
百山祖自然保护区植物群落beta多样性
2013-12-09 05:58:36谭珊珊叶珍林袁留斌周荣飞金孝锋于明坚
生态学报 2013年21期
谭珊珊,叶珍林,袁留斌,周荣飞,胡 广,金孝锋,于明坚,*
(1. 浙江大学生命科学学院,杭州 310058; 2. 凤阳山-百山祖国家级自然保护区百山祖管理处,庆元 323800; 3. 丽水学院生态学院,丽水 323000; 4. 杭州师范大学生命与环境科学学院,杭州 310036)
百山祖自然保护区植物群落beta多样性
谭珊珊1,叶珍林2,袁留斌3,周荣飞2,胡 广1,金孝锋4,于明坚1,*
(1. 浙江大学生命科学学院,杭州 310058; 2. 凤阳山-百山祖国家级自然保护区百山祖管理处,庆元 323800; 3. 丽水学院生态学院,丽水 323000; 4. 杭州师范大学生命与环境科学学院,杭州 310036)
采用植物群落学的典型样方法,研究了百山祖自然保护区森林植物群落beta多样性格局及其维持机制。通过对45个20 m×20 m标准样地的调查数据进行分析,运用Chao′s群落距离指数衡量该植物群落beta多样性格局,并通过Mantel检验、基于距离矩阵的偏RDA分析和方差分解等方法初步检验和衡量了各环境因子差异(包括群落郁闭度、海拔、坡度、坡向和坡位)和群落空间距离对该区域beta多样性格局的影响。结果显示,该区域内植物群落beta多样性随着群落间综合环境差异或群落空间距离的增加而增大,但环境差异和群落空间距离只能解释36%左右的beta多样性格局。检验的5个环境因子中,只有群落郁闭度和海拔对百山祖自然保护区植物群落beta多样性有显著影响,并且群落郁闭度对beta多样性的解释度(20.0%)略高于海拔对beta多样性的解释度(18.0%)。群落空间距离对百山祖自然保护区beta多样性的解释度最小(9.0%)。展现了百山祖自然保护区内植物群落beta多样性格局及其与群落环境和空间距离的关系,所获得的结果支持生境异质性和扩散限制联合对植物群落beta多样性起作用的假说。
百山祖自然保护区;beta多样性;环境因子;空间距离;森林植物群落
Beta多样性是指区域环境梯度下生物群落物种组成的差异[1- 3],是生物多样性的3个尺度之一。它不仅能代表区域物种多样性格局及周转率,还能反映物种-环境之间的关系,故对其进行深入研究能够更好地认识生物群落,进而更好地保护和管理生物多样性[4- 7]。从Whittaker首次提出beta多样性这一概念开始[8],对beta多样性的研究历史已有50多年,但至今对于它的形成和维持机制仍然存在争议[9- 13]。
目前对beta多样性维持机制的争议主要集中在两大主要生态学过程,即生态位分化和扩散限制[9,11]。首先,经典生态位理论认为物种共存的先决条件是生态位分化,共存物种需要适应不同的环境,并在某些特定的环境中占据竞争优势。因此,当群落赖以生存的生境异质性越大时,群落结构的差异也应该会随之增加,大多数实验研究也支持这一理论[14- 16]。另一方面,扩散限制也会影响群落beta多样性格局,因为扩散限制作用(例如种子扩散限制等[17])会限制种子的扩散距离,故物种更替速率会随着空间距离的增加而增加,最终提高大尺度上的beta多样性。这一理论推测也得到了很多实验的证实[18- 19]。以Hubbell为代表的中性理论认为单一的扩散限制能产生与实际群落十分类似的beta多样性格局,因而传统的生态位理论受到了挑战,两个理论间的差异也激发了一大批生态学家重新关注和验证群落beta多样性的形成和维持机制[11- 12,20]。
此外,还有相当一部分研究表明生态位分化和扩散限制可能同时对群落beta多样性格局产生影响[12,20- 23]。例如Tuomisto等人对亚马逊热带雨林beta多样性的研究显示,生境异质性、生态位分化对不同研究地点植被组成差异的解释度大于扩散限制作用的解释度,二者结合能够解释70%—75%的物种多样性差异,并且得出扩散作用在80 km内对群落植物物种组成具有强烈的作用,超过这个范围主要是环境因子起主要作用[20]。
基于上述存在的有关beta多样性维持机制的争论,本文将关注浙江百山祖自然保护区内植物群落beta多样性的形成机制,具体研究5个生境因子以及群落空间距离对植物群落beta多样性的影响,以期能为亚热带地区的beta多样性形成和维持机制的研究提供事实证据。本文通过对植物群落调查样地数据的分析,回答以下问题:(1)百山祖自然保护区内植物群落beta多样性格局是什么?(2)群落空间距离和群落环境差异是否对这一beta多样性格局有显著影响?(3)若综合环境因子具有显著作用,具体是5个环境因子中的哪些在起作用?
