
灵石山不同海拔米槠林优势种叶片δ13C值与叶属性因子的相关性
2013-12-25 03:35:43王英姿
生态学报 2013年10期
王英姿
(福州市林业局,福州 350005)
植物组织的δ13C提供了整合分析植物固碳过程中内部生理特征和外部影响光合气体交换的环境因子的途径[1-2],稳定性碳同位素技术的使用对于了解植物养分含量和生理生态过程的相互作用提供了重要的视角[3],可以揭示与植物生理生态过程相联系的一系列气候环境信息。稳定同位素技术已成为解释植物与环境相互作用的有力工具,被广泛应用于全球变化、地球科学和植物生理生态学等研究领域[4-6]。植物是环境变化的指示器,叶片是植物对环境变化较为敏感的营养器官,其特征能体现环境因子变化的影响或植物对环境的适应。叶片δ13C值、比叶面积 (SLA)、叶片干物质含量(LDMC)、营养元素含量等叶属性特征体现了物种本身的生物学特性,是决定其在群落中地位的重要因素,这些叶片性状共同体现了植物为获取最大碳收获所采取的生存适应策略,具有重要的生态学和生物进化意义[7-8]。阔叶树种的叶片特征在不同生境之间会有很大的变化[9],在同一生境内叶片特征也会随叶片年龄的变化而变化,研究叶片特征之间的关系,找出不同性状之间的联系,然后确定形成植物生态策略某一维度的性状组合,是生态学研究中非常重要的内容[10-11]。
海拔是一个重要的地形因子,各种环境因子如气温、气压、CO2分压、光照等在海拔梯度上也表现出梯度性变化,这为研究植物生理形态对环境的响应提供了很好的条件[12]。目前较多的研究集中在运用稳定性碳同位素技术探讨单一植物的叶片δ13C特征与环境因子的关系[6,13-14],有关植物特别是优势种植物叶片δ13C的环境响应规律还缺乏系统研究[15]。
米槠(Castanopsiscarlessii)是我国东部湿润亚热带地区常绿阔叶林的重要建群种,也是世界上罕见的植被类型[16]。米槠适应性强,分布广,培肥土壤、涵养水源的能力较强,米槠林群落结构较为复杂,物种相对丰富,近年来笔者所在课题组首次对米槠林优势种叶片养分结构、叶绿素等叶属性特征沿海拔的变化进行研究并报道[17-19],表明叶属性是植物长期适应环境的结果,本文进一步对不同海拔米槠林叶片δ13C值与各叶属性因子的关系进行分析,进而探讨不同生境下植物叶片生理生态特性,以期为中亚热带相对贫瘠的森林立地上常绿阔叶植物的生态适应策略和环境响应机制以及生态系统的保护提供理论依据。
1 研究区概况
灵石山国家森林公园位于闽中福清市西部,北纬25°40′,东经119°13′,总面积2275.0 hm2,其中有林地面积2040.5 hm2,森林覆被率93.5%。属南亚热带气候带闽东南沿海海洋性季风气候区域。全年平均气温为19.7℃,7月平均气温29℃,1月平均气温7℃,极端最高气温34℃,极端最低气温0℃。年均日照时数2000h,无霜降期340—360d,有雾日20—30d。年均降水量1780mm,雨季多在3—6月,相对湿度86%。森林植被保持良好,公园内水质甚佳,清澈透明,各山涧小溪水源常年不断。土壤大多为花岗岩发育的酸性土壤,地带性土壤为红壤,其中海拔600m以上为黄红壤。公园内土壤表层物理性能良好,土层深厚,腐殖质层较厚。在《福建植被区划》上,灵石山森林公园属于闽粤沿海丘陵平原南亚热带雨林区闽江口、鹫峰山南湿暖南亚热带雨林小区。海拔250m以下为地带性植被,随着海拔的增高,森林植被依次为亚热带常绿阔叶林、山地矮林,间有马尾松林和竹林。
2 研究方法
2.1 调查与实验方法
根据灵石山国家森林公园米槠林的主要分布范围,结合GPS定位技术,采用样线和样地相结合的方法,于2008年3月在灵石山设9个不同海拔梯度,根据米槠林生长状况及地形等环境因子,按海拔从低到高分别记为A1(157m)、A2(200m)、A3(242m)、A4(332m)、A5(442m)、A6(531m)、A7(632—662m)、A8(762m)、A9(842m)。