
南岭小坑天然次生林生态系统生物量的估算
2013-12-27 04:57黄志宏陈步峰周光益吴仲民梁瑞友
中南林业科技大学学报 2013年8期
黄志宏,陈步峰 ,周光益 ,吴仲民 ,王 旭 ,李 根 ,梁瑞友
(1. 中南林业科技大学 生命科学与技术学院, 南方林业生态应用技术国家工程实验室, 湖南 长沙 410004;2. 中国林业科学研究院热带林业研究所,广东 广州510520; 3. 韶关市曲江区国营小坑林场,广东 曲江512162)
南岭小坑天然次生林生态系统生物量的估算
黄志宏1,陈步峰2,周光益2,吴仲民2,王 旭2,李 根2,梁瑞友3
(1. 中南林业科技大学 生命科学与技术学院, 南方林业生态应用技术国家工程实验室, 湖南 长沙 410004;2. 中国林业科学研究院热带林业研究所,广东 广州510520; 3. 韶关市曲江区国营小坑林场,广东 曲江512162)
粤北南岭是我国中亚热带生物多样性热点地区,森林覆盖率高,具有典型的中亚热带常绿阔叶林群落。本研究在进行样地测定的基础上,采用传统的样地收获法实测了小坑流域地带性植被天然次生林3块代表性样地(小红栲-木荷群落、藜蒴栲-小红栲群落、木荷-杉木群落)生物量,在此基础上,应用相对生长方程法估算了该生态系统生物量。结果表明:(1) 群落是以藜蒴栲、小红栲与木荷为主要建群种,为地带性植被类型;杉木也占有重要地位,这与其中一个群落由杉木林皆伐转换而来有关;群落物种的重要值分布符合正态分布;(2)胸径与地上生物量的曲线拟合回归方程B (kg·tree-1) = 0.128* D2.3678可获得二者较高的相关性(p=0.000 1);树高与胸径关系符合H=3.712 0D0.4746(p<0.000 1);(3) 地上生物量的估算结果为119.720 Mg·hm-2,比实测值高2.59%;地下生物量的估算结果为 23.922±4.201 Mg·hm-2,系统总生物量为 143.642 ±8.896 Mg·hm-2。
天然次生林;地上生物量;地下生物量;相对生长方程;南岭
森林在全球碳循环中起着陆地碳库(Carbon sinks)重要作用[1]。据估算,大约陆地上碳的86%和土壤碳的73%贮藏于森林中[2]。因为森林资源连续清查(Forest inventories)的缘故,森林地上生物量(Aboveground forest biomass)作为碳库理所当然地为人们所熟知[3]。关于森林地上生物量的研究很多,森林生物量可以通过野外测量和遥感与GIS方法估算[4-5],如对热带成熟林与次生林的生物量积累研究[6]等。Brown等[7-8]认为,利用综合生长方程(Improved allometric equations)是研究单木、样地和区域地上生物量的标准方法。因此,综合生长方程被认为是生物量估算中应用最为广泛的方法[9]。最近,Vashum等[1]对林分生物量估算方法进行了综述,认为整体生长模型方法和遥感-GIS方法是其中最为广泛使用的。即便如此,关于不同地带性森林碳储量能力,除温带和北方森林外,其他区域所知甚少,因而碳储量的“变化”,更可能是改进了的信息,不是真正地在碳储存的变化[10]。可见,估算的精确性问题一直是生物量研究的关注焦点之一。然而,估算精度的提高有赖于多项技术水平的提高与应用,包括样地技术、连清资料、改进综合生长模型、遥感技术等[11]。
广东南岭地处粤北韶关,属中亚热带季风气候区,特殊的地理位置等因素使得其具有很高的森林覆盖率,并成中国生物多样性重点保护地区之一[12]。然而,有关这一地区地带性森林的生物量报道尚不多见。李根等[13-14]报道了南岭小坑藜蒴栲群落和木荷群落地上部分生物量。本文对南岭小坑流域天然次生常绿阔叶林群落系统生物量进行估算,以期进一步充实这一特殊区域的生物量研究,为将来探讨该区域森林碳储量与碳汇功能估算提供基本参考。
1 材料与方法
1.1 研究地概况
研究地小坑位于粤北山区北缘,地处南岭山地大庾岭南缘(24°39′~ 24°42′N,113°49′~113°52′E)。地貌特征以中低山为主,局部为丘陵。成土母岩为石灰岩与变质花岗岩。海拔200~1 300 m之间,坡度在15°~40°,局部地大于40°。气候属中亚热带季风气候区,年平均气温18.8℃~21.6℃;最冷月份(1月)平均气温8℃~11℃;最热月份(7月)平均气温28℃~29 ℃;无霜期310 d左右,部分年份冬季有雪。流域内雨量充沛,年降雨量1 400~2 400 mm,3~8月为雨季,9~2月为旱季;年蒸发量1 530 mm,年均相对湿度79%。地带性土壤为山地红壤,土层较薄,多数不足1 m。地带型植被类型为中亚热带季风常绿阔叶林,群落外貌终年常绿,物种组成多样,层次结构复杂。森林总面积16 000 hm2,森林覆盖率85%, 小坑国家森林公园有野生植物162科、216属、772种,以壳斗科、樟科、茶科、金缕梅科、木兰科的常绿阔叶树种为林分的建群种和优势种。