1 野外调查与数据分析
1.1 研究地概况
百山祖自然保护区为“浙江凤阳山-百山祖国家级自然保护区”的一个组成部分,地处我国东南沿海的闽浙丘陵区,由华夏古陆华南地闽浙地盾演变而成。地貌以侵蚀地貌为主,山地坡度大多在30°以上,多深谷陡坡。属中亚热带湿润季风气候,年降水量2341.8 mm,年均温12.8 ℃,无霜期187 d。保护区800 m以下为红壤,800 m以上以黄壤为主。植被在全国植被分区中属“中亚热带常绿阔叶林南部亚地带”,地带性植物为亚热带常绿阔叶林[24- 25]。百山祖自然保护区的核心区分为两块:一块位于庆元县东北部,以百山祖为中心,面积为10440.7 hm2,另一块位于庆元县南部的安南乡五岭坑,面积仅439.4 hm2。
1.2 植物群落及环境数据调查
为定量计算百山祖自然保护区内植物群落beta多样性格局,依据百山祖自然保护区GIS地形数据和实地勘察结果,我们在保护区的不同区域设置了45个20 m×20 m的森林群落样地,并依据经典植物群落学方法对每个植物样地进行了详细的调查(图1)。首先进行乔木调查,调查前先将每个20 m×20 m的样地划分为16个5 m×5 m的小样方。在每个小样方内对树高≥5 m的乔木层个体进行每木调查,记录每个个体的物种名、所处小样方号、胸径、高度、冠幅等。然后进行8个灌木样方的调查,灌木调查的样方大小均为2 m×2 m,分别分布在8个不相邻小样方的右下角;记录每一灌木样方内每种灌木的物种名、平均高度和盖度;调查草本时样方大小设置为1 m×1 m,镶嵌在每个灌木样方右下角处,同样记录每个草本样方内各草本物种的物种名、平均高度和盖度。

图1 百山祖自然保护区内植物群落调查样地分布图Fig.1 Spatial distribution of 45 invesitigated plots (black points with plot name labels) in two areas (top left panel shows the Zhejiang province) of Baishanzu Nature Reserve45个植物样方主要分布在百山祖自然保护区的两块区域(左上图为浙江省地图); 具体两块区域内植物样方的详细分布情况见左下及右图,其中每一黑点表示一个典型植物样方,边上并配有该样方的编号
同时,记录每个样地的环境因子以及空间位置。环境因子包括样地内的植物群落郁闭度(作为群落内光照特征的综合参数)、海拔、坡度、坡向和坡位。坡度和坡向均采用地质罗盘仪测量。坡向用0°—360°表示(0°表示正北方向、90°为正东方向、180°为正南方向、270°表示正西方向),分析时经过正弦转化;样地坡位分为7个等级,分别为沟谷、下坡、中下坡、中坡、中上坡、上坡、山顶或山脊,并按此顺序转变为数值变量,分别用1—7数字表示;样地空间位置信息用每一样地中心位置的经纬度表示。
1.3 植物群落beta多样性指数
