
影响翅果油树种子萌发及幼苗生长的土壤因素
2014-01-22 01:21翟静娟毕润成
经济林研究 2014年2期
翟静娟,毕润成
(1.朔州职业技术学院,山西 朔州 036002; 2.山西师范大学 生命科学学院,山西 临汾 041004)
翅果油树Elacagnus mollisDiels又名泽绿旦、柴禾、车勾子等,是胡颓子科胡颓子属的大灌木或小乔木,为我国特有种[1-3]。翅果油树有较高的抗旱、耐寒和耐瘠薄能力,20世纪80年代被列为国家第一批二级重点保护植物[4-5]。国内外的相关研究多集中于翅果油树种群、濒危原因、脂肪酸成份等,关于翅果油树种子萌发及幼苗指标方面的研究报道较为鲜见[6]。提高种子的发芽率对有效扩大翅果油树的种植面积具有重要意义。本试验在不同果皮处理方法和不同土壤条件组合下,通过对翅果油树种子萌发和幼苗生长过程中抗氧化酶活性、异柠檬酸裂解酶活性等指标的测定,尝试探讨解除翅果油树种子休眠的最佳方法[7-8],为提高翅果油树种子的萌发率提供依据。
1 材料与方法
1.1 供试种子
翅果油树的成熟坚果(以下简称“种子”)于2006年10月采自山西省临汾市翼城县甘泉林场。该林场位于 35°73′N;111°68′E,海拔800~1 200m,年平均气温为12.5℃,年平均降雨量为550mm,土壤pH为8.4。将除去杂质后的种子放在布袋中贮藏在于4℃环境下备用(International Seed Testing Association,1985)。
1.2 土壤的收集
为了探讨不同土壤条件对种子萌发和幼苗生长的影响,本研究从3个地方获取实验所需土壤,分别为:山西师范大学校内花园的土壤(1)、乡宁县翅果油树生长地的土壤(2)以及翼城甘泉林场翅果油树生长地的土壤(3)。
1.3 果皮处理方法
本试验的果皮处理方法包括:全中果皮处理(A)、尖端开口但不去除中果皮处理(B)、 1/2中果皮处理(C)、去除中果皮但保留表皮毛处理(D)、去除中果皮且不保留表皮毛处理(E)、去除种皮处理(F),共6种处理。
试验共设置18种处理,其中包含3种土壤和6种果皮处理的方法,编号分别是1A、1B、1C、1D、1E、1F、2A、2B、2C、2D、2E、2F、3A、3B、3C、3D、3E和3F(1A为对照)。
1.4 种子的播种
2007年4月,将上述三种土壤分别与细纱混合(混合比例为1∶1)后,分别倒入30个花盆中,共计90盆。花盆规格为深11cm、内径19cm。在每种土壤条件下,每种种子各播种5盆,每盆5株,盆中装土2 kg,放置于温室使种子萌发(25℃,12h光照)。
1.5 指标的测定
在实验开始的第3、6、9、12、15、18天,各挖取一次翅果油树种子,分别测定其超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、异柠檬酸裂解酶(ICL)的活性。翅果油树萌发3周后,测定其幼苗的发芽率、发芽势、萌发指数、根长、芽长、株高和茎粗等各项幼苗指标[9-11]。所有指标的测定均重复3次。各种酶活性的测定参照宋松泉的《种子生物学研究指南》[12]进行。