1.2 数据采集
1.2.1 标准地调查
样地踏查以后,选择3块各800 m2的样地作为标准地,记录海拔、坡度、坡向、坡位、土壤等立地因子,做标准地调查。将每块标准地分成8个10 m×10 m的样方,进行乔木层调查,再在每个10 m×10 m的样方内分别选择一个2 m×2 m和一个1 m×1 m的小样方做灌木层和草本层调查。乔木层进行每木调查,记录树名(Species)、树高(H /m)、胸径(DDBH/cm)、第一活枝下高(Hb/m);灌木层记录各树种株数、平均株高、平均地径、盖度;草本层记录各草种平均高度、盖度。藜蒴栲样地为山地红壤,其余两样地为山地黄壤。
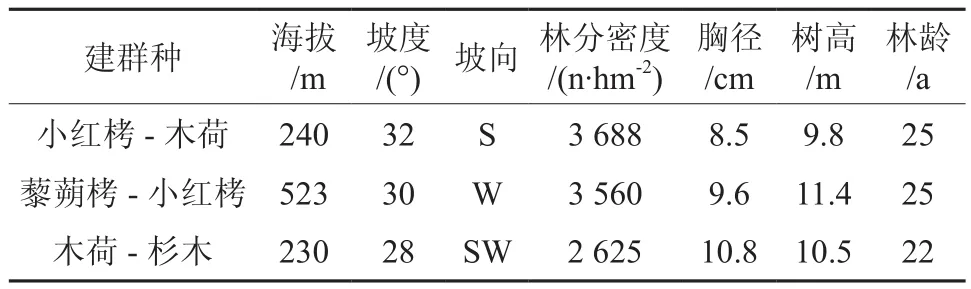
表1 样地基本情况Table 1 Basic background of sampled plots
1.2.2 生物量测定
生物量测定方法:灌木层、草本层、枯枝落叶层生物量测定采用直接收获法,乔木层生物量测定采用皆伐法[15]。其中:
(1)枯枝落叶层生物量测定。在每个10 m×10 m的样方内选取一块1 m×1 m的小样方,共8个小样方,并收集每个小样方内的枯枝落叶测定鲜重。
(2)草本层生物量测定。在每个收集枯枝落叶层的小样方内收集全部草本活体,测定鲜重。
(3)灌木层生物量测定。在每个10 m×10 m的样方内选取一块2 m×2 m的小样方,从植株基部砍伐,分别收集叶、枝、茎测定鲜重。
(4)乔木层生物量测定。乔木层生物量采用皆伐法测定,逐株伐倒,记录树名,测量树高、枝下高、胸径。枝叶剪除全部称量,树干采用Monsi分层切割法,从基部开始,以2.0 m为一个区分段,分段称量。在树木采伐过程中,干型通直的树木作为解析木,测量树高、枝下高、胸径、树木1/4高处的围径、树木1/2高处的围径、树木3/4高处的围径;枝叶鲜重全部测定后,选取其中5%~10%,摘除树叶,换算枝叶比例;树干分段称重,并截取圆盘。
所有鲜重样品均取300 g带回实验室,于105℃恒温干燥箱中烘干24 h后至恒重,求出含水率,将各器官鲜重换算成干重。
1.3 数据统计
根据野外样地实测资料,计算乔木层各树种的立木密度、胸高断面积(AB)、平均胸径(D)、平均树高(H)和相对密度(AR)、相对频度(FR)、相对显著度(SRD)以及重要值(VI) 等指标。统计样地地上部分生物量与胸径、树高的相关关系。统计采用Excel 2003与SAS8.02软件相结合进行。
2 结果与分析
2.1 群落特征
样地调查表明,样地群落结构较为复杂,可分为乔木层(Ⅰ、Ⅱ)、灌木层、草本层以及层间植物。本研究中的3块抽样样地,分别以荷木-杉木、小红栲-荷木、藜蒴栲-小红栲为优势种的群落,各样地群落乔木层平均胸径、平均树高等存在一定差异,如藜蒴栲-小红栲样地平均高度13~15 m,乔木层的主要优势树种为藜蒴栲、小红栲[13];荷木-杉木样地平均胸径是10. 8 cm,平均树高10. 5 m[14];小红栲-荷木样地平均树高16~18 m,乔木层的主要优势树种为小红栲、荷木。林下植物中微毛山矾、罗浮柿等较多。草本植物比较稀疏,多为芒萁,层间植物多为鸡血藤、酸藤子Embelia laeta。
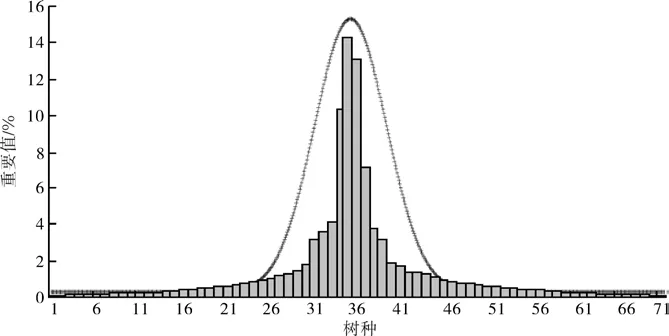
图1 群落中各树种重要值†Fig. 1 Distribution of important values of each tree species in sampled community† 1毛冬青Ilex pubescens 2赛山梅Styrax confusus 3长花厚壳树Ehretia longiflora 4罗浮栲Castanopsis fabri 5杨梅叶青冈Cyclobalanopsis myrsinaefolia 6鸭脚木Schefflera octophylla 7黄杞Engelhardtia roxburghiana 8樟树Cinnamomum