每一海拔高度视野外情况而定,选取1—2 个20m × 20m的典型样地,将各样地分成16个5m×5m的小样方,对各小样方内胸径≥2.5 cm的树种进行每木调查,实测胸径、树高、冠幅等指标;通过计算重要值确定优势种,A1—A9的所有优势种有米槠(C.carlessii)、闽桂润楠(Machilusminkweiensis)、鹅掌柴(Scheffleraheptaphylla)、木荷(Schimasuperba)、微毛柃(Euryahebeclados)、细齿柃(Enitidaacuminatissima)、罗浮栲(Castanopsisfabri)、山杜鹃(Rhododendronsimisii)、九节木(Psychotriarubra)、罗伞树(Ardisiaquinquegona)、围涎树(Pithecellobium.clypearia)、赤楠(Syzygiumbuxifolium)、青冈栎(Cyclobalanopsisglauca)、密花树(Rapaneaneriifolia)、乌药(Linderaaggregata)、油茶(Camelliaoleifera)、石栎(Lithocarpusglabra),柏拉木(Blastuscochinchinensis)、肉实树(Sarcospermalaurinum)、黑壳楠(Linderamegaphylla)、厚叶冬青(Ilexpachyphylla)、狗骨柴(Diplosporadubia)、八瓣糙果茶(Camelliaoctopetala)(表1)。然后每样地中选择各优势树种接近平均胸径的标准木3株,在树冠的中上层分东西南北四个方向采集树枝[12],按叶鳞痕等生长状况将叶片分为1年生和2年生叶片[20-21],封装后立刻带回实验室处理,SLA为叶片干重与面积的比值[22-23];Chl含量测定采用丙酮乙醇混合液法测定[24];叶片N含量采用靛酚蓝比色法测定,P含量用钼锑抗比色法测定,K含量用火焰光度计法测定,Ca、Mg、Fe、Mn、Zn含量用原子吸收分光光度计测定[25-26]。并进行SLA、Chl含量以及养分含量等叶属性的测定[22-26]。用MAT-251DELTA plus Advantage 型稳定同位素质谱仪(mass spectrometer)分析样品的碳同位素组成13C/12C。采用PDB(Pee Dee Belemnite)标准,测量误差小于0.05‰,δ13C值按国际通用标准测定形式[27]:
δ13C=[(13C/12C)sample-(13C/12C)standard]/(13C/12C)standard×1000。
2.2 数据分析
实验所得数据采用SPSS13.0软件进行相关性及回归统计分析(P<0.05)。

表1 灵石山米槠林不同海拔优势种列表
3 结果与分析
3.1 灵石山米槠林不同海拔δ13C的特征
表2总结了灵石山米槠林不同海拔优势种叶片δ13C值,优势种叶片δ13C平均值为(-30.885±1.205)‰,最高值为A1的细齿柃,为-28.806‰;最低值来自A1的围涎树,为-34.399‰。C3植物的δ13C变化范围在-35‰—-20‰之间,说明灵石山米槠林优势种采用C3植物光合代谢途径,属于C3植物。
3.2 各海拔梯度米槠林优势种不同叶龄δ13C值与各叶属性因子的相关系数
分别对灵石山不同海拔米槠林不同年龄叶片δ13C值与各属性因子的相关性进行分析(表1,表2),结果表明,不同年龄叶片δ13C值与各属性因子的关系表现不尽相同,1年生叶片δ13C值与SLA在A1、A5和A8、2年生叶片δ13C值与SLA除在A4和A6外,在其它海拔梯度均呈现显著负相关。1年生叶片δ13C值与Chl在A1、A2、A6和A9、2年生叶片δ13C值与Chl在A1、A5、A6、A7和A9呈现正相关。1年生叶片δ13C值与N含量在A1和A6呈显著负相关, 2年生叶片δ13C值与N在A1,A2,A6,A7,A8呈显著负相关。1年生叶片δ13C值与P含量在A1、A2、A3、A5和A7呈显著正相关,在A6呈显著负相关;2年生叶片δ13C值与P在A1、A2、A3和A5呈显著正相关。1年生叶片δ13C值和K含量在A7呈显著正相关,2年生叶片δ13C值和K含量在A6呈显著负相关。1年生叶片δ13C值与Na含量在A2、A3、A4和A6呈显著负相关,2年生叶片δ13C值与Na含量在A1,A2,A3,A4,A8呈显著负相关,在A7和A9呈显著正相关。1年生叶片δ13C值与Ca含量在A1、A2、A3和A6呈显著正相关,在A4和A5呈显著负相关,2年生叶片δ13C值与Ca含量除在A8相关性不显著、在A4呈负相关外,在其它海拔梯度上均呈显著正相关。不同年龄叶片在各海拔梯度上δ13C值和Mg含量的相关性均未达到显著性水平。不同年龄叶片δ13C值和Fe、Mn、Zn含量的相关性在不同的海拔梯度上表现形式比较复杂。1年生和2年生叶片δ13C值和LDMC分别在A1、A4、A6、A9和A1、A3、A6、A7、A9呈显著正相关。1年生叶片δ13C值和Ash(灰分)含量在A5、A6、A7呈显著负相关,2年生叶片δ13C值和Ash含量在A2、A5、A7二者呈显著负相关。

表2 灵石山米槠林各海拔优势种δ13C值
A1—A9代表不同的海拔梯度,At代表所有海拔
