
中小尺度下西北太平洋柔鱼资源丰度的空间变异
2013-12-21 04:59杨铭霞陈新军冯永玖官文江
生态学报 2013年20期
杨铭霞,陈新军,2,3,* ,冯永玖,2,3,官文江,2,3
(1.上海海洋大学海洋科学学院,上海 201306;2.国家远洋渔业工程技术研究中心,上海 201306;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306)
柔鱼(Ommastrephes bartramii)是西北太平洋海域重要经济头足类种类之一,目前主要由中国(包括台湾省)、日本等国家和地区所开发利用。传统鱿钓作业渔场主要分布在150°—160°E、38°—48°N海域,渔汛时间为6—12月,盛渔期为8—10月[1]。柔鱼渔场形成受到西北太平洋黑潮和亲潮的影响,因此其资源丰度的时空分布应受到一年四季周期性变化的海洋环境的影响。海洋动物分布在某种特定的空间或时间上,它们具有高度的空间和时间异质性[2]。生物的资源丰度变化及其空间异质性是景观生态学研究的核心问题[3-4],通常以地统计学中变异函数为手段,研究那些在空间分布上既具有结构性又有随机性,或有空间相关和依赖性的自然现象[5-7],这一方法已在海洋渔业领域得到了较为广泛的应用[8-14],但是多数海洋生物资源丰度的研究中,其空间分辨率多以经纬度0.5°×0.5°或1°×1°为基础的生产统计数据进行分析。由于不同种类其栖息环境不一样,影响资源分布的海洋环境因子也有显著差异,比如在西北太平洋海域,由于黑潮和亲潮2支海流的激烈交汇,使得表层水温等温线高度密集且弯曲;而在170°E以东海域的北太平洋,由于只有受到黑潮续流的北太平洋海流影响,其表层水温等温线基本上与纬度平行[1],因此,在传统鱿钓作业渔场(150°—160°E、38°—48°N)中,用于柔鱼资源丰度研究的最适空间分辨率是多少,目前未见有研究的报道。为此,本文应用地统计学方法对中小尺度下西北太平洋柔鱼资源丰度空间变异情况进行分析,探讨获得最佳的空间尺度,同时结合柔鱼洄游分布以及海流对柔鱼资源丰度分布的影响,进一步阐明其机理,为西北太平洋柔鱼资源的生产统计和科学管理提供参考。
1 材料和方法
1.1 研究数据
本研究采用2011年8—10月我国鱿钓船在西北太平洋海域生产的统计数据,包括作业时间、经度、纬度、单船产量等。单位捕捞努力量渔获量(CPUE)为一艘船一天的产量,并作为资源丰度的指标。以传统鱿钓作业渔场(150°-160°E、38°-48°N海域)作为研究对象,对各月CPUE数据进行7个空间尺度的划分,分别为经纬度 10'×10',经纬度 20'×20',经纬度 30'×30',经纬度 40'×40',经纬度 50'×50',经纬度 60'×60',经纬度 70'×70'。将经纬度 10'×10'、20'×20'、30'×30'定义为小尺度,将 40'×40'、50'×50'、60'×60'、70'×70'定义为中尺度。