camphora 9猴欢喜Sloanea sinensis 10中华杜英Elaeocarpus chinensis 11深山含笑Machelia maudiae 12马尾松Pinus massoniana 13泡花润楠Machilus pauhoi 14东京野茉莉Styrax tonkinensis 15石木木姜子Neolitsea chuii 16罗浮椎Castanopsis fabri 17金毛石栎Lithocarpus chrysocomus 18狗骨柴Diplospora dubia 19青冈栎Cyclobalanopsis glauca 20密花山矾Symplocos congesta 21柳叶润楠Machilus salicina 22香粉树Machilus pauhoi 23越南安息香Styrax tonkinensis 24东南石栎Lithocarpus harlandii 25毛桃木莲Manglietia moto 26羊角杜鹃Rhododendron cavaleriei 27拟赤杨Alniphyllum fortunei 28红润楠Machilus thunbergii 29枫香Liquidambar formosana 30广东润楠Machilus kwangtungensis 31微毛山矾Symplocos wikstroemiifolia 32罗浮柿Diospyros morrisiana 33山乌桕Sapium discolor 34小红栲Castanopsis carlesii 35藜蒴栲Castanopsis fissa 36荷木Schima superba 37杉木Cunninghamia lanceolata 38千年桐Aleurites montana 39南酸枣Choerospondias axillaris 40灰岩润楠Machilus calcicola 41尖叶柃Eurya acuminaissima 42黄樟cinnamomum parthenoxylon 43白椆Lithocarpus glabra 44野漆树Toxicodendron succedaneum 45毛栲Castanopsis rufotomentosa 46鸭脚木Schefflera minutistellata 47鼎湖血桐Macaranga sampsonii 48东南栲Castanopsis jucunda 49冬桃Elaeocarpus duclouxii 50越南山矾 Symplocos cochinchinensis 51青钩Castanopsis kawadamii 52钩椎Castanopsis tibetana 53网脉山龙眼Helicia reticulata 54格药柃Eurya muricata 55岭南槭Acer tutcheri 56甜槠Castanopsis eyrei 57中华槭Acer sinense 58小果八角枫Alangium faberi 59少花桂Cinnamomum pauciflorum 60青冈Cyclobalanopsis glauca 61木棉Bombax malabaricum 62山苍子Litsea cubeba 63大果马蹄荷Exbucklandia tonkinensis 64水青冈Fagus longipetiolata 65软枣红豆Ormosia semicastrata 66尖尾柃Eurya acuminatissima 67桔梗大参Macropanax rosthornii 68华南吴茱萸Evodia austro-sinensis 69桃叶石楠Photinia prunifolia 70罗伞Ardisia quinquegona 71中华杜英Elaeocarpus chinensis
由3块样地组成的近2 400 m2的群落上有乔木树种71种,乔木层DBH>3 cm的树平均胸径9.5 cm,平均树高10.5 m。3样地所在的群落中重要值(IV)较大的分别是小红栲Castanopsis carlesii(10.315)、藜蒴栲 Castanopsis fissa(14.263)、木荷Schima superba(13.116)、杉木Cunninghamia lanceolata(7.088)。这4个树种重要值分别占群落重要值总和的10.5%、14.5%、13.4%和7.2%。前三者均是南岭小坑流域地带性树种,也是群落的建群种。杉木在群落中也占有一定的重要性,但其本身并不是地带性树种,而是因为群落前身为杉木人工林皆伐而来,故群落中还散生有一定数量的杉木。群落中各树种的重要值分布见图1所示。值得指出的是,群落树种的重要值分布符合正态分布。