表2各海拔梯度米槠林优势种1年生叶片δ13C值与各属性因子的相关系数
Table2Relatedcoefficientsbetweenfoliarδ13Cvalueandleaftraitfactorsofone-year-oldleavesofdominantspeciesateachaltitudegradientinCastanopsiscarlessiiforestsinLingshishanNationalForestPark

海拔SLAChlNPKNaCaA1-0.665∗0.684∗-0.630∗0.732∗∗-0.623-0.680.821∗∗A2-0.0450.68∗-0.6120.804∗-0.551-0.733∗0.709∗A3-0.371-0.24-0.4320.754∗-0.21-0.706∗0.758∗A4-0.896-0.708-0.7460.871-0.15-0.936∗-0.925∗A5-0.647∗0.230.3120.5190.412-0.200-0.803∗A6-0.1750.858∗-0.865∗-0.881∗∗-0.13-0.811∗0.810∗∗A70.914∗-0.644-0.3320.986∗∗0.990∗∗0.3750.620A8-0.957-0.7011-0.2180.944-0.911-0.2540.586A9-0.943∗0.891-0.378-0.8110.860-0.2310.723海拔MgFeMnZnLDMCAshA1-0.090.5260.884∗∗-0.745∗0.713∗-0.078A2-0.390.957∗0.010.842∗0.054-0.785A30.40-0.687∗0.5250.6390.1930.864∗A4-0.41-0.914∗0.960∗0.720.994∗-0.763A5-0.31-0.787∗-0.698∗-0.766∗∗-0.526-0.710∗A6-0.040.900∗∗0.904∗∗-0.786∗0.781∗-0.861∗A7NS-0.848∗-0.3270.963∗∗0.655-0.889∗A8NS-0.5000.867-0.971∗0.952-0.575A9NS-0.262-0.349-0.931∗0.953∗-0.471
表3各海拔梯度米槠林优势种2年生叶片δ13C值与各属性因子的相关系数
Table3Relatedcoefficientbetweenfoliarδ13Cvalueandleaftraitfactorsoftwo-year-oldleavesofdominantspeciesateachaltitudegradientinCastanopsiscarlessiiforestsinLingshishanNationalForestPark
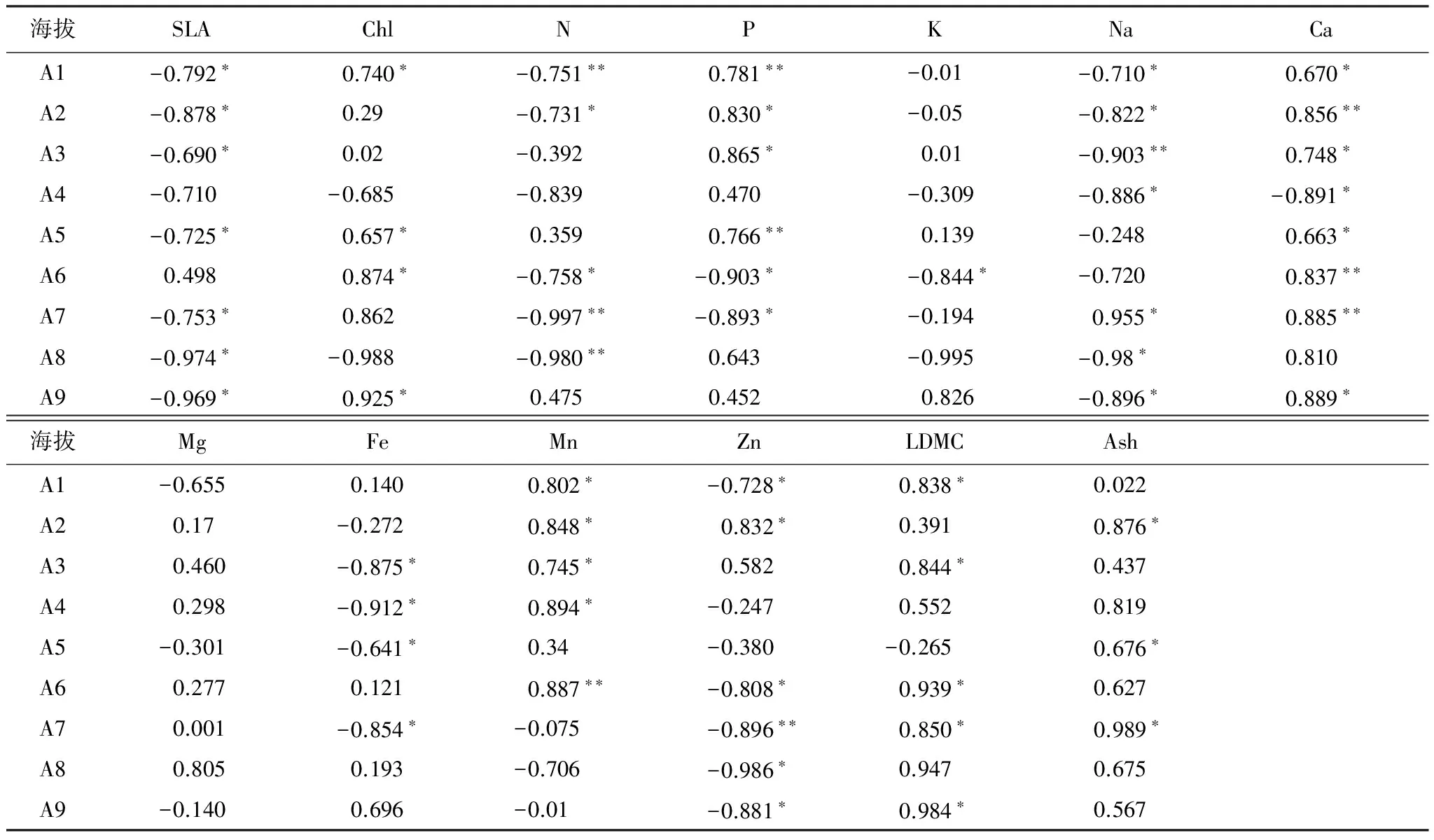
海拔SLAChlNPKNaCaA1-0.792∗0.740∗-0.751∗∗0.781∗∗-0.01-0.710∗0.670∗A2-0.878∗0.29-0.731∗0.830∗-0.05-0.822∗0.856∗∗A3-0.690∗0.02-0.3920.865∗0.01-0.903∗∗0.748∗A4-0.710-0.685-0.8390.470-0.309-0.886∗-0.891∗A5-0.725∗0.657∗0.3590.766∗∗0.139-0.2480.663∗A60.4980.874∗-0.758∗-0.903∗-0.844∗-0.7200.837∗∗A7-0.753∗0.862-0.997∗∗-0.893∗-0.1940.955∗0.885∗∗A8-0.974∗-0.988-0.980∗∗0.643-0.995-0.98∗0.810A9-0.969∗0.925∗0.4750.4520.826-0.896∗0.889∗海拔MgFeMnZnLDMCAshA1-0.6550.1400.802∗-0.728∗0.838∗0.022A20.17-0.2720.848∗0.832∗0.3910.876∗A30.460-0.875∗0.745∗0.5820.844∗0.437A40.298-0.912∗0.894∗-0.2470.5520.819A5-0.301-0.641∗0.34-0.380-0.2650.676∗A60.2770.1210.887∗∗-0.808∗0.939∗0.627A70.001-0.854∗-0.075-0.896∗∗0.850∗0.989∗A80.8050.193-0.706-0.986∗0.9470.675A9-0.1400.696-0.01-0.881∗0.984∗0.567
