重影响了农民种植泡桐的积极性和泡桐产业的发展。近年来,我们创新了多个四倍体泡桐新种质[15-20],一方面丰富了泡桐种质资源,另一方面也为筛选出符合人们意愿的泡桐新品种提供了材料支撑。过去,虽然科技工作者开展过四倍体泡桐叶片结构、生长和木材特性等方面研究工作[21-22],但至目前还未见国内外有关泡桐二倍体及其同源四倍体遗传关系研究的报道。为了弄清南方泡桐二倍体染色体加倍前后DNA 的变化情况,阐明南方泡桐四倍体优良特征的分子机制,本文利用DNA 扩增片段长度多态性(AFLP)和DNA 甲基化敏感扩增多态性(MSAP)技术研究南方泡桐二倍体及其同源四倍体的DNA 碱基序列及其甲基化变化,以期为南方泡桐四倍体大面积推广提供理论支撑。
1 材料与方法
试验材料为河南农业大学泡桐研究所经体胚发生途径获得,并培养30 d 的南方泡桐二倍体及其同源四倍体组织培养苗。剪取适量生长状况良好、大小一致幼苗长0.3 ~0.5 cm 的顶芽用液氮冷冻后放于冰箱内-86 ℃备用。组培苗培养温度为(25±2)℃、光照强度2 000 lx、光照时间16 h·d-1。
泡桐DNA 的提取:南方泡桐二倍体及其同源四倍体DNA 的提取参照张延召等[23]方法。
AFLP 和MSAP 扩增:南方泡桐二倍体及其同源四倍体DNA 的AFLP 和MSAP 扩增所需96 对引物组合的碱基序列、扩增程序及其产物的电泳分别参照曹喜兵等[24-25]法。
电泳凝胶的谱带分析:电泳结束后,对用硝酸银染色的胶板谱带进行统计分析。其中,H 和M 分别为EcoRⅠ/HpaⅡ和EcoRⅠ/MspⅠ双酶切产物电泳谱带,每一条谱带代表一个酶切位点,将谱带有或无分别记作1 或0。每个DNA 样品的H 和M 扩增谱带可划分为4 种[种类I(H,M=1,1),无甲基化发生;种类Ⅱ(H,M=1,0),单链DNA 外甲基化;种类Ⅲ(H,M=0,1),双链DNA 内甲基化;种类IV(H,M=0,0),双链DNA 外甲基化]。DNA 甲基化类型分为多态性和单态性类型。DNA 多态性类型为甲基化型(A)、去甲基化型(B)和不定型(C)。其中,A 型中的A1 和A2 代表DNA 重新甲基化(对照样H 和M 泳道均有带,而处理样仅H 或M 泳道有带);A3和A4 代表DNA 超甲基化(对照样仅H 或M 有1 条带,而处理样H 和M 泳道都没带)。