2.2 地上生物量估算
相对生长方程的函数形式有多种形式。本研究中选择两个方面的函数形式来进行探讨。其一是胸径与地上生物量相关关系,其二是树高与胸径相关性分析。首先是胸径与地上生物量相关关系。根据实测胸径与单株生物量数据,对二者分别取对数后作散点图,然后进行曲线拟合,结果如图2所示。图2中(a)、(b)、(c)和(d)分别是样地荷木-杉木、小红栲-荷木、藜蒴栲-小红栲群落及由3样地综合得到的大样本群落的胸径与单株生物量对数关系图。从图2中可见,胸径与生物量对数曲线呈显著线性关系(p=0.000 1),且回归系数变化很小,变化范围在2.313 2~2.381 3之间。
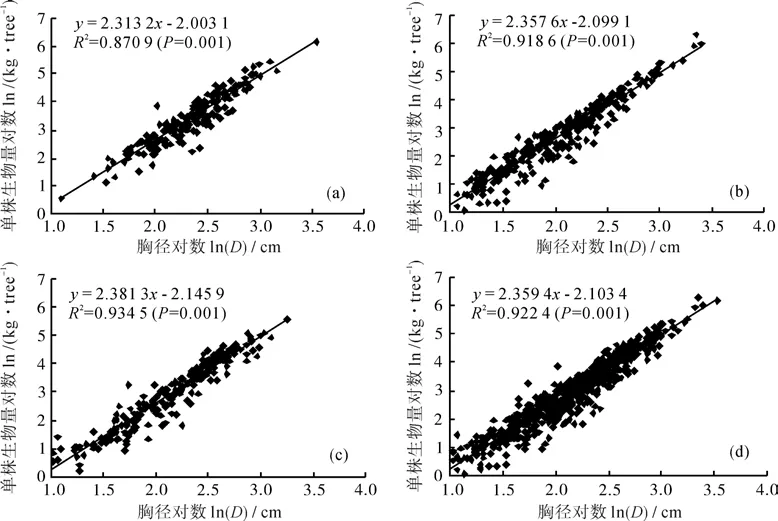
图2 胸径与单株生物量对数关系Fig. 2 Logarithmic relations between DDBH and BAS for all calibration trees in sampled community
取综合样本群落的胸径与其对应生物量的对数,进行曲线拟合,得到如下回归方程:
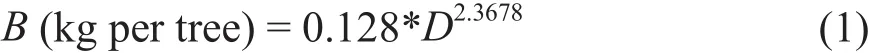
方程(1)中系数a和b的标准误分别为0.003和0.024,方程与系数均达到极显著水平(p=0.000 1),相关系数R2=0.9224。
在生物量幂函数方程(1)中,可将生物量分成两个部分,即B=μ+ε。对于单株而言,其中μi=aDib,μi为估算生物量,εi为估算误差。将样地实测数据按不同径级范围分别拟合其生物量估算方程。同时,拟合了不分径级时的总体生物量估算方程,其拟合参数结果如表2。
树高与胸径关系。树高与胸径的关系在生物量估算方程中通常以D2H的形式出现。在生物量回归模型通常将胸径-树高(D2H)作为自变量,根据相对生长方程式W=a(D2H)b来拟合地上部分各器官生物量和林分生物量。可见树高H在生物量估算中具有重要作用。将实测的765株伐倒木按胸径分级,样木胸径范围在1.6~34.4 cm之间,树高在1.6~18.8 m之间。将所有胸径与生物量分别取自然对数后作图,曲线拟合后得到如图4所示结果。
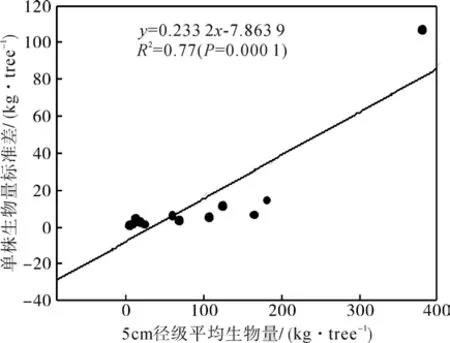
图3 胸径径级与单株生物量估算偏差度较验Fig.3 Standard deviation of tree biomass per 5 cm diameter size classes as a function of mean biomass for all of calibration trees from sampled community

表2 生物量幂函数模型Bi=μi+εi 及μi=aDb与var(εi)=ψμi2的拟合参数值Table 2 Fitting parameters of a power model Bi=μi+εi with μi=aDb and var(εi)=ψμi2
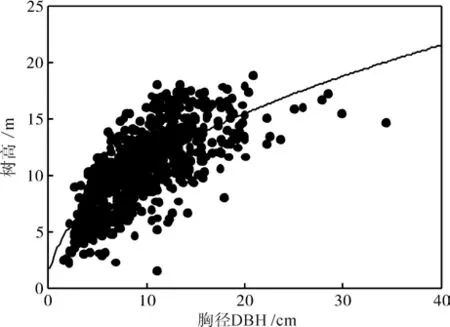
图4 树高-胸径相关关系Fig. 4 Relationship between tree height and diameter at breast height
采用通用方程H =kDc[16]形式,对实测数据进行幂函数曲线拟合,得到:
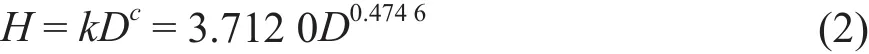
其中H为树高(m),D为胸径(cm)。k和c的标准误分别为0.152 3和0.017 1,二者达到95%的置信区间(p<0.0001)。在胸径与生物量回归方程(1)中得到参数b值为2.367 8,此处树高与胸径回归方程(2)中得到参数c值0.474 6,二者与b=2+c相吻合。因此,本研究中采用形如[17]:

方程来估算生物量,其中c通过样地树高与胸径曲线拟合求得。
在相对生长方程(3)中,D是树木胸径(cm),c是一个反映树高与胸径关系的参数,a是一个通过样地资料拟合得到的参数。单株生物量可看作其体积与其密度之积。因此,假定方程(3)中:

其中r 是一个常数,ρ为样地木材平均密度。根据样地实测数据,样地的平均木材密度为0.64±0.11 kg·dm-3。r值为 a/ρ,根据表 2 中 a 值0.1280,算出r值为0.19。在此,我们给出小坑流域单株生物量估算方程式为:

其中各参数的意义如前所述。c和ρ均可从样地调查资料中估算得到。方程(5)中c=0.4647,ρ=0.51。根据方程(5)估算得到各径阶生物量,同时与实测各径阶生物量比较结果如图5所示。
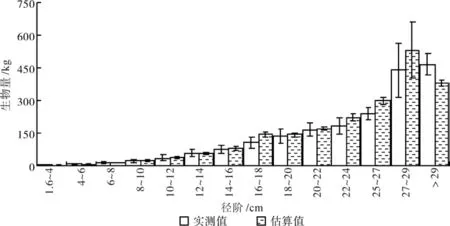
图5 不同径阶生物量估算值与实测值的比较Fig. 5 Comparison between predicted biomass and measured biomass according to different classes of DBH among sampled plots
总体来看,与实测值116.695 Mg·hm-2相比,估算结果为119.720 Mg·hm-2,比实测值高2.59%,表明估算方法是可行的。
2.3 地下生物量估算及系统生物量
地下生物量的估算根据不同的研究结果有不同的关系式。根据IPCC[16]提供的根茎比0.205(SE=0.036),南岭小坑流域天然次生林地下生物量估算为 116.695 Mg·hm-2×0.205=23.922±4.201 Mg·hm-2。
综合地上生物量与地下生物量结果,小坑流域天然次生林生态系统总生物量为143.642 ±8.896 Mg·hm-2,CI(95%置信区间)为137.093~150.190 Mg·hm-2。
3 讨 论
3.1 群落特征
反映群落特征的指标比较多,本研究中仅从重要值这一角度来说明。之所以选择重要值这个指标,主要是考虑到重要值本身就是一个比较综合的指标,能反映群落的许多特性。南岭小坑流域天然次生林的群落特征主要表现为林分的建群种以地带性特征优势种为主要组成部分。如样地木荷花群落中乔木树种20 种,其中木荷、杉木、微毛山矾、灰岩润楠、南酸枣、千年桐Aleurites montana等11个种为主要树种,重要值之和为261.71,占乔木层的87.24% ,另外9 个树种占12.76%[13]。在藜蒴栲群落,优势树种藜蒴栲和小红栲生物量占乔木层地上部分总生物量的46.3%和9.8%[14]。本研究也表明,3块样地分别是以地带性特征种为建群种的群落。将3块样地综合为一个大样地群落时,由子样地的建群种仍为大样地的建群种主要成分。杉木本身并不是地带性树种,但有的林地前身为杉木林,经皆伐杉木林地后转化而来,因而杉木仍占有一定比重。这是林地转化的结果。
本研究还发现,由3块子样地组成的大样地树种数量分布符合正态分布特征(图1)。这种正态分布特征对于天然次生林而言是显而易见的。这种正态分布也表明本研究中样地的选择能反映该流域的地带性植被分布状况。同时,这也是接下来的生物量研究结果能准确反映当地的林分自然状况的前提。
3.2 地上生物量
相对生长方程有多种形式,应用最广泛的有多项式和幂函数模型两种形式[16]。幂函数形式早先由Huxley(1932)[19]在生物学研究中发现普遍存在。本研究中首先采用幂函数形式的常见形式之一B=aDb对样地实测数据进行拟合,并分别从3块子样地与综合样地两个方面得到其拟合参数(见图2),均有较高的相关系数且显著性p<0.0001。这表明这种幂函数形式可以获得比较理想的效果。李根等[13-14]在此地的研究应用B=aDb幂 函 数 得 到Wtagb = 0.116*D2.384,R2=0.934与Wtagb=0.12*D2.479,R2=0.949。本研究中系数b变化范围在2.313 2~2.381 3之间,综合幂函数方程B = 0.128*D2.3678,系数与上述结果相近。
可见,D的系数b是在一定范围内变化的,这种变化可能包括样地本身的变化,并且对于特定的样地而言,其系数b应是相对稳定的。从此角度出发,考虑将样地树高因子加入到相对生长方程中来估算生物量,采用方程H = kDc的形式得到H=3.7120*D0.4746。Brown等[7]考虑树高因子对热带森林生物量估算的影响,针对不同性质的热带森林,分别给出H = 2.918 3*D0.5677(Moist forests)和H = 3.325 7*D0.5627(Wet forests)。可见,系数c的变化反映的是特定立地条件下特定群落高度与群落总体胸径的相对关系,随着群落的变化而发生变化。