采用Chao′s群落距离指数度量百山祖自然保护区内植物群落beta多样性。该指数能够综合考虑不同样地中物种丰富度及物种多度信息,且可在一定程度上考虑未调查到的稀有种对群落beta多样性的影响,因而能较合理地衡量百山祖自然保护区内植物群落的beta多样性大小[26]。若给定i、k两个植物样地,Chao′s群落距离指数可以简单的表示为:1-Ui×Vk/(Ui+Vk-Ui×Vk),Ui=Ci/Ni+(Ni-1)/Ni×a1/2/a2×Si/Ni,Vk的计算过程与Ui类似。其中Ci是样方i中的且也出现在样方k中所有物种的总个体数。Ni是样方i中所有物种的总个体数。a1和a2是仅在样方k中一个和两个个体,且在样方i中出现的总物种数。Si是在样方k中仅有一个个体且出现在样方i中的总个体数。若植物群落beta多样性越高,则不同植物群落组成差异越大,Chao′s群落距离指数也会越大,并趋向于1;反之则趋向于0。最终,根据45个植物群落数据,可获得45×45的植物群落Chao′s距离矩阵。Chao′s群落相似性指数则为1-Chao′s群落距离指数。在Chao′s群落距离计算中,已通过等密度放大法一定程度上矫正了因乔灌草不同取样面积而带来的误差。
1.4 环境因子矩阵和空间距离矩阵
依据每个植物群落样地的5个环境因子,标准化后(即每一环境因子具有同等的权重)可获得一个45×45的植物群落综合环境距离矩阵。为分析每一环境因子对植物群落beta多样性的影响,也可根据每一环境因子构建5个45×45的植物群落综合环境距离矩阵。同理,根据每一植物样地的经纬度信息,可获得一个45×45的植物群落空间距离矩阵,其中植物群落的空间距离为植物群落间的地表球面距离。所有这些距离矩阵都是对称矩阵,且每一元素与植物群落距离矩阵一一对应。以植物群落距离矩阵为x轴,群落综合环境矩阵或群落空间距离矩阵为y轴,可初步查看植物群落距离矩阵与植物群落综合环境矩阵或群落空间距离矩阵的关系。
1.5 综合环境因子和空间距离对植物群落beta多样性影响的显著性分析
本文采用Mantel检验回答植物群落综合环境或植物群落空间位置是否对植物群落beta多样性有显著影响。以植物群落综合环境为例,Mantel检验分析植物群落综合环境矩阵与植物群落Chao′s距离矩阵之间是否存在显著相关性。具体过程为先计算这两个矩阵对应元素间的观测Person相关性系数,然后通过随机重置某一矩阵中所有元素在矩阵中的位置来构建这一观测矩阵的随机矩阵,之后计算出随机矩阵与另一观测矩阵的Person相关系数。重复该随机置换过程999次即可获得零假设下(两个矩阵无相关性)Person相关系数的分布。最后将零假设下的Person相关系数从小到大排列,若实际观测Person相关系数位于第25或第975位之外(5%显著水平),则可认为实际观测矩阵间存在显著相关性,反之则不存在显著相关性。采用同样的Mantel检验,可以检验植物群落空间距离矩阵与植物群落beta多样性是否存在显著关系。