B 型(Bl、B2、B3 和B4)代表DNA 去甲基化,DNA 甲基化谱带与A 型相反。C 型代表DNA 甲基化的不确定性(对照样与处理样DNA 甲基化差异谱带无法确定)。单态性类型为D 型(对照样与处理样间DNA 谱带相同)。同时,计算样品的总DNA 甲基化水平[(种类II 谱带数量+种类Ⅲ谱带数量)/(种类I 谱带数量+种类II 谱带数量+种类III 谱带数量)×100%]和总DNA 甲基化多态性[(A 型谱带数量+B 型谱带数量+C 型谱带数量)/(A 型谱带数量+B 型谱带数量+C型谱带数量+D 型谱带数量)×100%]及DNA 单态性[D 型谱带数量/(A 型谱带数量+B 型谱带数量+C 型谱带数量+D 型谱带数量)×100%]。
2 结果与分析
2.1 南方泡桐二倍体染色体加倍后DNA 的碱基序列变化
南方泡桐二倍体及其同源四倍体DNA 的AFLP扩增产物的电泳结果(图1)表明,虽然南方泡桐二倍体及其同源四倍体DNA 经不同引物扩增后产生的片段大小和数量存在一定的差异,但同一引物扩增后产生了数量相同和长度一致的DNA 片段,并且每对引物平均可扩增出50 ~70 条谱带。也就是说,南方泡桐二倍体及其同源四倍体DNA 的AFLP 酶切位点没有发生变化。该结果说明,南方泡桐二倍体及其同源四倍体的DNA 碱基序列在AFLP 水平上是相同的。
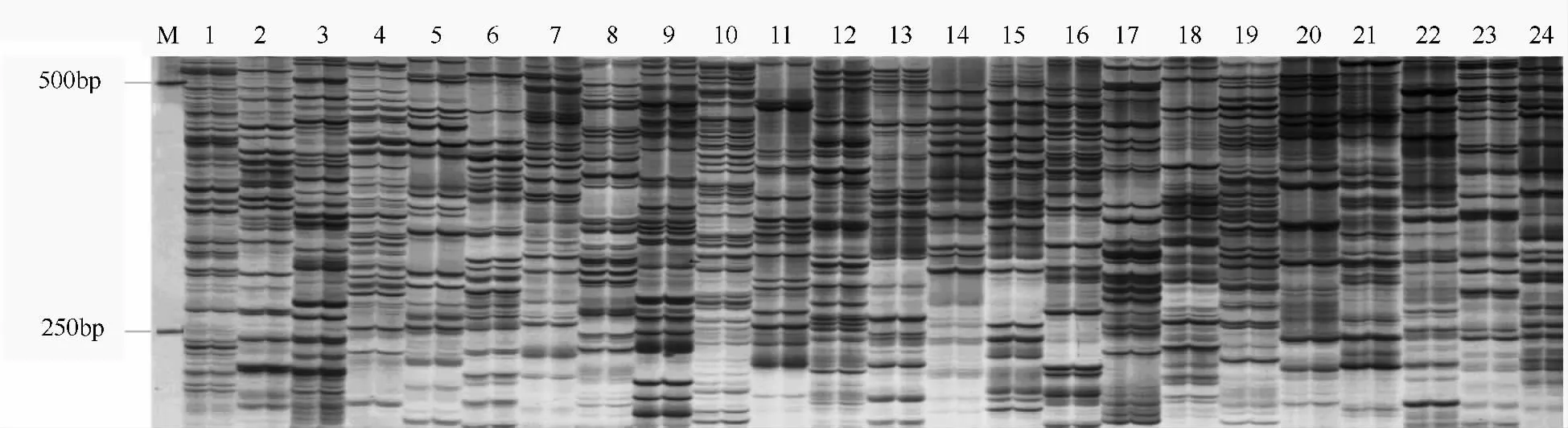
图1 南方泡桐二倍体及其同源四倍体的AFLP 扩增产物
2.2 南方泡桐二倍体及其同源四倍体的DNA 甲基化差异
2.2.1 南方泡桐二倍体及其同源四倍体DNA 甲基化水平的差异
南方泡桐二倍体及其同源四倍体的MSAP 扩增产物的电泳结果(表1)表明,南方泡桐同源四倍体总DNA 甲基化水平高于其二倍体。南方泡桐二倍体DNA 的MSAP 扩增位点为2 059 个,其中全甲基化位点数为201 个(占总扩增带数的9.76%),半甲基化位点数为506 个(占总扩增位点的24.58%),总甲基化位点为707 个(占总扩增带数的34.34%);南方泡桐同源四倍体的扩增位点为2214 个,其中全甲基化位点数为260 个(占总扩增带数的11.74%),半甲基化位点数为554 个(占总扩增位点的25.02%),总甲基化位点为814 个(占总扩增带数的36.77%)。