幼苗指标的测定[13-15]:
发芽率(%)=发芽的种子数×100/供试种子数;
发芽势(%)=规定时间内发芽的种子数×100/供试种子数。
采用精确到1mm的直尺测量株高(子叶节到生长点之间的距离),重复3次。采用游标卡尺测量茎粗(子叶节下1cm处),重复3次。
1.6 数据的统计分析
所有指标的测定均重复3次。采用Excel 2003分析数据并绘制图表。采用t检验进行不同组间差异的比较。
2 结果与分析
2.1 不同土壤处理对翅果油树种子萌发过程中抗氧化酶活性的影响
不同土壤处理对翅果油树种子萌发过程中超氧化物歧化酶活性的影响结果如图1所示。在翅果油树种子萌发过程中,随着萌发天数的增加,SOD活性从3d到18d呈先升后降的变化,9d时后开始急速上升,15d时SOD活性达到最大值,在18d时各处理都达到最低点。其中,3E处理、3D处理、3F处理、2E处理、2F处理以及1E处理的SOD活性高于其它处理组和对照组。图1表明,翅果油树种子的SOD活性随其吸水膨胀以及胚根的出现和伸长逐渐增强。在翅果油树种子萌发两周后,随着胚芽的伸长,SOD活性在其萌发15d时基本达到稳定。采自乡宁和翼城的土壤都不同程度地提高了翅果油树萌发过程中的SOD活性,增强了翅果油树种子抗逆萌发能力。15d时,乡宁和翼城土壤处理都能明显提高SOD活性,(2)土壤处理与1A处理相比差异显著(P<0.05),(2)土壤处理与(3)土壤处理差异极显著(P<0.01)。去除中果皮且不保留表皮毛处理、去除中果皮但保留表皮毛处理去除种皮处理都能有效的提高翅果油树种子萌发过程中SOD活性。
不同土壤处理对翅果油树种子萌发过程中过氧化物酶和过氧化氢酶的影响结果分别如图2和图3所示。植物抗氧化酶系统的另外两种酶POD和CAT活性在翅果油树种子萌发过程中,呈现和SOD相似的先升后降的变化趋势。
2.2 不同土壤处理对翅果油树种子萌发过程中异柠檬酸裂解酶活性的影响
在翅果油树种子萌发过程中,脂肪酸经β-氧化作用,最终转化为糖的途径与乙醛酸循环中的关键酶之一ICL密切相关。不同土壤处理对翅果油树种子萌发过程中异柠檬酸裂解酶的影响结果如图4所示。异柠檬酸裂解酶活性随翅果油树种子吸水膨胀及胚根出现,在9d前逐渐增强。9d时开始急速升高,在12~15d时达到最大值,15d之后ICL活性开始急速下降,在18d时各处理都达到最低点。异柠檬酸裂解酶活性达到最大值时,3E处理的ICL活性最大,从大到小依次分别为3D处理、3F处理、2E处理、2F处理。由此可知,3E处理能有效提高翅果油树种子萌发过程中异柠檬酸裂解酶的活性,并且去除中果皮且不保留表皮毛处理、去除中果皮但保留表皮毛处理去除种皮处理都能有效提高翅果油树种子萌发过程中异柠檬酸裂解酶的活性。翼城土壤处理与1A(CK)的差异极显著(P<0.01),与乡宁土壤处理的差异显著(P<0.05)。