3.3 灵石山米槠林优势种叶片δ13C值与各属性因子的回归分析
对灵石山米槠林所有海拔梯度上优势种不同年龄叶片δ13C值与各属性因子的相关性进行回归分析(图1),结果表明不同叶龄叶片δ13C值与SLA呈线性负相关;2年生叶片δ13C值与Chl呈现正相关,1年生叶片δ13C值与Chl相关性不显著;不同叶龄叶片δ13C值与N含量呈线性负相关;不同叶龄叶片δ13C值与P呈线性正相关;1年生叶片δ13C值和K含量相关性不显著,2年生叶片呈显著负相关;不同年龄叶片δ13C值和Na含量均呈显著线形负相关;不同年龄叶片δ13C值和Ca含量均呈线性正相关;不同叶龄叶片δ13C值和Mg含量均呈线性负相关;不同年龄叶片δ13C值和Fe含量没有呈现显著的相关关系;不同年龄叶片δ13C值和Zn含量没有呈现显著的相关关系;不同叶龄叶片δ13C值和LDMC呈线性正相关;不同叶龄叶片δ13C值和Ash含量呈负线性相关。
4 讨论
4.1 灵石山米槠林优势种叶片δ13C与SLA的关系
SLA是植物叶片长期对生长环境的一种适应,是一个衡量植物种相对生长速率的重要参数,是叶片厚度和组织组成的综合反映[28],植物的很多特征都直接或间接与SLA有关[29]。本研究中,灵石山米槠林优势种叶片δ13C与SLA呈负相关。Luo对中国东南部不同海拔云杉种群的研究及Ares等对美国夏威夷柳叶桉和红椿中也得出了一致的结果[3,30];王玉涛和薛慧勤等的研究得出了一致的结论,并指出在实际应用中可以使用SLA代替碳同位素比率来估测水分利用效率[31-32]。叶片δ13C与SLA呈负相关说明植物通过降低叶片面积来提高水分利用效率。
4.2 灵石山米槠林优势种叶片δ13C与Chl以及养分含量的关系
Chl是是植物吸收太阳能进行光合作用的重要物质,起到接受和转换能量的作用。目前,有关叶片δ13C值与Chl关系的研究报道不多。本研究中,2年生叶片δ13C与Chl含量显著负相关。与本研究结果不同,马剑英等对红砂叶片δ13C值与Chl含量的分析表明二者没有显著相关性[33]。Bort 等对大麦的研究表明叶片δ13C和基于面积的SPAD值没有相关性[34]。
表4灵石山米槠林优势种不同年龄叶片稳定性碳同位素和叶片属性的回归关系
Table4Linearregressionbetweenfoliarδ13CvalueandleaftraitfactorsofdifferentageleavesofdominantspeciesinCastanopsiscarlessiiforestsinLingshishanNationalForestPark

1年生Oneyear回归关系式R22年生Twoyear回归关系式R2SLA=-22.702X-540.850.617∗∗SLA=-22.863X-559.360.651∗∗Chl=0.0202X+2.07460.063Chl=0.1126X+5.020.388∗∗N=-2.4724X-61.5090.447∗∗N=-2.7855X-71.8320.487∗∗P=0.0998X+3.94040.520∗∗P=0.0666X+2.81160.395∗∗K=-0.5463X-9.03760.162K=-2.8641X-78.9080.525∗∗Na=-0.1401X-3.82460.553∗∗Na=-0.1332X-3.66540.564∗∗Ca=4.3692X+166.40.435∗∗Ca=4.4364X+168.850.448∗∗Mg=-0.1511X-1.51320.322∗Mg=-0.1506X-1.59190.325∗Fe=-0.7257X+72.0360.035Fe=-1.3462X+59.3220.056Mn=-0.1596X-2.57370.305∗Mn=0.1878X+8.13660.364∗∗Zn=-4.3471X-80.7940.232Zn=-3.5105X-50.7790.138LDMC=0.0352X+1.34710.692∗∗LDMC=0.0383X+1.47580.603∗∗Ash=-8.7623X-216.640.435∗∗Ash=-7.725X-184.070.409∗∗
X代表叶片δ13C,回归分析的样本数n=66