1.2 研究方法
1.2.1 K-S 检验法
校验数据的正态分布性是使用地统计学和Kriging方法进行空间分析的前提[1]。为此,本文采用K-S(Kolmogorov-Smirnov)检验法,并运用SPSS17.0统计软件对各月各尺度CPUE单一样本进行正态分布检验。对某些尺度下CPUE不符合正态分布的数据,对其进行Box_Cox转换,以便达到地统计学分析的要求。单样本K-S检验方法如下:
假设H0为样本所来自的总体分布服从正态分布,H1所来自的总体分布不服从正态分布。F0(x)表示理论分布的分布函数,Fn(x)表示一组随机样本的累计频率函数。设D为F0(x)与Fn(x)差距的最大值,定义:
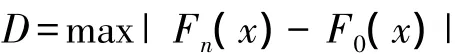
标准显著性水平α设置为0.05,若显著性概率P(Sig.(2-tailed))>0.05,表明接受假设H0,样本服从正态分布,反之则不服从。
1.2.1 地统计方法
借助ArcGis9.0地统计模块中Kriging方法对数据进行分析。Kriging插值是目前地统计学中应用最为广泛的内插方法,该方法能最大限度地利用各种信息对区域化变量做出线性无偏最优估计[2]。其基本原理和计算方法见文献[2-5]。通过地统计方法对西北太平洋柔鱼CPUE数据进行分析,建立变异函数理论模型(主要为球状模型、指数模型和高斯模型),并通过地统计学中用来描述研究对象空间分布结构的主要参数块金值、基台值、偏基台值、变程和结构比等参数来分析柔鱼资源丰度的空间变异规律。Kriging算法可表示为:
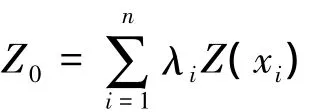
式中,Z0为采样区域内任意一点的特征值估值结果;λi为通过半方差函数分析得到的样本点xi特征值的权重。
主要参数表达的意义为:1)块金值,用C0表达,表示随机变异引起的空间异质性程度;2)偏基台值,用C表达,表示变量由自身结构性引起的空间异质性程度;3)基台值,用C0+C表达,表示区域化变量的最大变异,是半变异函数达到的极限值,即表示总空间异质性程度;4)变程,用A表达,反映了变量空间自相关范围的大小,它与观测尺度以及在取样尺度上影响柔鱼资源丰度的各种生态过程相互作用有关[15];5)变异值,用C/(C0+C)来表达,反映的是自相关部分的空间异质性占总空间异质性的程度,一般认为,其值若<25%时,表示变量的空间自相关较弱;其值>25%且<75%时,表示变量的空间自相关性处于中等;其值>75%时,则表示变量有很强的空间自相关性[2]。
2 结果