图1 不同土壤处理对翅果油树种子萌发过程中超氧化物歧化酶的影响Fig.1 Effect of different soil treatments on SOD activity in E.mollis seeds during germination
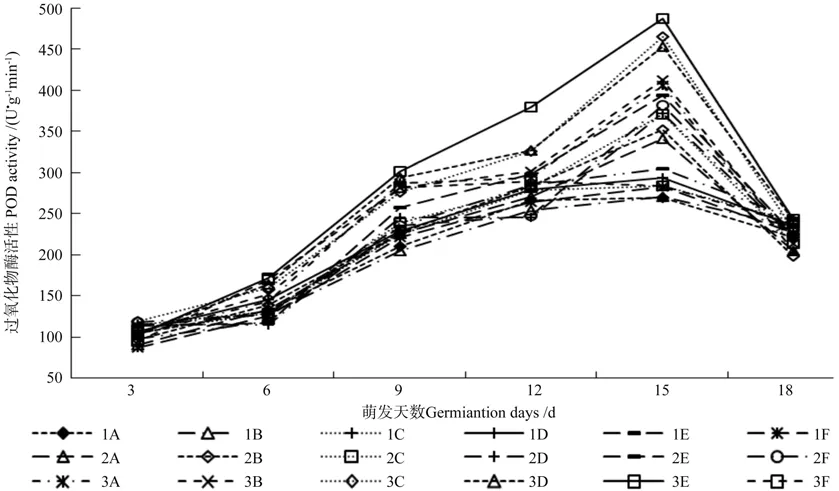
图2 不同土壤处理对翅果油树种子萌发过程中过氧化物酶的影响Fig.2 Effect of different soil treatments on POD activity in E.mollis seeds during germination
2.3 不同土壤处理对翅果油树种子萌发的影响
不同土壤处理对翅果油树种子萌发的影响结果如表1所示。不同土壤处理的翅果油树种子的发芽率、发芽势和萌发指数都较1A(CK)有不同程度的提高。在相同土壤处理下,1E处理、2E处理、3E处理均能最显著地提高翅果油树种子的发芽率、发芽势和萌发指数(表1),表明去除中果皮且不保留表皮毛处理能最有效地提高翅果油树种子的萌发。在不同土壤且相同果皮处理的情况下,3号土壤种植翅果油树种子的发芽率、发芽势和萌发指数都高于其它两种土壤的处理,其次为2号土壤,而发芽率、发芽势及萌发指数最低的是种植在1号土壤的种子。与CK相比,经乡宁的土壤栽植处理后,翅果油树幼苗的发芽率、发芽势和萌发指数差异显著(P<0.05),而经翼城甘泉林场的土壤栽植处理后,翅果油树幼苗的发芽率、发芽势和萌发指数差异极显著(P<0.01)。

图3 不同土壤处理对翅果油树种子萌发过程中过氧化氢酶的影响Fig.3 Effect of different soil treatments on CAT activity in E.mollis seeds during germination

图4 不同土壤处理对翅果油树种子萌发过程中异柠檬酸裂解酶的影响Fig.4 Effect of different soil treatments on ICL activity in E.mollis seeds during germination
2.4 不同土壤处理对翅果油树幼苗生长的影响
不同土壤处理对翅果油树幼苗生长的影响结果如表2所示。表2的数据表明,经不同土壤处理和不同果皮处理的翅果油树幼苗的根长、芽长、株高和茎粗都较1A(CK)有所提高。1E、2E和3E处理的翅果油树幼苗的根长、芽长、株高和茎粗均显著高于其它处理组。在不同土壤、相同果皮处理情况下,3号土壤种植翅果油树幼苗的根长、芽长、株高和茎粗都显著高于2号和1号土壤。与CK相比,经翼城甘泉林场的土壤栽植处理后,翅果油树幼苗的根长、芽长、株高和茎粗差异极显著(P<0.01),经乡宁土壤栽植处理后差异显著(P<0.05)。