叶片N含量在一定程度上反映了叶片吸收和固定大气CO2的能力,进而影响δ13C值[35],本研究中叶片δ13C值与N含量呈显著负相关,与李善家、Vitousek和Hultine等研究结果一致[15,36-37],也有研究表明植物叶片中的N含量与δ13C值呈正相关关系[33,38],但是较多的研究发现植物叶片中的δ13C值与叶片N含量之间没有相关性[30,39-42 ]。叶片δ13C值与N含量的相关性,除了与叶片的生理结构有关,还与叶N在光合器官和非光合器官的分配、以及N在光合器官内部各组分之间的分配比例以及N素光合利用效率等因素有关[43-46]。
P在植物新陈代谢过程中起重要作用,不但通过影响植物的渗透调节能力和保水力来增强植物组织的抗旱能力,而且能够通过提高植物体内可溶性糖和磷脂的含量增强植物的抗寒性。本研究中,叶片δ13C与P含量呈正相关,这与前人的研究结果一致[33,42]。中国植物普遍缺P,特别在南方热带亚热带地区表现更为明显[47],因此P作为生理元素,对植物的光合作用以及同位素判别变得更加重要。
K在调节气孔功能和提高植物水分利用效率方面有着重要的作用,同时能够促进植物光合作用和光合产物的运输、提高CO2的同化率[48];Na属于“功能性营养元素”[49],Na离子可以替代K离子进行细胞渗透调节的生理功能[50-51]。本研究中,2年生叶片δ13C与K含量显著负相关,不同叶龄叶片δ13C与Na含量负相关,说明K、Na可以通过调节叶片胞间CO2浓度,使植物达到合适的水分利用效率。
Ca作为结构性元素与叶片δ13C呈显著正相关,说明Ca对细胞的渗透调节十分重要。Mg与δ13C值呈显著负相关。微量元素Fe、Mn、Zn对于植物的光合作用起着重要作用,但是各海拔梯度上叶片δ13C值和Fe、Mn和Zn含量的相关性表现形式比较复杂,这可能与环境因素以及各叶属性因子对δ13C值的影响机理有关,还有待于进一步研究。
由于LDMC在重复测定中较为稳定,很少受叶片厚度的影响,这使它成为一些学者预测植物资源利用策略的一个重要指标。本研究中,不同年龄叶δ13C值与LDMC显著正相关,这与前人的研究结果基本一致[34,52]。本研究中叶片δ13C值和Ash的负相关性与前人的研究结果一致[53-54]。
本研究选择不同海拔梯度米槠林优势种不同叶龄叶片,因此叶片δ13C与叶属性的关系更能突出了植物叶片生理关系的内在属性。总体来看,植物叶片δ13C值及分馏结果受环境因子以及养分元素等叶片属性的影响。不同年龄叶片δ13C与叶属性的关系表现形式也不同,说明植物生长过程中资源利用策略的变化。
Refenerces:
[1]Li C,Berninger F,Koskela J,Sonninen E.Drought responses of Eucalyptus microtheca provenances depend on seasonality of rainfall in their place of origin.Australian Journal of Plant Physiology,2000,27(3): 231-238.
[2]Ponton S,Dupouey J L,Breda N,Dreyer E.Comparison of water-use efficiency of seedlings from two sympatric oak species: genotype × environment interactions.Tree Physiology,2002,22(6): 413-422.
[3]Luo J X,Zang R G,Li C Y.Physiological and morphological variations ofPiceaasperatapopulations originating from different altitudes in the mountains of southwestern China.Forest Ecology and Management,2006,221(1/3): 85-290.
[4]He C X,Li J Y,Meng P,Zhang Y X.Changes in leaf stable carbon isotope fractionation of trees across climatic Gradients.Acta Ecologica Sinica,2010,30(14): 3828-3838.
[5]Lin G H.Stable isotope ecology: a new branch of ecology resulted from technology advances.Chinese Journal of Plant Ecology,2010,34(2): 119-122.