此外,南方泡桐二倍体及其同源四倍体的CCGG 位点均以双链全甲基化(CmCGG)为主,并且南方泡桐同源四倍体DNA 全甲基化和半甲基化水平皆高于其二倍体。结合南方泡桐二倍体染色体加倍前后DNA 碱基序列变化情况可以推出,南方泡桐同源四倍体DNA 甲基化水平升高可能是其生物学和木材理化性质未呈现倍增的主要原因之一。

表1 南方泡桐二倍体及其同源四倍体总DNA 甲基化水平
2.2.2 南方泡桐二倍体及其同源四倍体DNA 甲基化模式的差异
由南方泡桐二倍体及其同源四倍体DNA 的MSAP 扩增产物的凝胶电泳结果(图2)和谱带统计结果(表2)可以看出,南方泡桐二倍体染色体加倍前后的DNA 甲基化模式发生了变化。南方泡桐二倍体DNA 甲基化(A 型)位点数为281(占总甲基化多态性位点数的15.47%),去甲基化(B 型)位点数为449(占总甲基化多态性位点数的24.72%)。去甲基化(C型)位点数为11(占总甲基化多态性位点数的0.61%),与南方泡桐二倍体相比,其四倍体总甲基化多态性比率为40.80%,甲基化单态性(D 型)位点数1075(占总甲基化多态性位点数的59.20%)。该结果说明,南方泡桐同源四倍体DNA 甲基化模式变化频率小于其二倍体,并且南方泡桐同源四倍体DNA 去甲基化模式发生的频率高于其发生甲基化的频率。
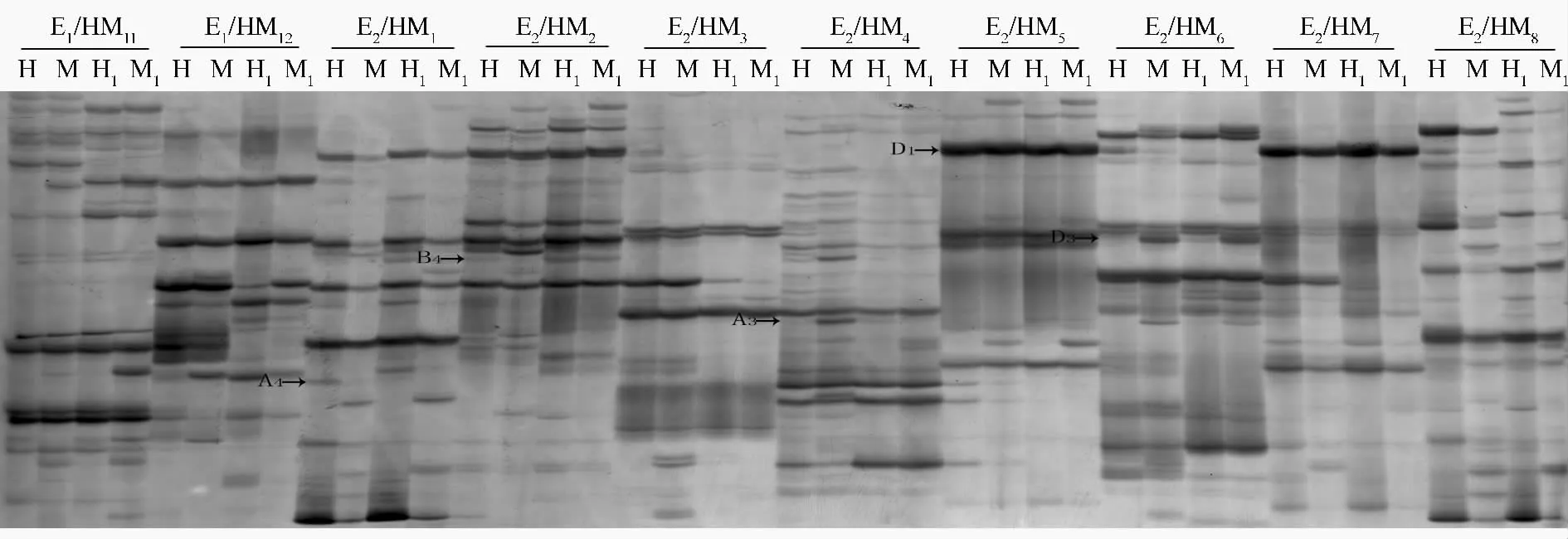
图2 南方泡桐二倍体及其同源四倍体的MSAP 扩增产物
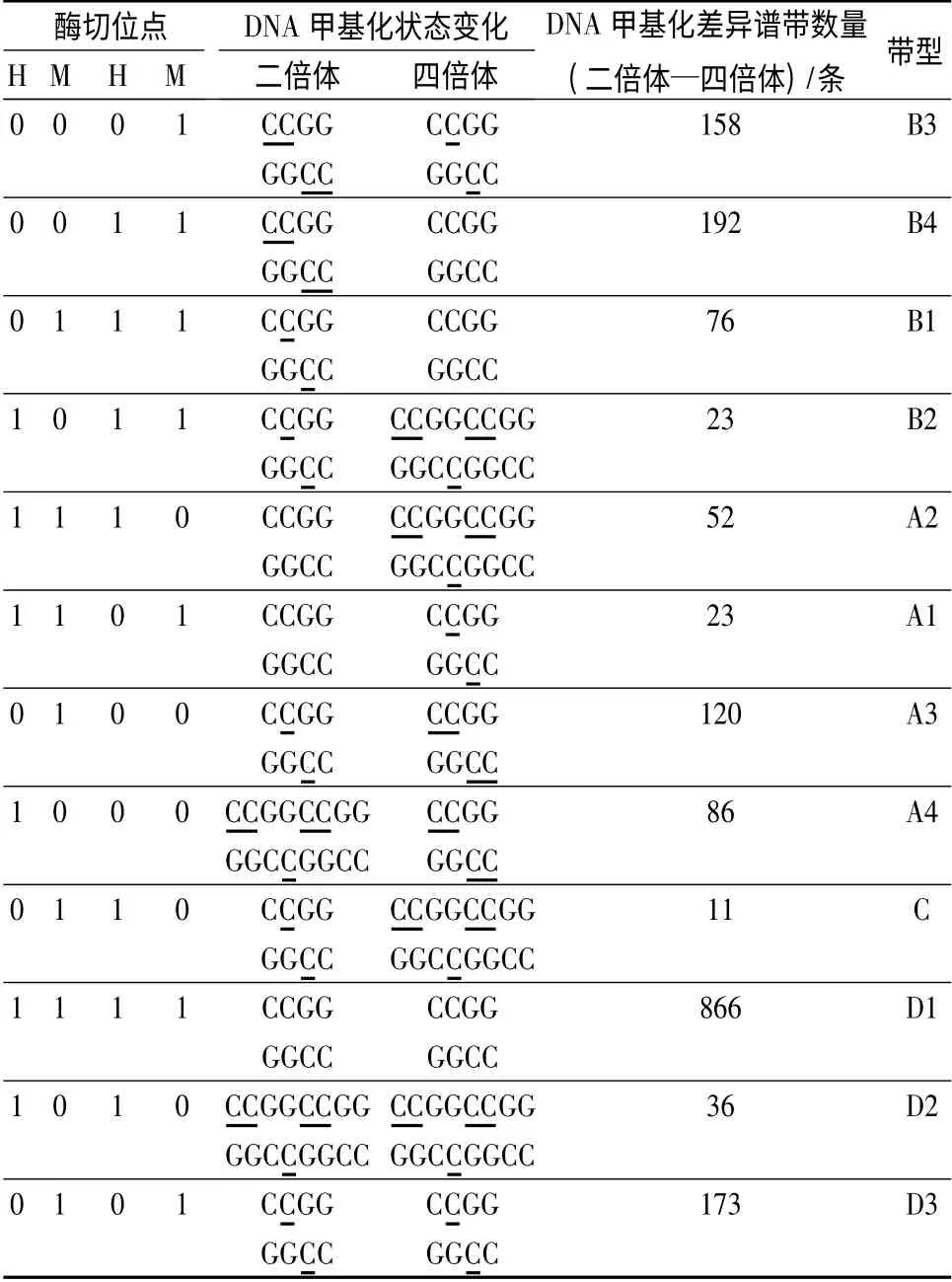
表2 南方泡桐二倍体及其同源四倍体DNA 甲基化模式
3 结论与讨论
多倍化是推动植物进化和新物种形成的重要动力。根据加倍染色体的起源,科技工作者将多倍体划分为同源多倍体和异源多倍体[26-28],四倍体是多倍体中最常见的类型之一[26]。近30 a 来,人们已对异源四倍体与其对应二倍体的遗传变异进行过大量的研究[3,26-31],而对同源四倍体的研究则相对较少[3]。研究表明,同源四倍体植株具有器官和生物量增大的明显特征及较强适应生物和非生物胁迫的能力[3-4],这些变化必然与染色体加倍后植物DNA的变化有一定的联系。