然而上述的Mantel检验无法回答群落环境本身或空间距离本身是否对植物群落beta多样性产生显著影响。例如,当综合环境因子本身对植物群落beta多样性无显著影响,而与综合环境因子高度正相关的空间距离对植物群落有显著影响时,上述Mantel检验仍可能会得到综合环境因子对植物群落距离有显著影响的结果。因此,在完成Mantel检验后,本研究还采用了偏Mantel检验以探索环境因子本身以及空间距离本身对植物群落beta多样性的影响。偏Mantel检验是在上述Mantel检验的基础上,运用类偏相关性检验的思想,通过控制群落空间距离对植物群落的影响来检验群落环境因子本身对植物群落距离矩阵是否有显著影响。
1.6 各因素对植物群落beta多样性影响强弱分析
为进一步探索各环境因子或空间距离对植物群落beta多样性影响的强弱,采用了基于距离矩阵的偏RDA分析和RDA分析[27]。其中偏RDA分析可在控制其他因子的影响下,检验某一因子对应变量影响的显著性;而RDA分析可获得不同因子对应变量影响的强弱。基于距离的含义为两种分析中应变量均为不同样地间Chao′s距离矩阵的前两个PCA主轴;RDA分析中自变量为不同样地的环境因子矩阵和一个空间距离矩阵。该自变量矩阵的每一行代表一个样地,每一列表示一种环境因子或空间位置;其中空间位置由二维的经纬度信息通过PCA转化为一维信息(保留了95%的空间位置信息)。偏RDA分析中自变量为不同样地的目标因子向量,条件自变量是除去目标因子列的RDA分析自变量矩阵。先通过偏RDA检验挑选显著影响植物群落beta多样性的影响因子,再将具有显著影响作用的因子纳入RDA分析,最终可从RDA图中大致获得各因子对植物群落beta多样性影响作用的强弱。最后,定量分析了显著作用于植物群落beta多样性的每一相关因子的贡献率,从而定量回答不同影响因子对百山祖自然保护区植物群落beta多样性格局的重要性和相对强弱。
本文还采用上述相同的方法分析乔、灌、草3个层次中生境异质性和空间距离对各个层次beta多样性的影响,以期更进一步回答影响各层次beta多样性的主要因子。
2 结果
2.1 综合环境因子与空间距离对植物群落beta多样性的影响
Chao′s群落相似性随群落空间距离和群落环境距离的增加有下降的趋势(图2)。Mantel检验结果进一步显示群落空间距离(r=0.2515,P=0.001)和群落环境距离(r=0.2673,P=0.001)均对Chao′s植物群落距离产生显著影响,并且偏Mantel检验也得到了类似的结果(空间距离本身r=0.2231,P=0.003;环境距离本身r=0.2411,P=0.001)。这些结果都证明了百山祖自然保护区内植物群落beta多样性会随着植物群落空间距离和综合环境距离的增加而增加。