表1 不同土壤处理对翅果油树种子发芽率和发芽势的影响†Table 1 Effect of different soil treatments on germinationrate and germination energy of E.mollis seeds %
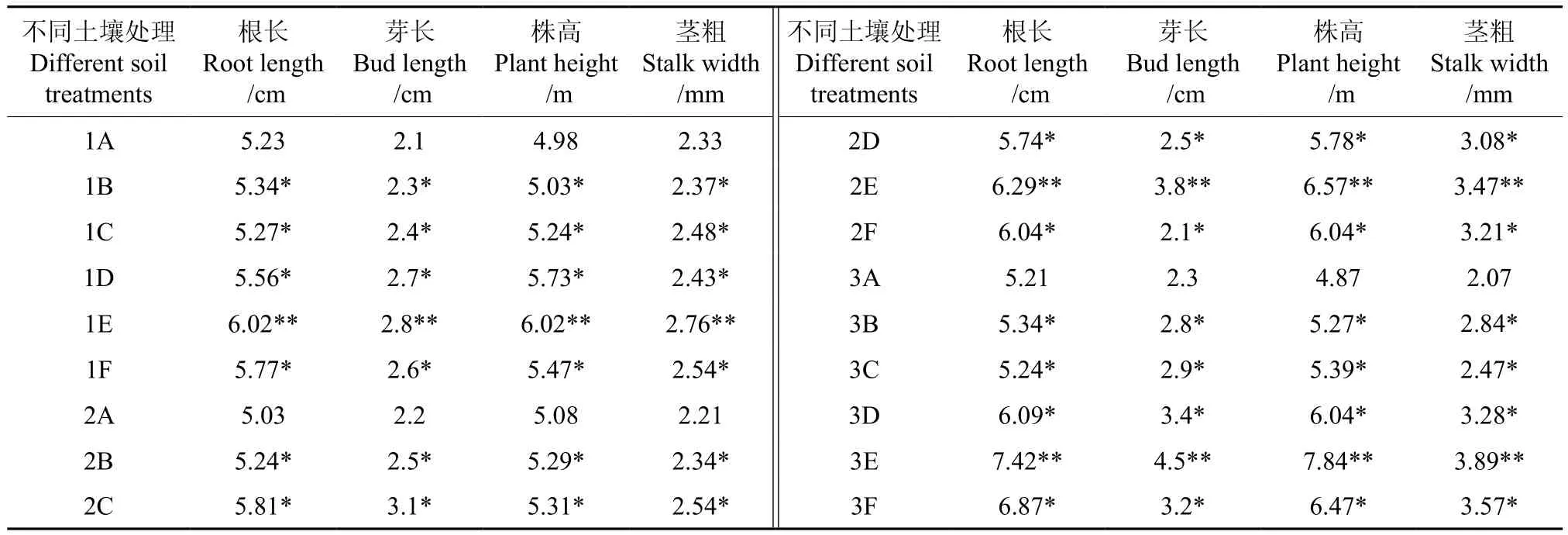
表2 不同土壤处理对翅果油树幼苗生长的影响Table 2 Effect of different soil treatments on seedling growth of E.mollis
3 结论与讨论
翅果油树种子中果皮不易腐烂是造成其种子萌发率低的原因之一。果皮不易腐烂导致种子发芽后很难冲破坚硬果皮的束缚[16]。已有研究表明,未经处理的种子发芽率为5.93%,发芽后能正常发育成籽苗的仅为1.69%。混沙处理的种子发芽率为45.76%,发芽后能正常发育成籽苗的占11.02%。主要原因是种子中的发芽抑制物质使胚处于休眠状态[17]。
在相同的外界环境条件下,不同的土壤处理可以提高翅果油树种子的萌发率。在翅果油树种子萌发过程中,经过不同土壤处理和不同果皮及种皮处理的翅果油树种子的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)3类抗氧化保护酶活性呈先升后降的变化。异柠檬酸裂解酶(ICL)是油料种子在萌发过程中的关键酶之一,在翅果油树种子萌发过程中ICL活性先升后降,在12~15d时达到最大值,在18d时各处理都达到最低点。经不同土壤处理的翅果油树种子的发芽指标和活力指数都较1A处理(CK)有不同程度的提高。1E处理、2E处理、3E处理均能最显著的提高翅果油树种子的发芽率、发芽势和萌发指数,表明去除中果皮且不保留表皮毛处理能最有效地提高翅果油树种子的萌发,且未去除中果皮且未经过任何处理会抑制翅果油树种子的萌发。在相同土壤种植处理下,1E处理、2E处理、2F处理、3E处理、3F处理能显著地提高翅果油树幼苗的株高和茎粗,而1A处理、2A处理、3A处理后的翅果油树幼苗的株高和茎粗都为最低,说明未去除中果皮且未经过任何处理会抑制翅果油树种子的萌发。在相同土壤种植处理下,1E、2E、3E处理后的翅果油树幼苗的根长和芽长均显著高于其它处理组。
文中试验结果表明,对翅果油树种子进行不同土壤处理和对果皮进行不同处理均可以促进濒危植物翅果油树种子的萌发。其中,1E、2E、3D和3E处理都能明显地促进翅果油树种子的萌发和幼苗的生长。由此可知,去除中果皮且不保留表皮毛处理、去除中果皮但保留表皮毛处理、去除种皮处理都能有效提高翅果油树种子的萌发率,促进幼苗的生长。经3种不同土壤处理的结果表明,使用翅果油树生长地乡宁和翼城甘泉林场的土壤有助于提高翅果油树种子萌发过程中超氧化物歧化酶的活性,其中以翼城甘泉林场的土壤为优。
[1] 杨克明,刘珍贵,杨天恩,等.高寒阴湿山区翅果油树引种育苗与造林试验[J].经济林研究,2010,28(4):104-107.