在此,采用Kettering等[16]提出的B=aD2+c方程形式,将树高因子加入相对生长方程中来,得到地上生物量估算方程B=aD2.4746。这一方程中系数a分解为更为广义的形式a = rρ(方程式4),其中r为一常数,ρ为木材密度。木材密度在样地生物量估算中是一个重要的因子,有着广泛的应用。如Komiyama等[20]在芒果树生物量研究中得到Wtop = 0.251ρD2.46,这一结果与本研究结果较为相近。
木材密度对于热带次生林与老龄林碳估算是一个极为关键的变量[21],将材积转换为生物量过程中,木材密度是影响生物量估算的关键因子[22]。尽管如此,关于木材密度的取值,这是一个较为困难的事。群落由各种不同的树种组成,不同的树种有不同的木材密度。本研究中通过对样木的测定分析,得到样地平均木材密度为 0.64±0.11 kg·dm-3,但在生物量估算中,发现使用这个值估算的生物量值比实测生物量高出26.7%。这与在计算木材平均密度时仅仅是简单将各测定值相加后取平均值而未考虑不同树种的权重有关,从而导致计算结果偏高。Kettering等[16]曾报道在印度林地木材密度0.60 kg·dm-3(0.53~0.67 kg·dm-3)且不同树种密度变化在0.35~0.91 kg·dm-3之间。Brown等[23]报道亚马逊森林木材密度在0.52~0.80 kg·dm-3之间,平均值为0.67±0.09 kg·dm-3。此后 Brown 等[24]基于 428 个树种分析推荐 0.57 kg·dm-3(0.40 ~ 0.69 kg·dm-3)作为亚洲树种平均木材密度。若采用此推荐值计算得到的地上生物量估算较高实测值高14.7%。Liang等[25]报道广西23 a生藜蒴栲木材密度范围在 0.462 ~ 0.583 g·cm-3间,均值为 0.517 g·cm-3。结合本样地群落建群种为藜蒴栲,且林龄、地域的相近性,本研究中取木材平均密度0.51 g·cm-3。以此值估算得地上生物量为119.720 mg·hm-2,仅比实测值高2.59%。这一结果表明估算方法以及木材密度的取值均是合理的。
3.3 地下生物量
地下生物量的了解对于增进陆地生态系统碳分配与贮存认识非常必要的[26]。地上生物量与地下生物量的关系,即所谓的根茎比(The ratio of root to shoot)。该指标已成为通过相对容易测定的地上生物量来估算地下生物量的一种主要方法[27],并根据不同的植被类型给出森林地下与地上生物量之比0.26。Cairns等[26]提出一个描述根茎关系通用方程y(root)=0.489 x(shoot)0.890来通过地上生物量估算地下生物量。这个通用方程对于估算地下生物量提供了有益的参考。本研究中采用IPCC[18]给出的参考值0.205(S.E.=0.036)(范围0.06~0.33),该参考值对于地上生物量低于125 g·hm-2的热带亚热带林地适用。陈章和等[28]发现广东省黑石顶森林地下与地上生物量之比在0.214~0.282之间,并给出南亚热带、亚热带地区森林根茎比在0.19~0.27之间。根茎比受多种因素的影响,如气候、土壤质地等非生物因素影响[29],还与林龄有关。一般随林龄增大而呈现增加趋势[30]。小坑流域天然次生林林龄在22~25 a之间,因此小坑流域地下-地上生物量之比值取0.205是合理的。
4 结 论
(1) 群落是以藜蒴栲、小红栲与木荷为主要建群种,为地带性植被类型;杉木也占有重要地位,这与其中一个群落由杉木林皆伐转换而来有关;群落物种的重要值分布符合正态分布。
(2)胸径与地上生物量的曲线拟合回归方程B(kg·tree-1) = 0.128*D2.3678可获得二者较高的相关性(p=0.0001);树高与胸径关系符合H=3.712 0D0.4746(p < 0.0001)。
(3) 地上生物量的估算结果为 119.720 Mg·hm-2,比实测值高2.59%;地下生物量的估算结果为23.922±4.201 Mg·hm-2,系统总生物量为 143.642± 8.896 Mg·hm-2。
[1] Vashum Kuimi T. and Jayakumar S. Methods to Estimate Above-Ground Biomass and Carbon Stock in Natural Forests - A Review[J]. J Ecosyst Ecogr, 2012, 2(4): 116. doi:10.4172/2157-7625. 1000116.