2.1 CPUE数据描述性统计与正态分布检验
数据统计显示,总体平均CPUE为2t/d左右,最高CPUE为5t/d。在7个不同的尺度下分别统计CPUE最小值、最大值和均值,发现它们都有所差异,其中每一尺度的最小值基本呈现出随空间尺度变大而逐渐递增的趋势(表1)。
K-S检验表明(表1),除8月在10'×10'、40'×40'和50'×50'这3个尺度下不符合检验条件外,其他数据均达到了地统计分析的要求。因此,对8月这3个尺度的CPUE数据进行对数、平方根、反正弦平方根、倒数及Box-Cox等方法的转换(表2),结果发现平方根、反正弦平方根和Box-Cox转换后均达到了地统计分析的要求。为了与ArcGIS9.3的地统计模块匹配,拟采用Box-Cox转换方法获得的数据(表2)。计算得知,以上3个尺度下 Box-Cox 转换的 λ 值分别为 0.687,0.612 和 0.541。
2.2 CPUE时空变异特征
(1)变异程度比较
由图1可知,8—10月各月CPUE空间变异程度均存在较大的差异。在各向同性条件下(图1),在球状模型、指数模型和高斯模型3种理论模型中,总体上看以指数模型下的CPUE空间自相关异质性程度较强,球状模型次之,高斯模型最弱。8月CPUE的空间自相关性在小尺度情况下,其变异值基本稳定;在中尺度情况下,其变异值随尺度的增加而减少,在尺度60'×60'时,其变异值几乎下降到0,CPUE的空间自相关较弱;但是,尺度70'×70'时出现一个异常的变异值,为各尺度变异值的最高,达到中等及其以上的空间自相关程度。分析发现,指数模型的空间自相关程度为最高,其变异值在不同尺度下都是最高。9月和10月3个模型CPUE空间自相关程度均为中等(变异值为25%—75%间)的出现在尺度30'×30'时,而在小尺度的其它尺度下,3个模型的变异值变化不一,均表现为弱空间自相关和中等空间自相关。9月和10月在空间尺度50'×50'、60'×60'、70'×70'情况下,其变异值均在10%以下,表现为弱的弱空间自相关程度。
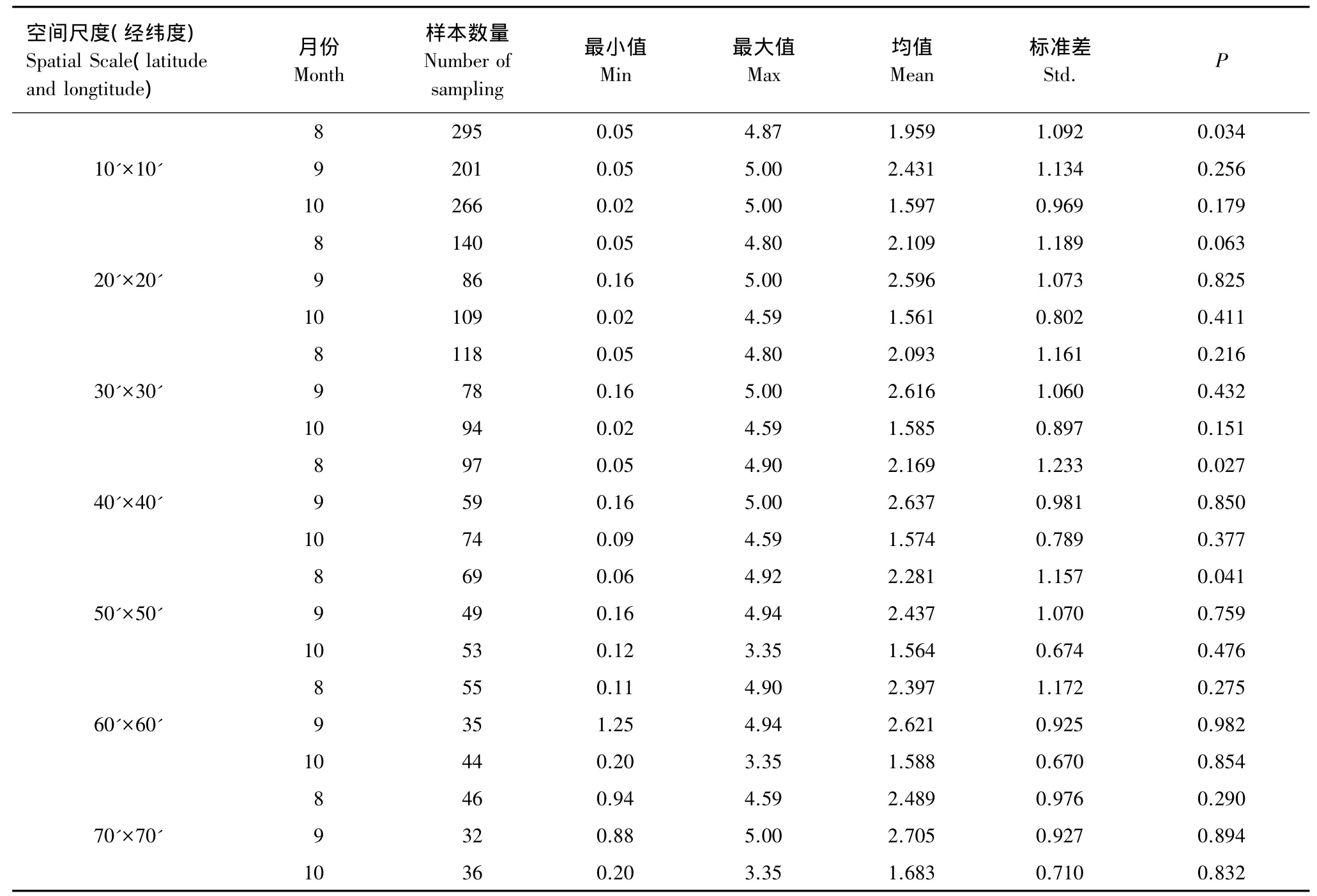
表1 不同空间尺度下的统计量及K-S检验值Table 1 The statistics data under different spatial scale and the K-Stest value
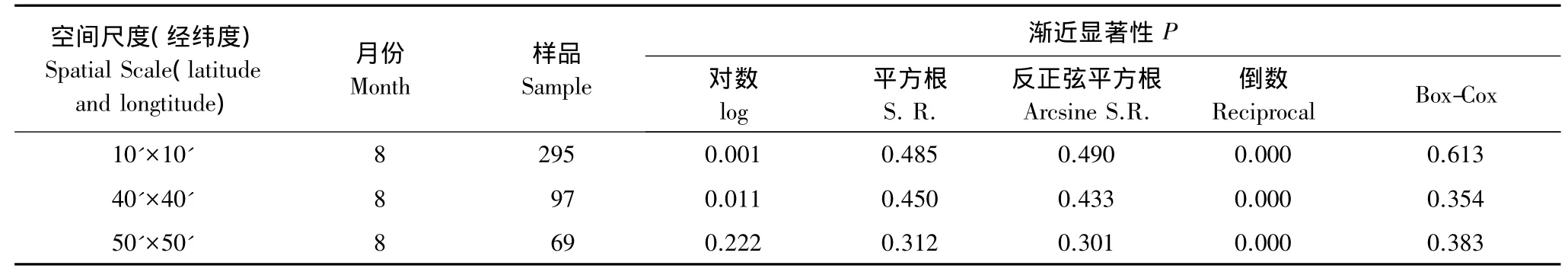
表2 数据转换后的K-S检验值Table 2 K-S test value after data conversion