不过,图2中Chao′s群落相似性指数在不同空间尺度和不同环境距离间并没有显著差异,暗示着除了空间距离或环境距离对Chao′s群落相似性有影响外,可能还有其他一些重要因素也对Chao′s群落相似性产生影响。
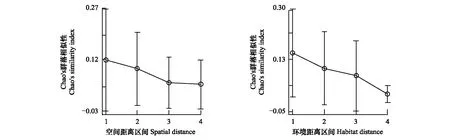
图2 百山祖自然保护区植物群落Chao′s差异指数与群落空间距离区间、群落综合环境距离区间的关系。Fig.2 Relationships between Chao′s different index of plant communities and environmental and spatial distance among different plots, in the Baishanzu Nature Reserve 空间距离区间为1:[0, 0.18], 2:(0.18, 0.35], 3:(0.35, 0.53], 4:(0.53, 0.70]; 环境距离区间为1:[0.11, 0.64], 2:(0.64, 1.16], 3:(1.16, 1.69], 4:(1.69, 2.21]; 图中圆点和上下垂直线段表示每个群落空间距离区间内Chao′s差异指数均值及其标准差
2.2 单个环境因子和空间距离对植物群落beta多样性的影响
通过偏RDA分析,发现5个环境因子中只有海拔和群落郁闭度两个环境因子对百山祖植物群落beta多样性产生极显著影响,而坡度、坡向和坡位并未检验出有显著作用。空间距离仍对百山祖植物群落beta多样性有显著影响(表1),与上述的Mantel检验和偏Mantel检验结果一致。但分层分析结果表明(表2—表4)乔木层中beta多样性主要受海拔和空间距离的显著影响,灌木层的beta多样性除了受海拔和空间距离的显著影响外,还受群落郁闭度的显著作用,草本层的beta多样性主要受海拔和群落郁闭度的影响,空间距离、坡度、坡向和坡位等因素均不起显著作用。
表1环境因子和空间距离对百山祖自然保护区内植物群落beta多样性影响的显著性检验
Table1Significanttestoftherelationshipsbetweenbetadiversityandfiveenvironmentalvariables,spatialdistancebetween45plotsintheBaishanzuNatureReserve

环境因子EnvironmentalvariablesF方差VarianceP海拔Altitude6.9740.00990.005∗∗坡向Slopeaspect0.68730.00100.47坡度Slopegrade1.80890.00260.18群落郁闭度Crowndensity11.2630.01600.005∗∗坡位Slopeposition0.39070.00060.7空间距离Spatialdistance2.45110.00350.0875∗