[2] 任国臣.濒危植物翅果油树种子蛋白多样性研究[D].临汾:山西师范大学,2010.
[3] 智旭丹.翅果油树组织培养及其器官发生的细胞学研究[D].长沙:湖南农业大学,2008.
[4] 陈 惠,卢英梅,申峻沛.不同栽培基质对翅果油树种子萌发的影响及不同种质幼苗生长发育的表型研究[J].山西师范大学学报,2010,24(3):63-66.
[5] 孙晓春.翅果油树叶片中生物碱酶法提取、分离表征及化感研究[D].临汾:山西师范大学,2012.
[6] 马引利,闫桂琴,陈 洁.外源一氧化氮供体浸种对翅果油树种子萌发和幼苗生长的影响[J].西北林学院学报,2007,22(1):5-7.
[7] 程 鹏,王 平,孙吉康,等.植物种子休眠与萌发调控机制研究进展[J].中南林业科技大学学报,2013,33(5):52-58.
[8] 沈素贞,梁文斌,宋 雷.野生观赏植物短梗大参种子萌发特性的研究[J].中南林业科技大学学报,2013,33(7):57-62.
[9] 曹基武,刘春林,吴 毅,等.不同处理方法对南方红豆杉苗木生长发育的影响[J].中南林业科技大学学报,2013,33(10):9-14.
[10] 李俊南,雄新武,习学良,等.植物激素对薄壳山核桃种子萌发及幼苗生长的影响[J].经济林研究,2013,31(1):81-86.
[11] 殷丽青,朱建军,陈必胜,等.上海地区山桐子种子育苗技术研究[J].经济林研究,2012,30(3):111-114.
[12] 宋松泉.种子生物学研究指南[M].北京:科学出版社,2005.
[13] 沈 凌,丁贵杰.1、2代马尾松林土壤浸提液对马尾松种子萌发及芽苗生长的影响[J].中南林业科技大学学报,2012,32(5):59-62.
[14] 阮长林,冯 剑,刘 强,等.水黄皮种子发芽试验的初步研究[J].中南林业科技大学学报,2013,33(4):38-42.
[15] 洑香香,周晓东,刘红娜.山茱萸种子休眠机理与解除方法初探[J].中南林业科技大学学报,2013,33(4):7-12.
[16] 赵悦茗,王建中.翅果油树资源研究进展[J].河北林果研究,2007,02(5): 154-158.
[17] Delouche J C,Baskin C C.Accelerated aging techniques for predicting the relative storability of seeds lots[J].Seed science and Technology,1973,1(2):427-452.
猜你喜欢
现代园艺(2020年19期)2020-10-02
中国油脂(2020年9期)2020-09-16
中国油脂(2019年7期)2019-08-22
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
种子科技(2018年11期)2018-09-10
许昌学院学报(2018年8期)2018-09-05
天津农林科技(2015年1期)2015-12-30
塑料包装(2014年3期)2014-05-07