植物染色体加倍前后,DNA碱基序列及其甲基化变化情况,不同研究者得出的结果不完全相同。焦锋等[32]利用RAPD 技术分析了桑树(Morus alba L.)二倍体和同源四倍体的DNA指纹,没有发现差异条带;赵卫国等[33]和林强等[34]分别利用ISSR 和PARD 技术研发现,农桑8 号(Nongsang 8)和湖桑199 号(Husang 199)的二倍体及其同源四倍体间扩增条带完全相同;栾丽等[35]发现的研究结果表明,水稻(Oryza sativa)二倍体及其四倍体DNA 碱基序列在SSR 水平上没有发生变化;聂丽娟等[36]利用AFLP 没有检测到西瓜(Citrullus lamatus)二倍体和同源四倍体间DNA 碱基序列的差异;土豆(Solanum tuberosum)二倍体染色体加倍前后DNA 碱基序列的变化情况也是如此[37]。但是,刘文革等[38]和王卓伟等[39]利用AFLP 研究发现,西瓜(Citrullus lanatus)和桑树(Morus alba L.)二倍体染色体加倍前后DNA 碱基序列发生了明显变化,类似结果在黄瓜(Cucumis sativus L.)二倍体及其同源四倍体的研究中得到[40]。本研究中,我们利用AFLP 分析发现,南方泡桐二倍体及其染色体加倍后的DNA 碱基序列没有发生变化。我们认为,造成该结果的原因可能与不同研究者在诱导同源四倍体过程中使用不同剂量诱变剂和不同时间跨度的试验材料有一定关系。如果使用的诱变剂浓度过大,必然引起DNA 结构的破坏,导致同源四倍体染色体的重组或基因重排[41-42]、染色体不等交换及DNA 碱基序列丢失[42-43]情况的发生,从而出现DNA 谱带的变化[37]。此外,植物染色体加倍后,植株细胞内发生了核质不平衡等现象,但又要适应环境条件,完成生长发育生活周期,迫使植物作出适宜的形态和遗传变化[44-46]。同源四倍体获得的时间越短,DNA碱基序列的一致性越高,用AFLP 等技术检测出差异性的可能性越小,但由于染色体加倍造成的DNA甲基化变化始终存在,故出现MSAP 片段多态性就是必然的[47]。同源四倍体植物获得的时间越长,DNA 碱基序列变化可能性越大,故就较容易用AFLP 和MSAP 检测出染色体加倍前后DNA 碱基序列及其甲基化的变化情况[9-12]。在此需要说明的是,虽然AFLP 较能准确反映DNA 碱基序列的变化,但其检测的毕竟仍是DNA 片段。故要证明二倍体及其同源四倍体南方泡桐间DNA 碱基序列是否发生变化还需在DNA 单碱基(SNP)水平上开展研究。
[1] Masterson J. Stomatal size in fossil plants:evidence for polyploidy in majority of angiosperms[J]. Science,1994,264:421-424.
[2] Wendel J F. Genome evolution in polyploids[J]. Plant Mol Biol,2000,42(1):225-249.
[3] Hilu K W. Polyploidy and the evolution of domesticated plants[J].Am J Bot,1993,80:1494-1499.
[4] Liu B,Wendel J F. Non-mendelian phenomena in allopolyploid genome evolution[J]. Current Genomics,2002,3(6):489-505.
[5] Madlung A,Masuelli R W,Watson B,et al. Remodeling of DNA methylation and phenotypic and transcriptional changes in synthetic Arabidopsis allotetraploids[J]. Plant Physiology,2002,129(2):733-746.
[6] Shaked H,Kashkush K,Ozkan H,et al. Sequence elimination and cytosine methylation are rapid and reproducible responses of the genome to wide hybridization and allopolyploidy in wheat[J].The Plant Cell,2001,13(8):1749-1759.