表2环境因子和空间距离对百山祖自然保护区内植物群落乔木层beta多样性影响的显著性检验
Table2Significanttestoftherelationshipsbetweenbetadiversityofthetreelayerandfiveenvironmentalvariablesandspatialdistancebetween45plotsintheplantcommunityofBaishanzuNatureReserve
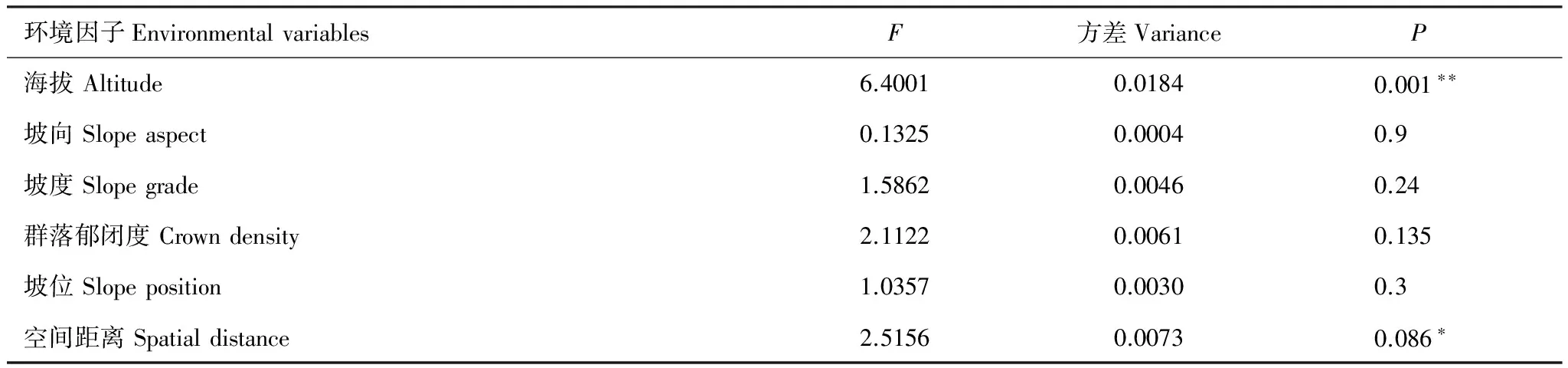
环境因子EnvironmentalvariablesF方差VarianceP海拔Altitude6.40010.01840.001∗∗坡向Slopeaspect0.13250.00040.9坡度Slopegrade1.58620.00460.24群落郁闭度Crowndensity2.11220.00610.135坡位Slopeposition1.03570.00300.3空间距离Spatialdistance2.51560.00730.086∗
表3环境因子和空间距离对百山祖自然保护区内植物群落灌木层beta多样性影响的显著性检验
Table3Significanttestoftherelationshipsbetweenbetadiversityoftheshrublayerandfiveenvironmentalvariablesandspatialdistancebetween45plotsintheplantcommunityofBaishanzuNatureReserve

环境因子EnvironmentalvariablesF方差VarianceP海拔Altitude4.70320.00760.0167∗∗坡向Slopeaspect0.7380.00120.48坡度Slopegrade2.4100.00390.15群落郁闭度Crowndensity8.87130.01430.005∗∗坡位Slopeposition1.26830.00200.26空间距离Spatialdistance2.96180.00480.061∗
表4环境因子和空间距离对百山祖自然保护区内植物群落草本层beta多样性影响的显著性检验
Table4Significanttestoftherelationshipsbetweenbetadiversityoftheherblayerandfiveenvironmentalvariablesandspatialdistancebetween45plotsintheplantcommunityofBaishanzuNatureReserve
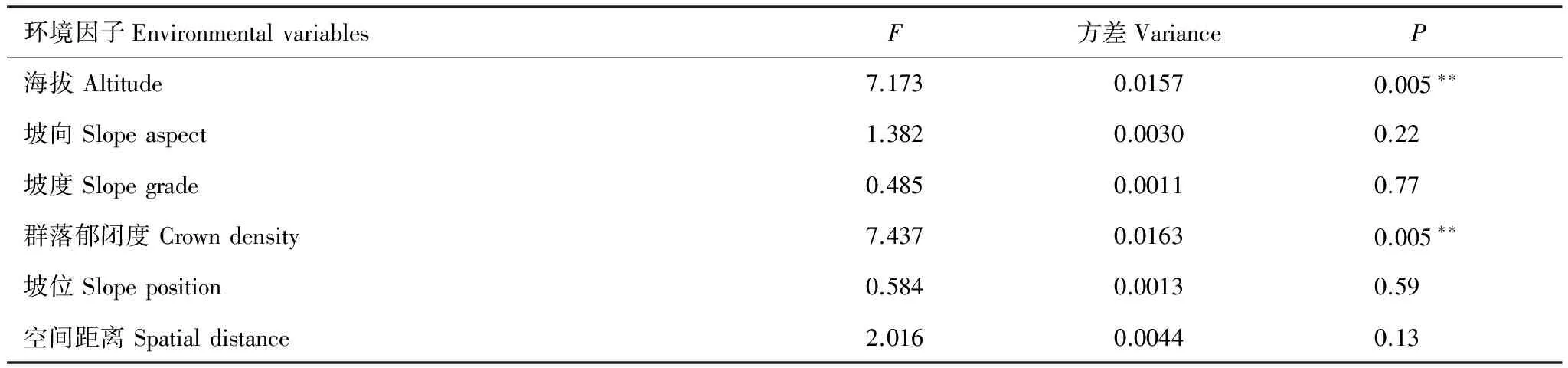