[6]Feng Q H,Cheng R M,Shi Z M,Liu S R,Liu X L,He F,Cao H M.Response of foliar δ13C ofQuercusspinosato altitudinal gradients.Acta Ecologica Sinica,2011,31(13): 3629-3637.
[7]Cordell S,Goldstein G,Meinzer F C,Vitousek P M.Regulation of leaf life-span and nutrient-use efficiency ofMetrosideropolymorphatrees at two extremes of a long chronosequence in Hawaii.Oecologia,2001,127: 198-206.
[8]Wright I J,Reich P B,Westoby M.The worldwide leaf economics spectrum.Nature,2004,428: 821-827.
[9]Liu F D,Wang Z S,Zhang M,Wang W J,An S Q,Zheng J W,Yang W J,Zhang S T.Photosynthesis in relation to leaf nitrogen,phosphorus and specific leaf area of seedlings and saplings in tropical montane rain forest of Hainan Island,South China.Acta Ecologica Sinica,2007,27(11): 4651-4661.
[10]Shipley B,Lechowicz M J.The functional co-ordination of leaf morphology,nitrogen concentration,and gas exchange in 40 wetland species.Ecoscience,2000,7(2): 183-194.
[11]Westoby M,Wright I J.Land-plant ecology on the basis of functional traits.Trends in Ecology and Evolution,2006,21(5): 261-268.
[12]Qi J,Ma K M,Zhang Y X.The altitudinal variation of leaf traits ofQuercusliaotungensisand associated environmental explanations.Acta Ecologica Sinica,2007,27(3): 930-937.
[13]Lin L,Chen L T,Zheng W L.Altitudinal Variation of the Foliar δ13C inAbiesgeorgeivar.smithiiandQuercusaquifolioidesin Tibet.Journal of Glaciology and Geocryology,2008,30(6): 1048-1054.
[14]Zhang P,Wang G,Zhang T,Chen N L.Responses of foliar δ13C inSabinaprzewalskiiandPiceacrassifoliato altitude and its mechanism in the Qilian Mountains,China.Chinese Journal of Plant Ecology,2010,34 (2): 125-133.
[15]Li S J,Zhang Y F,Chen T.Relationships between foliar stable carbon isotope composition and environmental factors and leaf element contents ofPinustabulaeformisin northwestern China.Chinese Journal of Plant Ecology,2011,35(6): 596-604.
[16]Yan S J,Hong W,Wu C Z,Bi X L,Lan B.The structure and distribution pattern of dominant populations inCastanopsiscarlesiiCommunity.Journal of Tropical and Subtropical Botany,2002,10(1): 15-21.
[17]Wang Y Z,Hong W,Wu C Z,Zheng G G,Fan H L,Chen C,Li J.Research on leaf traits for different age leaves of dominant species inCastanopsiscarlessiiforests in Lingshishan National Forest Park.Journal of Fujian College of Forestry,2009,29(3): 203-209.
[18]Wang Y Z,Hong W,Wu C Z,Lin H,Fan H L,Chen C,Li J.Research on the variation of uneven-aged leaf SPAD values and the correlation between SPAD and chlorophyll,nitrogen concentration of Dominant Species inCastanopsiscarlessiiforsests in Lingshihan National Forest Park.Journal of Forestry Research,2009,20(4): 362-366.
[19]Wang Y Z,Hong W,Wu C Z,Zheng G G,Fan H L,Chen C,Li J.Variation of chloroplast pigments of various age leaves along elevation gradients of dominant species inCastanopsiscarlessiiForests in Lingshishan National Forest Park.Scientia Silvae Sinicae,2010,46(11): 43-51.
[20]Cornelissen J H C,Lavorel S,Garnier E.A handbook of protocols for standardised and easy measurement of plant functional traits worldwide.Australian Journal of of Botany,2003,51(4): 335-380.
[21]Reich P B,Uhl C,Walters M B,Prugh L,S.Ellsworth A D.Leaf demography and phenology in Amazonian rain forest: a census of 40,000 leaves of 23 tree species.Ecological Monographs,2004,74(1): 3-23
[22]Yu S L,Ma K P,Chen L Z.Analysis on leaf forms inQuercusmongolicacommunity.Chinese Journal of Applied Ecology,2003,14(1): 151-153.
[23]Garnier E,Shipley B,Roumet C,Laurent G.A Standardized protocol for the determination of specific leaf area and leaf dry matter content.Functional Ecology,2001,15(5): 688-695.
[24]Zhang X Z.Determination of plant chlorophyll content by a mixture with Acetone and Ethanol.Liaoning Agricultural Sciences,1986,(3): 26-28.
[25]Li Y H,Luo T X,Lu Q,Tian X Y,Wu B,Yang H H.Comparisons of leaf traits among 17 major plant species in shazhuyu Sand Control Experimental Station of Qinghai Province.Acta Ecologica Sinica,2005,25(5): 994-999.
[26]LY/T 1210-1275—1999.Republic of China Forestry Industry Standard.Beijing: State Forestry Administration,P.R.China,1999: 274-298.