[7] Han F P,Fedak G,Ouellet T,et al. Rapid genomic changes in interspecific and intergeneric hybrids and allopolyploids of triticeae[J]. Genome,2003,46(4):716-723.
[8] Zhang H Y,Peng H,Li Y,et al. Patterns of DNA cytosine methylation between haploids and its corresponding diploids in rice[J].Chinese Science Bulletin,2006,51(14):1721-1728.
[9] Keyte A L,Percifield R,Liu B,et al. Infraspecific DNA methylation polymorphism in cotton (Gossypium hirsutum L.)[J]. Journal of Heredity,2006,97:444-450.
[10] Chen L Z H,Chen J F. Changes of cytosine methylation induced by wide hybridization and allopolyploidy in cucumis[J]. Genome,2008,51(10):789-799.
[11] Ochogavia A C,Cervigni G,Selva J P,et al. Variation in cytosine methylation patterns during ploidy level conversions in Eragrostis curvula[J]. Plant Mol Biol,2009,70(1/2):17-29.
[12] Rodriduez M P,Cervigni G D L,Quarin C L,et al. Frequencies and variation in cytosine methylation patterns in diploid and tetraploid cytotypes of Paspalum notatum[J]. Biologia Plantarum,2012,56(2):276-282.
[13] 胡宝全,王春国,方成泉,等.不同倍性(2X、3X、4X)鸭梨基因组DNA 甲基化水平与模式分析[J].南开大学学报:自然科学版,2011,44(2):32-37.
[14] Bao A L,Hu Q,Xue Z H Y,et al. DNA methylation in genomes of several annual herbaceous and woody perennial plants of varying ploidy as detected by MSAP[J]. Plant Mol Biol Rep,2011,29(4):784-793.
[15] 范国强,杨志清,曹艳春,等.秋水仙素诱导兰考泡桐同源四倍体[J].核农学报,2006,20(6):473-476.
[16] 范国强,杨志清,曹艳春,等.南方泡桐同源四倍体的诱导[J].植物生理学通讯,2007,43(1):109-111.
[17] 范国强,曹艳春,赵振利,等.白花泡桐同源四倍体的诱导[J].林业科学,2007,43(4):31-35.
[18] 范国强,魏真真,杨志清.南方泡桐同源四倍体的诱导及其体外植株再生[J].西北农林科技大学学报:自然科学版,2009,37(10):83-90.
[19] 范国强,翟晓巧,魏真真,等.豫杂一号泡桐体细胞同源四倍体诱导及其体外植株再生[J].东北林业大学学报,2010,38(12):22-26.
[20] 赵振利,何佳,赵晓改,范国强.泡桐9501 体外植株再生体系的建立及体细胞同源四倍体诱导[J].河南农业大学学报,2011,45(1):59-65.
[21] 张晓申,翟晓巧,范国强,等.四倍体泡桐叶片显微结构观察及其抗逆性分析[J].河南农业大学学报,2012,46(6):646-650.
[22] 翟晓巧,张晓申,赵振利,等.四倍体白花泡桐木材的物理特性研究[J].河南农业大学学报,2012,46(6):651-657.
[23] 张延召,曹喜兵,翟晓巧,等.适用于AFLP 分析的泡桐DNA提取方法研究[J].河南农业大学学报,2009,43(6):610-614.
[24] 曹喜兵,何佳,翟晓巧,等.泡桐AFLP 反应体系的建立及引物筛选[J].河南农业大学学报,2010,44(2):145-150.
[25] 曹喜兵,赵改丽,范国强.泡桐MSAP 反应体系的建立及引物筛选[J].河南农业大学学报,2012,46(5):536-541.
[26] Comai L. The advantages and disadvantages of being polyploidy[J]. Nat Rev Genet,2005,6(5):836-846.
[27] Doyle J J,Flagel L E,Paterson A H,et al. Evolutionary genetics of genome merger and doubling in plants[J]. Annu Rev Genet,2008,42:443-461.