环境因子EnvironmentalvariablesF方差VarianceP海拔Altitude7.1730.01570.005∗∗坡向Slopeaspect1.3820.00300.22坡度Slopegrade0.4850.00110.77群落郁闭度Crowndensity7.4370.01630.005∗∗坡位Slopeposition0.5840.00130.59空间距离Spatialdistance2.0160.00440.13
图3为RDA分析结果,其中B1—B45代表样地序号,序号在图中的位置越接近表示该样地群落组成越相近。箭头表示影响因子,箭头连线在两个轴上的投影长度代表了某个影响因子对研究对象影响的大小,连线在轴上的投影越长代表这个影响因子对研究对象的分布影响越大,箭头连线与排序轴的夹角代表了某个影响因子与排序轴的相关性大小,夹角越小则相关性越高。计算结果显示,RDA分析中x轴的解释度为25.76%,y轴的解释度为18.52%,由图3可知5个环境因子和群落空间距离对植物群落beta多样性的影响强弱依次为:群落郁闭度、海拔、空间位置、坡度、坡位和坡向。
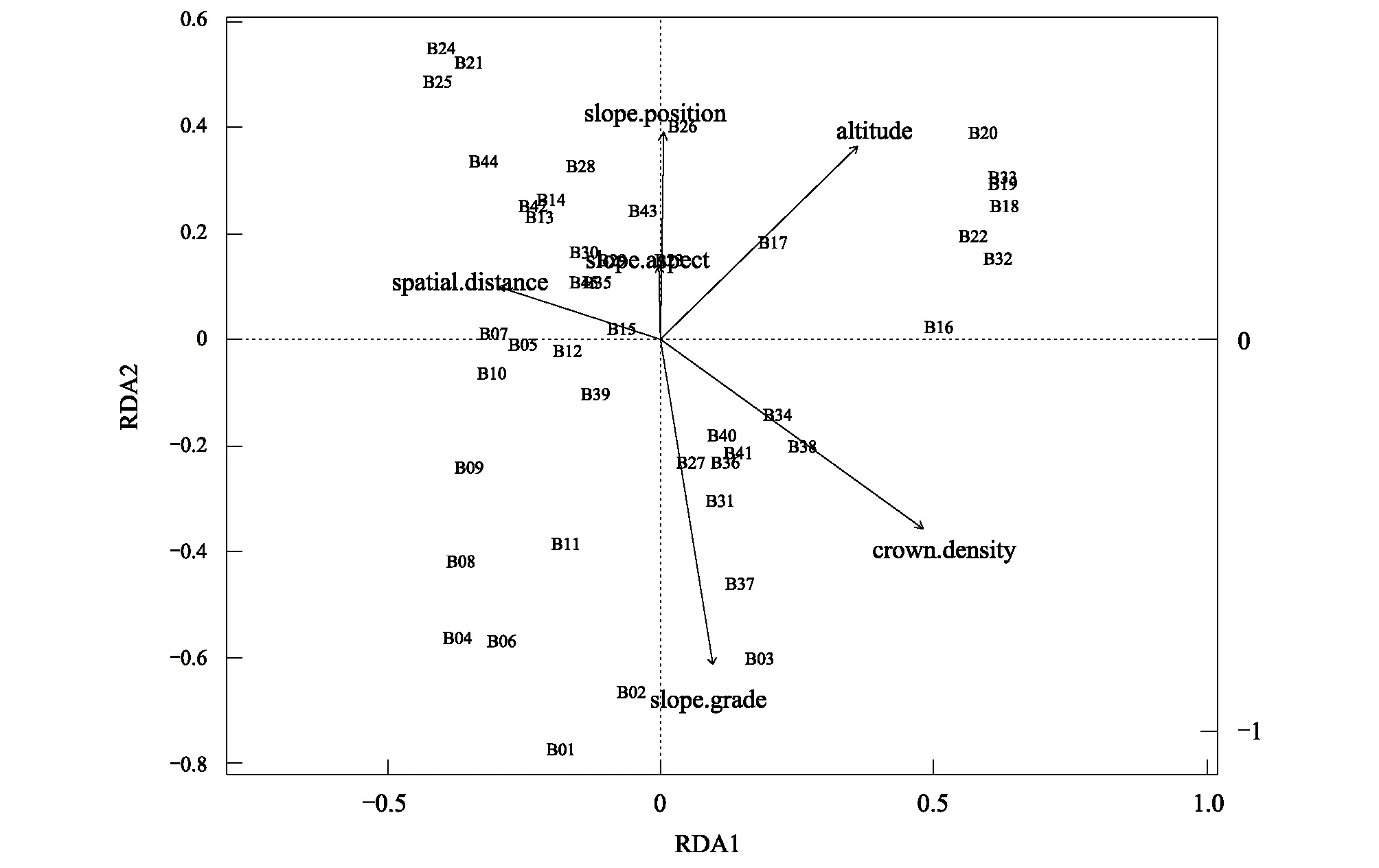
图3 空间距离和环境因子与百山祖自然保护区植物群落相似性的RDA分析Fig.3 Redundancy analyses (RDA) of plant community similarity in relation to spatial distance and environmental variables in the Baishanzu Nature Reserve

图4 海拔(a)、群落郁闭度(b)、空间距离(c)对百山祖自然保护区植物群落beta多样性的解释度Fig. 4 Venn diagram of altitude (a), crown density (b) and spatial distance (c) on the explanation of beta diversity pattern of plant communities in the Baishanzu Nature Reserve图中的数字表示对应区域对beta多样性格局的解释度; 其中a、b、c表示海拔本身、群落郁闭度本身和空间距离本身对beta多样性的影响;e表示群落郁闭度本身和空间距离的联合作用对beta多样性的影响(小于零的部分未列出)
由于群落郁闭度、海拔和空间位置均对该区域内植物群落beta多样性有显著作用,因此采用方差分解方法定量分析这3个显著相关因子对保护区内植物群落beta多样性的解释度。从图4中可以看到,群落郁闭度、海拔和空间距离一共解释了36%的beta多样性格局。这一结果与图2中出现较大的标准差相一致,共同暗示着可能还有其他重要的因素影响着百山祖自然保护区内植物群落beta多样性格局。从图4中还可以得到3个有显著作用的影响因子中,群落郁闭度对beta多样性的解释度最高,达到20%;其次为海拔的解释度18%;空间距离对该区域beta多样性解释度最低,仅有9%,这一结果与RDA排序分析结果一致。但需注意的是各影响因子对beta多样性格局的影响并不独立。例如群落郁闭度与海拔的联合作用只解释了27.9%的beta多样性格局(表5和图5),说明不同因子间可能存在着相关性。更详细的组合因子解释度见表5。