在各向异性条件下(图1),8—10月3种模型的总体空间自相关程度变化趋势与各向同性基本一致。8月小尺度下3种模型CPUE空间自相关程度均在中等及其以上,其中指数模型在尺度20'×20'下出现了强的空间自相关性,而在中尺度下,3个模型在40'×40'和70'×70'尺度下,CPUE空间自相关程度均在中等及其以上,而其它2个尺度CPUE基本上表现为弱空间自相关程度。9月小尺度下,高斯模型的CPUE变异值基本上在25%左右,表现为弱的空间自相关性,球状模型和指数模型的CPUE变异值均超过25%,表温为中等的空间自相关性;而在中尺度下,高斯模型和球状模型的CPUE变异值均在25%以下,表现为弱的空间自相关性,指数模型除了70'×70'尺度外,其变异值都在25%—50%间,表现为中等的空间自相关性。10月在3个模型的拟合下,其CPUE空间自相关的变异情况趋势与各向同性时大致相同(图1)。
(2)理论模型比较
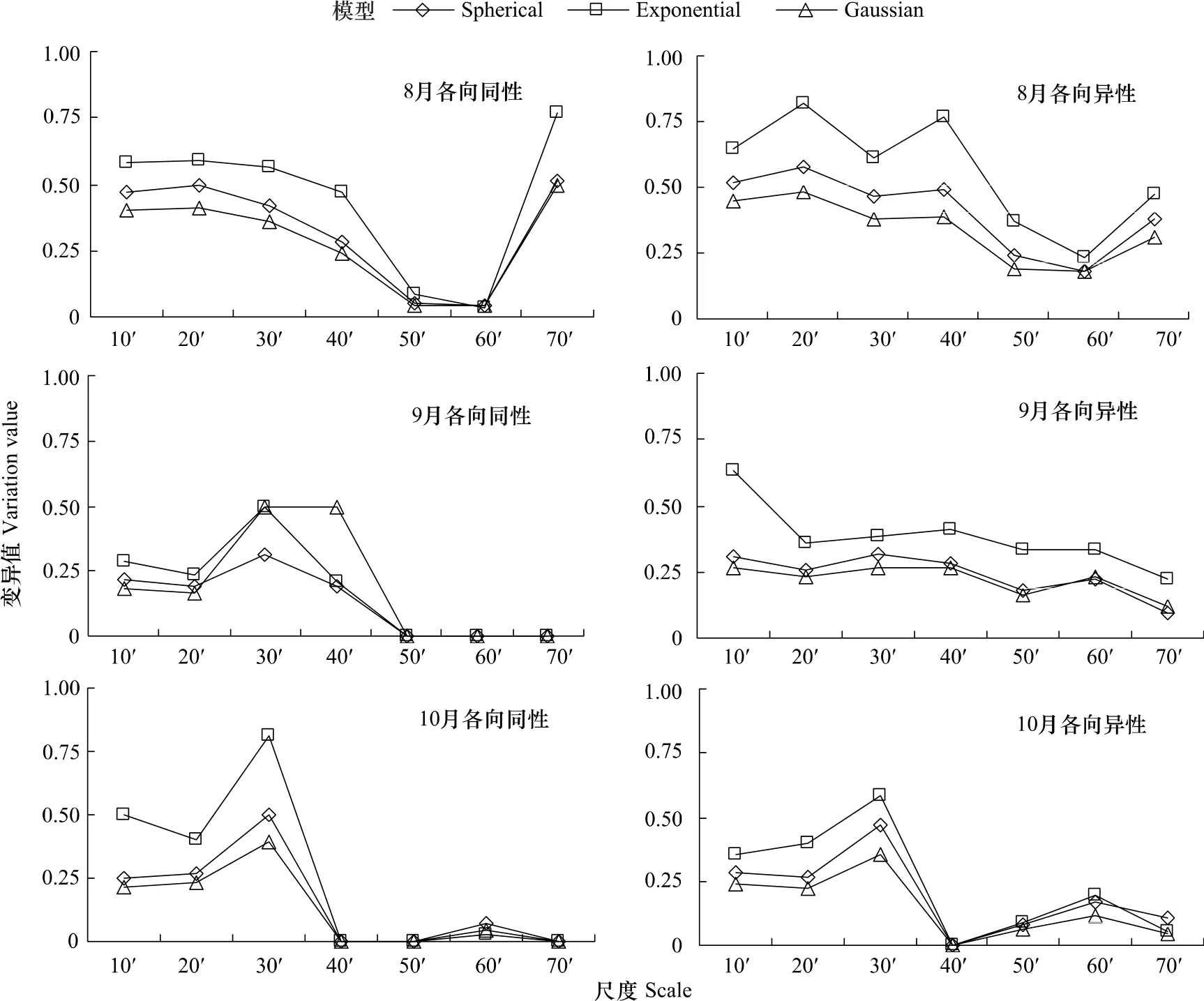
图1 各个尺度下各向同、异性比较Fig.1 Comparison of isotropic and anisotropic in various scales
根据建模误差最小原则分别选择了各月每一尺度的最优半变异函数模型。从8—10月最优模型在各个尺度下所占的比重来看(图2),球状模型占总数的29.4%,指数模型占47.1%,高斯模型占23.5%。球状模型比较适合中尺度下的柔鱼CPUE研究,指数模型则在小尺度的CPUE研究中为最适,其中尺度30'×30'时为最优,比重达到100%。而高斯模型对各个尺度则无明显偏好性,在 20'×20'、40'×40'、60'×60'、70'×70'尺度均有一定的比重。
从表3可知,小尺度下,8—10月的理论模型以指数模型为主,高斯模型次之,球状模型最少。中尺度下,以球状模型居多,指数模型较少,说明尺度变大后柔鱼资源丰度的空间分布也随之改变。总体上来看(表3),10月的块金值为最小,9月为最大,这说明10月份由随机变异引起的CPUE空间异质性程度较小。变程值基本上随着尺度的变大而变大,在各个月份中,10月的变程在各个尺度下均要小于其他2个月。
(3)方向角
从图3可知,柔鱼CPUE在空间上具有各向异性。在不同的尺度下,8月的方向角基本稳定在280°—320°之间,为西北—东南走向;9月在10'×10'尺度时方向角为280°左右,为西北—东南走向,其它尺度时均为东北—西南走向;10月在10'×10'和30'×30'尺度时方向角分别为282°和312°,为西北—东南走向,其他尺度时为东北—南西走向。在10'×10'尺度下,其各月方向角走势基本不变;而其它尺度下,各月方向角均发生了变化。
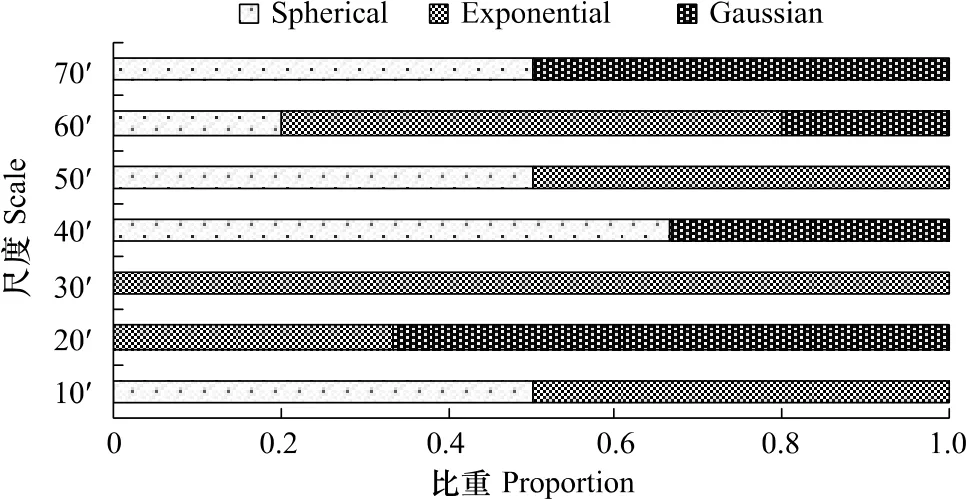
图2 最适模型在各个尺度下所占的比重Fig.2 Proportion of theoretical models in various scales