[2] Rodger A S. The carbon cycle and global forest ecosystem[J].Water, Air and Soil Pollution, 1993, 70: 295-307.
[3] Goodale C L, Apps M J, Birdsey R A, et al. Forest carbon sinks in the northern hemisphere. Ecological Applications, 2002, 12:891–899.
[4] Ravindranath N H, Ostwald M. Methods for estimating aboveground biomass. In N. H. Ravindranath, and M. Ostwald, Carbon Inventory Methods: Handbook for greenhouse gas inventory,carbon mitigation and roundwood production projects. Springer Science + Business Media B.V.2008, 113-14.
[5] Lu D. The potential and challenge of remote sensing-based biomass estimation. Int J Remote Sens, 2006, 27: 1297-1328.
[6] Eileen H. Helmer, Michael A. Lefsky, Dar A. Roberts. Biomass accumulation rates of Amazonian secondary forest and biomass of old-growth forests from Landsat time series and the Geoscience Laser Altimeter System. Journal of Applied Remote Sensing, 2009, 3: 1-31. DOI: 10.1117/1.3082116.
[7] Brown, S., Gillespie, A.J., Lugo, A.E. Biomass estimation methods for tropical forests with applications to forest inventory data. Forest Science, 1989, 35: 881–902.
[8] Brown, S. Estimating biomass and biomass change of tropical forests. Forest Resources Assessment Publication. Forestry Papers 134. FAO, Rome. 1997. 55.
[9] Vashum K T and Jayakumar S. Methods to Estimate Above-Ground Biomass and Carbon Stock in Natural Forests - A Review[J]. Journal of Ecosystem & Ecography, 2012, 2(4): 116.doi:10.4172/2157-7625.1000116
[10] Houghton R A. Aboveground Forest Biomass and the Global Carbon Balance[J]. Global Change Biology, 2005, 11: 945–958.
[11] José Návar. Measurement and Assessment Methods of Forest Aboveground Biomass: A Literature Review and the Challenges Ahead. In Biomass, edited by Maggie Momba and Faizal Bux.2010, Sciyo, Croatia. pp. 202.
[12] 广东南岭国家级自然保护区生物多样性硏究《编辑委员会》广东省林业局, 华南农业大学, 等. 广东南岭国家级自然保护区生物多样性硏究[M]. 广东科技出版社, 2003
[13] 李 根, 周光益, 王 旭, 等. 南岭小坑藜蒴栲群落地上部分生物量分配规律[J]. 生态学报, 2011, 31(13): 3650-3658.
[14] 李 根, 周光益, 吴仲民, 等. 南岭小坑木荷群落地上生物量[J]. 林业科学, 2012, 48(3): 143-147.
[15] 李意德. 海南岛热带山地雨林林分生物量估测方法比较分析[J]. 生态学报, 1993, 13(4): 313-320.
[16] Niklas K J. Plant allometry, the scaling of form and process[[M].University of Chicago Press, Chicago. 1994. 365pp.
[17] Ketterings, Q.M., Coe, R., van Noordwijk, M., Ambagau, Y.,Palm, C.A., 2001. Reducing uncertainty in the use of allometric biomass equations for predicting aboveground tree biomass in mixed secondary forests. Forest Ecology and Management 146,199–209.
[18] IPCC (1996) Land-Use Change & Forestry. In: Revised 1996 IPCC Guidelines for National Greenhouse Gas Inventories:Reference Manual, pp. 5.1–5. Intergovernmental Panel on Climate Change Geneva, Switzerland.
[19] Huxley A. In: Mac Veagh L.(Ed.), Problems of Relative Growth.The Dial Press, New York, 1932, 276p.
[20] Komiyama A, Poungparn S, Kato S. Common allometric equations for estimating the tree weight of mangroves[J]. Journal of tropical Ecology, 2005, 21: 471-477.
[21] Chave J, Muller-Landau H C, Baker T R, et al. Regional and phylogenetic variation of wood density across 2456 Neotropical tree species[J]. Ecological Applications, 2006, 16(6): 2356–2367.
[22] Fearnside, P. M. Wood density for estimating forest biomass in Brazilian Amazonia. Forest ecology and management, 1997,90(1): 59-87.
[23] Brown I f, Martinelli L A, Thomas W W, et al. Uncertainty in the biomass of Amazonian forests: an example from Rondonia,Brazil[J]. Forest Ecology and Management, 1995, 75: 175-189.
[24] Brown S. Estimating biomass and biomass change of tropical forests, a primer[R]. FAO Forestry paper No. 134. FAO, Rome, 55.
[25] Liang Hong-wen, Huang Shou-xian, Qin Ya-li, et al. Preliminary Researches of the Wood Physical-mechanical Properties of 23-year-old Castanopsis fssa[J]. Journal of Northwest Forestry University, 2007, 22(1): 115.
[26] Cairns MA, Brown S, Helmer EH, et al. Root biomass allocation in the world’s upland forests[J]. Oecologia, 1997, 111: 1–11.