[28] Soltis P S,Soltis D E. The role of hybridization in plant speciation[J]. Annu Rev Plant Biol,2009,60:561-588.
[29] Chen Z J,Ni Z. Mechanisms of genomic rearrangements and gene expression changes in plant polyploids[J]. Bio Essays,2006,28(3):240-252.
[30] Chen Z J. Genetic and epigenetic mechanisms for gene expression and phenotypic variation in plant polyploids[J]. Annu Rev Plant Biol,2007,58:377-406.
[31] Li Xiaodong,Yu Erru,Fan Chuchuan,et al. Developmental,cytological and transcriptional analysis of autotetraploid arabidopsis[J]. Planta,2012,236(2):579-596.
[32] 焦锋,楼程富,张有做,等.桑树变异株系基因组DNA 扩增多态性RAPD 研究[J].蚕业科学,2001,27(3):165-169.
[33] 赵卫国,苗雪霞,黄勇平.桑树二倍体及其同源四倍体遗传差异的ISSR 分析[J].蚕业科学,2005,31(4):393-397.
[34] 林强,邱长玉,朱方容,等.桑树二倍体及其人工诱导同源四倍体遗传差异的RAPD 分析[J].南方农业学报,2011,42(1):11-15.
[35] 栾丽,孔繁伦,何涛,等.用SSR 标记检测同源四倍体与二倍体水稻的遗传差异[J].应用与环境生物学报,2004,10(5):556-558.
[36] 聂丽娟,王子成,王一帆,等.二倍体和四倍体西瓜的DNA 甲基化差异分析[J].核农学报,2009,23(1):80-84.
[37] Stupar R M,Bhaskar P B,Yadell B S,et al. Phenotypic and transcriptomic changes associated with potato autopolyploidization[J]. Genetics,2007,176(4):2055-2067.
[38] 刘文革,王鸣,阎志红.西瓜二倍体及同源多倍体遗传差异的AFLP 分析[J].果树学报,2004,21(1):46-49.
[39] 王卓伟,余茂德,鲁成.桑树二倍体及人工诱导的同源四倍体遗传差异的AFLP 分析[J].植物学通报,2002,19(2):194-200.
[40] 张晓青,陈劲枫,雷春,等.不同倍性黄瓜遗传差异的AFLP 分析[J].西北植物学报,2006,26(11):2265-2269.
[41] Ozkan H,Levy A A,Feldman M. Allopolyploidy-induced rapid genome evolution in the wheat (Aegilops-Triticum)group[J].The Plant Cell,2001,13(8):1735-1747.
[42] Lukens L N,Quijada P A,Udall J,et al. Genome redundancy and plasticity within ancient and recent Brassica crop species[J].Biological Journal Linnean Society,2004,82(4):665-674.
[43] Kashkush K,Feldman M,Levy A A. Gene loss,silencing and activation in a newly synthesized wheat allotetraploids[J]. Genetics,2002,160(4):1651-1659.
[44] Finnegan E J,Peacock W J,Dennis E S. DNA methylation,a key regulation of plant development and other processes[J]. Curr Opion Genet Develop,2000,10:217-223.
[45] Xu Yanhao,Zhong Lan,Wu Xiaoming,et al. Rapid alterations of gene expression and cytosine methylation in newly synthesized Brassica napus allopdyploids[J]. Planta,2009,229:471-483.
[46] Martelotto L G,Ortiz J P A,Stein J,et al. Genome rearrangements derived from autopolyploidization in Paspalum sp.[J].Plant Sci,2007,172(5):970-977.
[47] 王春国,古瑜,陈成彬,等.不同倍性西瓜基因组DNA 甲基化水平与模式的MSAP[J].分子细胞生物学报,2009,42(2):118-125.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
数学物理学报(2022年1期)2022-03-16
航天工业管理(2019年11期)2019-04-20
现代园艺(2017年13期)2018-01-19
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
中成药(2016年4期)2016-05-17
中国果业信息(2015年6期)2015-01-23
汽车与新动力(2014年3期)2014-02-27
汽车与新动力(2012年4期)2012-03-25

- 东北林业大学学报的其它文章
- 萱草优良单株花茎离体快繁1)