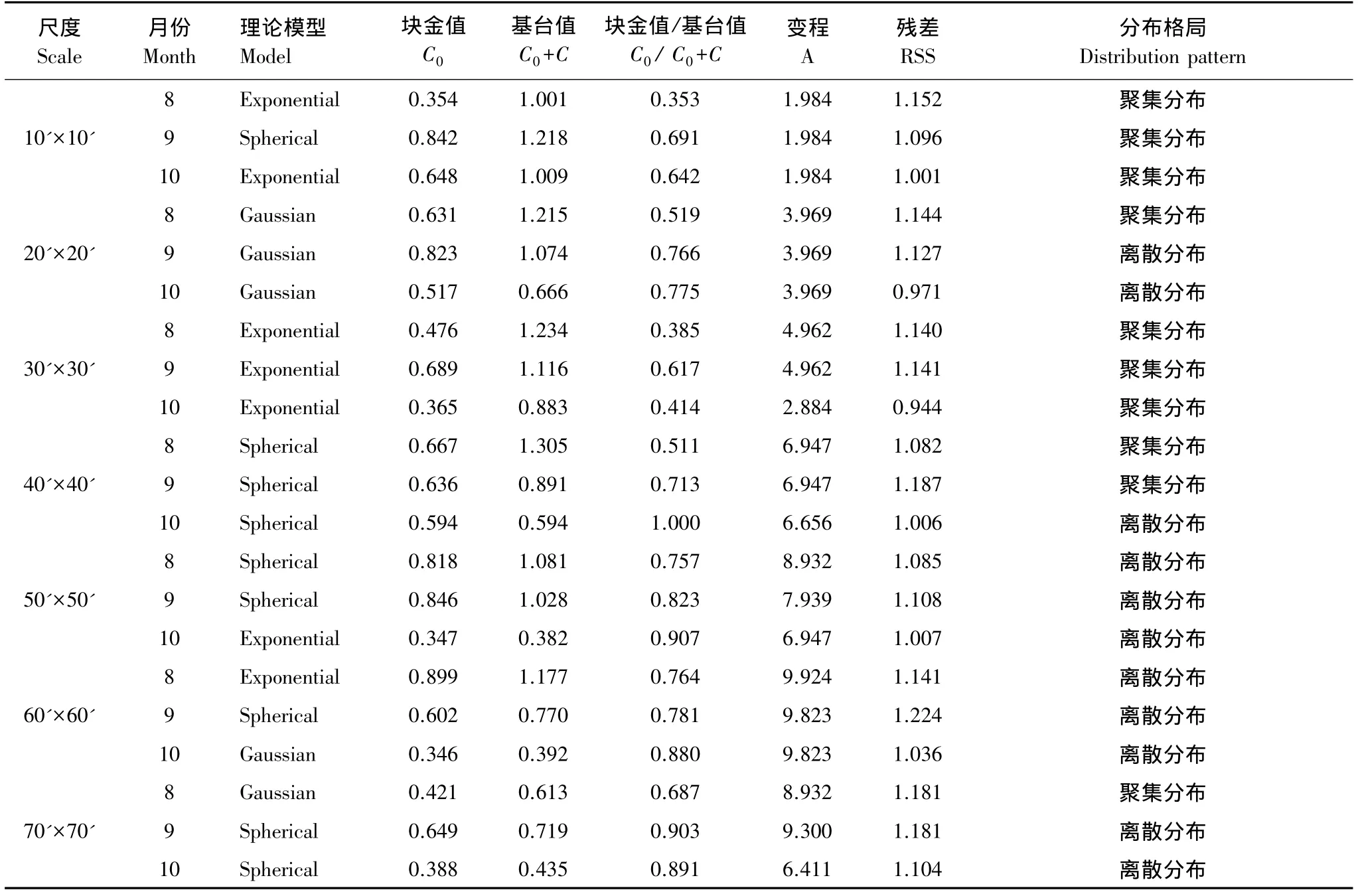
表3 半方差函数理论模型及相关参数Table 3 The semivariance theory model and related parameters
3 讨论与分析
3.1 适宜空间尺度的探讨
西北太平洋柔鱼生产统计数据具有多维性、动态性、多尺度等特点,同时具有强烈的时空相关性特点[16]。许多生态系统的空间异质性格局会随空间尺度大小的变化而变化[17-20],生态系统特性在不同尺度域上有着不同的变化速率,这种多尺度格局反映了生态系统的等级特征[18],因此利用地统计学来研究西北太平洋柔鱼资源丰度及其时空分布格局,采用适宜的空间尺度是极为重要的。从本文研究结果来看,8—10月在中尺度的 50'×50'、60'×60'、70'×70'下,柔鱼 CPUE 均表现为弱的空间自相关异质性,其变异值基本上在25%以下,这说明在低分辨率情况下不能很好地表达柔鱼群体本身固有的空间相关性,基本上表现为离散分布(表3)。在小尺度情况下,以经纬度30'×30'的尺度空间自相关性为最强且稳定,其值均在25%以上,均为聚集分布;尺度10'×10'的空间自相关性为次之,均为聚集分布;尺度20'×20'的空间自相关性则相对不稳定些,9月和10月柔鱼资源丰度的分布格局表现为离散分布(表3)。中小尺度下空间自相关性强弱显示,小尺度下空间自相关性普遍较强,更为适合,而中尺度次之。其中,尺度30'×30'在本研究中显示最为适宜。比杨晓明等[21]的西印度洋黄鳍金枪鱼1°×1°以及李灵智等[22]的大西洋金枪鱼5°×5°研究尺度小很多。由于不同尺度空间所表达的信息和含义及其特征均会有所差异,因此在今后研究中需要结合海洋环境条件及其所表达的渔业信息特征,对海洋实体或现象重点进行分析[23]。
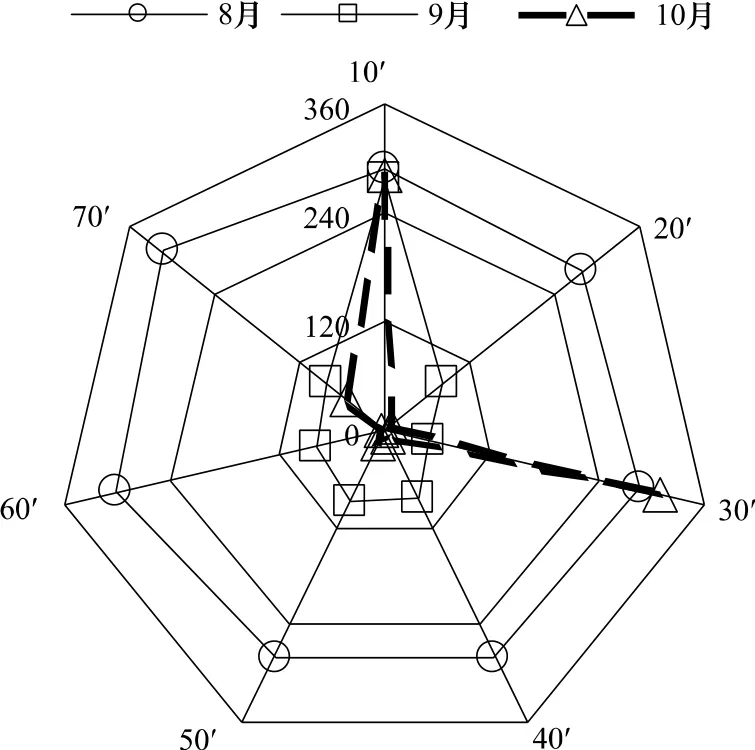
图3 各个尺度下方向角变化趋势Fig.3 Direction angle trends in various scales
3.2 时间和空间异质性特征分析
近年来,空间信息技术在海洋渔业领域的不断应用和深化[16]。对柔鱼CPUE数据可以按年、月、旬、周、日等多种时间分辨率进行数据平均或差异计算,本文按月进行了计算,张寒野[13]、杨晓明[21]等也采用月作为时间分辨率并取得较好研究结果。Tian等[24]研究认为,采用时间分辨率为双周的渔获统计数据和海洋遥感数据,比较适合栖息地指数模型的建立。