[27] Mokany K, Raison R J, Prokushkin A S. Critical analysis of root:shoot ratios in terrestrial biomes[J]. Global Change Biology,2006, 12: 84-96,doi: 10.1111/j.1365-2486.2005.001043.x
[28] 陈章和, 张宏达, 王伯荪, 等. 广东黑石顶常绿阔叶林生物量及其分配的研究[J]. 植物生态学与地植物学学报, 1993,17(4): 289-298.
[29] Voget K A, Vogt D J, Palmiotto P A, et al. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species[J]. Plant and Soil, 1996, 187: 159-219.
[30] Litton C M, Raich J W, Ryan M G. Carbon allocation in forest ecosystems[J]. Global Change Biology, 2007, 13: 2089-2109,doi: 10.1111/j.1365-2486.2007.01420.x
[31] 陈富强, 罗 勇, 李清湖. 粤东地区森林灌木层优势植物生物量估算模型[J]. 中南林业科技大学学报, 2013, 33(2): 5-10.
[32] 宁 晨, 闫文德, 叶生晶, 等. 喀斯特城市亮叶桦意杨混交林生态系统生物量及生产力研究[J]. 中南林业科技大学学报,2011, 31(5): 161-166.
Estimation of biomass in natural secondary forest ecosystem of Xiaokeng,Nanling mountain of southern China
HUANG Zhi-hong1, CHEN Bu-feng2, ZHOU Guang-yi2, WU Zhong-min2, WANG Xu2, LI Gen2, LIANG Rui-you3
(1. National Engineering Laboratory for Applied Technology of Forestry and Ecology in South China, Central-South University of Forestry & Technology, Changsha 410004, Hunan, China; 2 Research Institute of Tropical Forestry, CAF, Guangzhou 510520,Guangdong, China; 3 Xiaokeng State Forestry Farm of Shaoguan City, Qujiang 512162, Guangdong, China)
Subtropical evergreen broad-leaf forests were paid more attention for its higher biodiversity and potential in carbon sequenstration. While biomass of typical forest types in Nanling mountains in northern Guangdong province was poorly investigated.Allometric equations have been widely used to estimate biomass and carbon stock of forests. More accurate estimation of biomass is necessary for better studying carbon sequenstration in forest growth, especially, for the deforestation or reforestation areas. However,the higher accuracy in parameters estimation for allometric equations needs a larger numbers of samplings. Therefore, the allometric equations, combined with destructive samplings in the natural secondary forests of Xiaokeng, Nanling mountains of southern China,were employed to estimate biomass in the ecosystem. An extensive ground truth data base of natural secondary succession was constructed by using feld survey and detailed inventory and harvest method carried out in selected plots. The following results show that (1) the dominant tree species were Castanopsis fissa, Castanopsis carlesii, Schima superba, which were all of the regional typical vegetation types, with an exception of Cunninghamia lanceolata being one of key species due to one of communities transformed from clear-cut plantation of C. lanceolata. And the distribution of important values of the species in the sampled community obeyed to normal distribution. (2) allometric equations are presented as: B (kg·tree-1) = 0.128*D2.3678, based on the diameter at breast height and total aboveground biomass at signifcant level of p=0.000 1. At the same time, a power function of H=3.7120D0.4746(p<0.000 1) was yielded for co-relationship of tree height (H) and mean DDBH. Both of them were cooperated into a more general allometric equation as B=0.19ρD2+cto estimate aboveground biomass (BAGB) of the community. And (3) estimation of BAGBwas about 119.720 Mg·hm-2, which was a little higher than the measured BAGBby 2.59%. The belowground biomass (BBGB) was also calculated with a default value of 0.205(S.E.=0.036) presented by IPCC(1996) and estimated by 23.922±4.201 Mg·hm-2. By combination of BAGBand BBGB, the total biomass of the sampled ecosystem was about 143.642±8.896 Mg·hm-2. The research fndings suggested the natural secondary forest in this region would play an important role in carbon sequestration for the future due to its young stage at present.
natural secondary forest; aboveground biomass; belowground biomass; allometric equations; Nanling Mountains
2012-10-16
林业公益行业专项经费项目(200804006);广东省自然科学基金项目(8151200501000010);科技部国际合作项目(2007DFA31070);中南林业科技大学青年基金重点项目(2005009A)
黄志宏(1969-),男,湖北天门人,博士,副教授,主要从事生态系统生态学研究;E-mail:huanghugh2013@yahoo.com
S792
A
1673-923X(2013)08-0083-08
[本文编校:吴 彬]
猜你喜欢
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年2期)2021-07-13
农民致富之友(2020年8期)2020-05-11
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
莫愁(2017年6期)2017-02-15
莫愁·家教与成才(2017年2期)2017-02-14
绿色科技(2016年15期)2016-10-11
安徽农业科学(2015年9期)2015-01-12
西部(2011年23期)2011-08-15