[27]Farquhar G D,Ehleringer J R,Hubick k T.Carbon isotope discrimination and photosynthesis.Annual Review of Plant Physiology and Plant Molecular Biology,1989,40: 530-536.
[28]Garnier E,Cordonnier P,Guillerm J L,Sonié L.Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in Mediterranean old-fields.Oecologia,1997,111(4): 490-498.
[29]Wright I J,Reich P B,Westoby M.Strategy shifts in leaf physiology,structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats.Functional Ecology,2001,15: 423-434.
[30]Ares A,Fownes J H.Productivity,nutrient and water-use efficiency ofEucalyptussalignaandToonaciliatain Hawaii.Forest Ecology and Management,2000,139(1/3): 227-236.
[31]Wang Y T, Li J Y,Cheng W,Cheng W,Geng X.Study on seasonal variation in carbon isotope composition correspond with soil temperature and humidity and weather factors for main greening tree species in Beijing.Acta Ecologica Sinica,2008,28(7): 3143-3151.
[32]Xue H Q,Gan X M,Sun M H,Sun L Z.Study on the relationship between water use efficiency and carbon isotope discrimination in drought condition.Chinese Journal of Oil Crop Science,1999,21 (1): 27-34.
[33]Ma J Y,Chen F H,Xia D S,Sun H L,Duan Z H,Wang G.Correlations between leaf δ13C and physiological parameters of desert plant Reaumuria soongorica.Chinese Journal of Applied Ecology,2008,19 (5): 1166-1171.
[34]Bort J,Araus J L,Hazzam H,Grando S,Ceccarelli S.Relationships between early vigour,grain yield,leaf structure and stable isotope composition in field grown barley.Plant Physiology and Biochemisty,1998,36(12): 889-897.
[35]Zhao C M,Chen L T,Ma F,Yao BQ,Liu J Q.Altitudinal differences in the leaf fitness of juvenile and mature alpine spruce trees (Piceacrassifolia).Tree Physiology,2008,28(1): 133-141.
[36]Hultine K R,Marshall J D.Altitude trends in conifer leaf morphology and stable carbon isotope composition.Oecologia,2000,123(1): 32-40.
[37]Vitousek P M,Field C B,Matson P A.Variation in foliar δ13C in Hawaiian Metrosiderospolymorpha: a case of internal resistance.Oecologia,1990,84(3): 362 -370.
[38]Hamerlynck E P,Huxman T E,McAuliffe J R,Smith S D.Carbon isotope discrimination and foliar nutrient status ofLarreatridentata(creosote bush) in contrasting Mojave Desert soils.Oecologia,2004,138(2): 210-215.
[39]Ares A,Fownes J H.Water supply regulates structure,productivity,and water use efficiency ofAcaciakoain Hawaii.Oecologia,1999,121 (4): 458-466.
[40]Kao W Y,Chang K W.Altitudinal trends in photosynthetic rate and leaf characteristics of Miscanthus populations from central Taiwan.Australian Journal of Botany,2001,49(4): 509-514.
[41]Kao W Y,Tsai H C,Tsai T T.Effect of Nacl and nitrogen availability on growth and photosynthesis of seedlings of a mangrove species,Kandeliacandel(L.) Druce.Journal of Plant Physiology,2001,158(7): 841-846.
[42]DesRochers A,VandenDriessche R,Thomas B R.NPK fertilization at planting of three hybrid poplar clones in the Boreal region of Alberta.Forest Ecology and Management,2006,232(1/3): 216-225.
[43]Takashima T,Hikosake K,Hirose T.Photosynthesis or persistence:nitrogen all location in leaves of evergreen and deciduous Quercus species.Plant Cell and Environment,2004,27(8): 1047-1054.
[44]Warren C R,Adams M A.Evergreen trees do not maximize instantaneous photosynthesis.Trends in Plant Science,2004,9(6): 270-274.
[45]Welker J M,Jónsdóttir I S,Fahnestock J T.Leaf isotopic (δ13C and δ13N) and nitrogen contents of Carex plants along the Eurasian Coastal Arctic:Results from the Northeast Passage expedition.Polar Biology,2003,27(1): 29-37.
[46]Ripley B S,Muller E,Behenna M,Whittington-Jones G M,Hill M P.Biomass and photosynthetic productivity of water hyacinth (Eichhorniacrassipes) as affected by nutrient supply and mirid (Eccritotaruscatarinensis) biocontrol.Biological control,2006,39(3): 392-400.
[47]Ren S J,Yu G R,Tao B,Wang S Q.Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC.Environmental Science,2007,28(12): 2665-2673.
[48]Liu X N,Ma J Y,Sun W,Cui Y Q,Duan Z H.Advances in mechanisms underlying the responses of δ13C in alpine plants to the altitudinal gradients.Journal of Mountain Science,2010,28(1): 37-46.
[49]Li S X,Zhou X R,Wang S M.Positive Functions of Sodium in Plants.Journal of Desert Research,2008,28(3): 485-490.
[50]Very A A,Robinson M F,Mansfield T A,Sanders D.Guard cell cation channels are involved in Na+induced stomatal closure in a halophyte.The Plant Journal,1998,14(5): 509 -521.