研究已表明,地球上的动物等都能够很好地适应环境的异质性,异质性的环境条件有利于大多数生物的生存[25]。从本研究结果来看,柔鱼CPUE空间分布有着明显的空间格局,中小尺度下指数模型拟合的空间自相关性基本为强,也存在少量的中等空间自相关。这或许与柔鱼是一种对环境极为敏感的种类且进行大规模的南北洄游有关,柔鱼广泛分布在整个北太平洋海域,不同种群的洄游路线、洄游时期也不同[26],本研究样本主要集中在黑潮向北或东北移动的暖水分支海域及周围,因此其空间分布格局与黑潮的势力及其与亲潮形成的交汇区密切相关。本研究认为,小尺度下,其变程值A都不大,均在5以下,其分布格局大部分表现为聚集分布(表3),而中尺度下变程值A都比较大,其值在6—10之间,分布格局大多表现为离散分布(表3),随机程度较高。这说明西北太平洋柔鱼资源在小范围内的空间分布较为密集,大范围内可能有多个群体组成,而群体之间存在着差异,这一结果与西北太平洋复杂的海洋环境条件是一致的[26]。
3.3 海洋环境变化对柔鱼时空异质性的影响
西北太平洋柔鱼渔场的形成不仅与温度、盐度等密切相关,而且还与海流、锋面、叶绿素浓度、涡旋等有密切的关系,这些因子共同构成了柔鱼的生存环境,因此我们在柔鱼资源丰度空间分布的分析与研究时应予以充分考虑。研究资料表明[26-27],柔鱼渔场是由黑潮和亲潮交汇形成的,柔鱼通常在夏季随着黑潮北上,进行索饵洄游,因此黑潮北上的势力强弱和走向将影响到柔鱼的洄游分布和渔场形成[28-33]。黑潮大弯曲年份往往会带来较好的柔鱼产量[34]。数据表明,2011年为黑潮非大弯曲年份,5月下旬至10月初黑潮总体上为平直,仅小部分时间产生小弯曲,而亲潮势力从7月开始一直都处于较低水平。8月和9月柔鱼资源分布在空间上的方向角均比较平稳,分别为西北—东南走向和东北—西南走向,这与8月和9月黑潮势力稍强于亲潮但总体上较为平缓的海流条件非常吻合。而10月柔鱼资源分布的方向角在各个尺度下偏差较大,有多个方向的走向,这可能是由于10月初黑潮产生小弯曲、亲潮与黑潮交汇激烈,以及柔鱼因性成熟初步开始南下洄游等因素的影响结果。柔鱼的生长和栖息活动离不开海洋环境,其群体在海洋中的分布表现为异质性的分布格局,同样,海洋因子也有其空间特征,也有特定的分布格局,两种异质性格局相互反映了异质性分布特点,并且起着相互制约的作用。
本文分析了在各个尺度下柔鱼资源丰度分布产生的效应,对西北太平洋柔鱼资源分布的空间格局有了更深入的了解,并对其空间格局的异质性特征和产生的原因做了初步的阐述。另外,水温垂直结构、海面高度等影响因子与西北太平洋柔鱼资源丰度的空间分布特征也有着一定的联系[1]。
[1]Chen X J,Tian SQ,Chen Y,Cao J,Ma J,Li S L,Liu B L.Fisheries Biology of Ommastrephes bartramii in North Pacific.Beijing:Science Press,2011.