[51]Raghavendra A M,Rao J M,Das V S R.Replaceability of potassium by sodium for stomatal opening in epidermal strips of stomatal opening in epidermal strips of Commelina benghalensis.Z.Pflanzenphysiol,1976,80: 36 -42.
[52]Araus J L,Amaro T,Zuhair Y,Nachit M M.Effect of leaf structure and water status on carbon isotope discrimination in field grown durum wheat.Plant Cell and Environment,1997,20(12): 1484-1494.
[53]Tsialtas J T,Tokatlidis I S,Tsikrikoni C,Lithourgidis A S.Leaf carbon isotope discrimination,ash content and K relationships with seed cotton yield and lint quality in lines ofGossypiumhirsutumL..Field Crops Research,2007,107(1): 70-77.
[54]Araus J L,Casadesus J,Asbatia,Nachit M M.Basis of the relationship between ash content in the flag leaf and carbon isotope discrimination in kernels of durum wheat.Photosynthetica,2001,39(4): 591-596.
[4]何春霞,李吉跃,孟平,张燕香.树木叶片稳定碳同位素分馏对环境梯度的响应.生态学报,2010,30(14):3828-3838.
[5]林光辉.稳定同位素生态学:先进技术推动的生态学新分支.植物生态学报, 2010,34(2):119-122.
[6]冯秋红,程瑞梅,史作民,刘世荣,刘兴良,何飞,曹慧明.巴郎山刺叶高山栎叶片δ13C对海拔高度的响应.生态学报,2011,31(13):3629-3637.
[12]祁 建,马克明,张育新.辽东栎(Quercusliaotungensis)叶特征沿海拔梯度的变化及其环境解释.生态学报,2007,27(3):930-937.
[13]林玲,陈立同,郑伟列.西藏急尖长苞冷杉与川滇高山栎叶片δ13C沿海拔梯度的变化.冰川冻土,2008,30(6):1048-1054.
[14]张 鹏,王 刚, 张 涛,陈年来.祁连山两种优势乔木叶片δ13C的海拔响应及其机理.植物生态学报,2010, 34 (2): 125-133.
[15]李善家,张有福,陈拓.西北油松叶片δ13C特征与环境因子和叶片矿质元素的关系.植物生态学报, 2011,35(6):596-604.
[17]王英姿,洪伟,吴承祯,郑关关,范海兰,陈灿,李键.灵石山米槠林优势种群不同叶龄叶属性的研究.福建林学院学报,2009,29(3):203-209.
[19]王英姿,洪伟,吴承祯,郑关关,范海兰,陈灿,李键.灵石山米槠林优势种不同叶龄叶绿体色素沿海拔梯度的变化.林业科学,2010,46(11):43-51.
[22]于顺利,马克平,陈灵芝.蒙古栎群落叶型的分析.应用生态学报,2003,14(1):151-153.
[24]张宪政.植物叶绿素含量测定——丙酮乙醇混合液法.辽宁农业科学,1986,(3):26-28.
[25]李永华,罗天祥,卢琦,田晓娅,吴波,杨恒华.青海省沙珠玉治沙站17种主要植物叶性因子的比较.生态学报,2005,25(5):994-999.
[26]LY/T 1210-1275—1999.中华人民共和国林业行业标准.北京:国家林业局,1999:274-298.
[31]王玉涛,李吉跃,程炜,陈崇,耿欣.北京城市绿化树种叶片碳同位素组成的季节变化及与土壤温湿度和气象因子的关系.生态学报,2008,28(7):3143-3151.
[32]薛慧勤,甘信民,孙明辉,孙兰珍.干旱条件下花生水分利用效率与叶片碳同位素辨别力的相关性研究.中国油料作物学报,1999,21 (1):27-34.
[33]马剑英,陈发虎, 夏敦胜,孙惠玲,段争虎,王刚.荒漠植物红砂叶片δ13C值与生理指标的关系.应用生态学报,2008,19(5):1166-1171.
[47]任书杰,于贵瑞,陶波,王绍强.中国东部南北样带 654 种植物叶片氮和磷的化学计量学特征研究.环境科学,2007,28(12):2665-2673.
[48]刘小宁,马剑英,孙伟,崔永琴,段争虎.高山植物稳定碳同位素沿海拔梯度响应机制的研究进展.山地学报,2010,28(1):37-46.
[49]李三相,周向睿,王锁民.Na+在植物中的有益作用.中国沙漠,2008,28(3):485-490.
猜你喜欢
散文诗(2021年22期)2022-01-12 06:14:04
天津医科大学学报(2021年1期)2021-12-05 11:11:05
环球时报(2019-05-23)2019-05-23 06:17:36
音乐教育与创作(2019年9期)2019-05-16 09:34:08
音乐教育与创作(2019年8期)2019-05-16 04:06:28
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
小学生导刊(2017年17期)2017-05-17 05:44:28
宝藏(2017年2期)2017-03-20 13:16:43
中国医学影像技术(2017年11期)2017-01-16 12:54:05
考试周刊(2016年63期)2016-08-15 14:33:26