[2]Wang Z Q.Geo-Statistics and Its Application in Ecology.Beijing:Science Press,1999.
[3]Turner M G.Landscape ecology:the effect of pattern on process.Annual Review of Ecology and Systematics,1989,20:171-197.
[4]Pickett ST A,Cadenasso M L.Landscape ecology:Spatial heterogeneity in ecological systems.Science,1995,269(5222):331-334.
[5]Liu D,Jiang Y,Liang W J.Spatial heterogeneity of exchangeable calcium content in cultivated soils of Shenyang suburbs.Ecology Magazine,2003,22(4):6-9.
[6]Isaaks E H,Srivastava R M.An Introduction to Applied Geostatistics.New York:Oxford University Press,1989.
[7]Webster R.Quantitative spatial analysis of soil in the field.Advances in Soil Sciences,1985,3:1-70.
[8]Rivoirard J,Simmonds J,Foote K G,Fernandes P,Bez N.Geostatistics for Estimating Fish Abundance.New York:Wiley Blackwell,2000.
[9]Petitgas P.Geostatistics in fisheries survey design and stock assessment:models,variances and applications.Fish and Fisheries,2001,2(3):231-249.
[10]Nishida T,Chen D G.Incorporating spatial autocorrelation into the general linear model with an application to the yellowfin tuna(Thunnus albacares)longline CPUE data.Fisheries Research,2004,70(2/3):265-274.
[11]Su F Z,Zhou C H,Zhang T Y,Du Y Y,Yao C Q.Spatial heterogeneity of pelagic fishery resources in the East China Sea.Chinese Journal of Applied Ecology,2003,14(11):1971-1975.
[12]Su F Z,Zhou CH,Shi W Z,Du Y.Spatial heterogeneity of demersal fish in East China Sea.Chinese Journal of Applied Ecology,2004,15(4):683-686.
[13]Zhang H Y,Hu F.Spatial heterogeneity of Todarodes pacificus in East China Sea in winter.Chinese Journal of Ecology,2005,24(11):1299-1302.
[14]Zhang H Y,Cheng JH.Geostatistical analysis on spatial patterns of small yellow croaker(Larimichthys polyactis)in the East China Sea.Journal of Fishery Sciences of China,2005,12(4):419-423.
[15]Wang H B,Yang Q,Liu Z J,Yang C H.Determining optimal density of grid soil-sampling points using computer simulation.Transactions of the Chinese Society of Agricultural Engineering,2006,22(8):145-148.
[16]Ji M,Jing F X,Li Y L,Zhang L P.Multi-scale generalization and expression of ocean fishery thematic attribute data.Bulletin of Surveying and Mapping,2004,(6):28-31.
[17]Turner M G,O'Neil R V,Gardner R H,Milne B T.Effects of changing spatial scale on the analysis of landscape pattern.Landscape Ecology,1989,3(3/4):153-162.
[18]O'Neil R V,Gardner R H,Milne B T.Heterogeneity and spatial hierarchies//Kolasa J,Pickett ST A,eds.Ecological Heterogeneity.New York:Springer-Verlag,1991:85-96.
[19]Qi Y,Wu J G.Effects of changing spatial resolution on the results of landscape pattern analysis using spatial autocorrelation indices.Landscape Ecology,1995,11(1):39-50.
[20]O'Neil R V,Hunsaker CT,Timmins SP,Jackson B L,Jones K B,Riitters K H,Wickham JD.Scale problems in reporting landscape pattern at the regional scale.Landscape Ecology,1996,11(3):169-180.
[21]Yang X M,Dai X J,Zhu GP.Geostatistical analysis of spatial heterogeneity of yellowfin tuna(Thunnus albacares)purse seine catch in the western Indian Ocean.Acta Ecologica Sinica,2012,32(15):4682-4690.
[22]Li L Z,Wang L,Liu J,Liu Q,Huang H L.Geostatistical analysis of tuna(Thunnusobesus)longline fishing grounds in the Atlantic Ocean.Journal of Fishery Sciences of China,2013,20(1):198-204.
[23]Xiang QH.Study on Distant Water Fishery Production Networks from Different Spatial Scales[D].Shanghai:East China Normal University,2011.
[24]Tian SQ,Chen X J,Chen Y,Xu L X,Dai X J.Evaluating habitat suitability indices derived from CPUE and fishing effort data for Ommatrephes bratramii in the Northwestern Pacific Ocean.Fishery Research,2009,95(2/3):181-188.
[25]Chen Y F,Dong M.Spatial heterogeneity in ecological systems.Acta Ecologica Sinica,2003,23(2):346-352.
[26]Chen X J,Xu L X.Analysis of relationship between fishing ground of Ommastrephes bartramii and surface water temperature and its vertical distribution from 150°E to 165°E in the northwestern Pacific.Transaction of Oceanology and Limnology,2004,(2):42-43.
[27]Wang Y G,Chen X J.The Resource and Biology of Economic Oceanic Squid in the World.Beijing:Ocean Press,1999.
[28]Dong Z Z.The World Oceans Economic Cephalopod Biology.Ji'nan:Shandong Science and Technology Press,1991.
[29]Chen X J,Liu B L.Analysis on catch of Ommastrephesbatrami in squid jigging fishery and the relationship between fishing ground and SST in the north Pacific Ocean in 2004.Journal of Zhanjiang Ocean University,2005,12(6):43-44.
[30]Chen X J.An approach to the relationship between the squid fishing ground and water temperature in the northwestern Pacific.Journal of Shanghai Fisheries University,1995,4(3):181-185.
[31]Chen X J.An analysis on marine environment factors of fishing ground of Ommastrephes batrami in northwestern Pacific.Journal of Shanghai Fisheries University,1997,6(4):263-267.
[32]Chen X J.The preliminary study on fishing ground of large-sized Ommastrephes batrami in North Pacific waters between 160°E and 170°E.Journal of Shanghai Fisheries University,1999,8(3):197-201.
[33]Chen X J,Tian SQ.Analysis and discussion on fishing ground of Ommastrephesbartrami in northwestern Pacific.Fisheries Modernization,2001,3(3):3-6.
[34]Shen H M.Summary of sea conditions in the squid fishing ground from June to December of 1999 in the North Pacific.Distant-Water Fisheries,2000,(3):15-18.
参考文献:
[1]陈新军,田思泉,陈勇,曹杰,马金,李思亮,刘必林.北太平洋柔鱼渔业生物学.北京:科学出版社,2011.
[2]王政权.地统计学及在生态学中的应用.北京:科学出版社,1999.
[5]刘丹,姜勇,梁文举.沈阳市郊耕地土壤交换性Ca含量的空间异质性特征.生态学杂志,2003,22(4):6-9.
[13]张寒野,胡芬.冬季东海太平洋褶柔鱼的空间异质性特征.生态学杂志,2005,24(11):1299-1302.
[16]季民,靳奉祥,李云岭,张连蓬.海洋渔业专题属性数据多尺度综合与表达.测绘通报,2004,(6):28-31.
[21]杨晓明,戴小杰,朱国平.基于地统计分析西印度洋黄鳍金枪鱼围网渔获量的空间异质性.生态学报,2012,32(15):4682-4690.
[22]李灵智,王磊,刘健,刘勤,黄洪亮.大西洋金枪鱼延绳钓渔场的地统计分析.中国水产科学,2013,20(1):198-204.
[23]向清华.不同空间尺度下的远洋渔业生产网络研究[D].上海:华东师范大学,2011.
[25]陈玉福,董鸣.生态学系统的空间异质性.生态学报,2003,23(2):346-352.
[26]陈新军,许柳雄.北太平洋(150°—165°E)海域柔鱼渔场与表温及水温垂直结构的关系.海洋湖沼通报,2004,(2):42-43.
[27]王尧耕,陈新军.世界大洋性经济柔鱼资源及其渔业.北京:海洋出版社,2005.
[28]董正之.世界大洋经济头足类生物学.济南:山东科学技术出版社,1991.
[29]陈新军,刘必林.2004年北太平洋柔鱼钓产量分析及作业渔场与表温的关系.湛江海洋大学学报,2005,12(6):43-44.
[30]陈新军.西北太平洋柔鱼渔场与水温因子的关系.上海水产大学学报,1995,4(3):181-185.
[31]陈新军.关于西北太平洋的柔鱼渔场形成的海洋环境因子的分析.上海水产大学学报,1997,6(4):263-267.
[32]陈新军.北太平洋(160°E—170°E)大型柔鱼渔场的初步研究.上海水产大学学报,1999,8(3):197-201.
[33]陈新军,田思泉.西北太平洋海域柔鱼渔场分析探讨.渔业现代化,2001,3(3):3-6.
[34]沈惠明.1999年6—12月北太平洋鱿钓渔场海况总结.远洋渔业,2000,(3):15-18.
猜你喜欢
海洋通报(2022年6期)2023-01-07
中国水产(2021年12期)2021-12-06
大自然探索(2021年12期)2021-02-07
学生天地(2020年19期)2020-06-01
当代水产(2019年4期)2019-05-16
中国石油石化(2018年17期)2018-11-10
散文诗(2017年15期)2018-01-19
海洋世界(2017年1期)2017-02-13
渔业致富指南(2016年12期)2016-11-11
海洋科学(2016年1期)2016-07-06