空间距离和环境因子与百山祖自然保护区植物群落乔灌草3个层次相似性的RDA分析显示,海拔和群落郁闭度对各群落3个层次相似性的影响较大,坡度、坡向和坡位的影响同样较小(图6—图8)。通过分析海拔、群落郁闭度和空间距离对各群落乔、灌、草3个层次beta多样性的解释度可以看出,对乔木层而言,海拔对beta多样性差异的解释度最大,达到26.0%,而群落其中X1表示取样样地海拔,X2表示取样群落的郁闭度,X3为取样群落间的空间距离;a—g表示与该表相对应的文恩图(图5)中所对应的字母区域郁闭度为13.0%,空间距离对该层次beta多样性解释度最低,只有8.0%。在灌木层和草本层中同样是海拔的解释度最大,分别为17.0%和20.0%,群落郁闭度的解释度均为15.0%,灌木层中空间距离的解释度(11.0%)大于草本层中空间距离的解释度(5.0%)(图9—图11)。
表5海拔、群落郁闭度与群落间空间距离对百山祖自然保护区植物群落beta多样性影响的方差分解
Table5ANOVAofelivation(X1),crowndensity(X2)andspaitaldistanceamongsurveiedplots(X3)ontheexplanationofbetadiversitypatternofplantcommunitiesintheBaishanzuNatureReserve

因子代码Factorcode自由度df拟合系数Rsquare调整后的拟合系数AdjustedRsquare[a+b+f+g]=X110.10630.0856[b+d+e+g]=X210.15200.1323[c+e+f+g]=X310.04500.0228[a+b+d+e+f+g]=X1+X220.31220.2794[a+c+d+e+f+g]=X1+X320.20630.1685[b+c+d+e+f+g]=X2+X320.21570.1783[a+b+c+d+e+f+g]=All30.39940.3555[a]=X1|X2+X310.1771[b]=X2|X1+X310.1869[c]=X3|X1+X210.0761[d]0-0.0314[e]00.0069[f]0-0.0300[g]0-0.0302[h]=Residuals0.6445[a+b]=X1|X310.1457[a+f]=X1|X210.1472[b+d]=X2|X310.1555[b+e]=X2|X110.1938[c+e]=X3|X110.0830[c+f]=X3|X210.0461
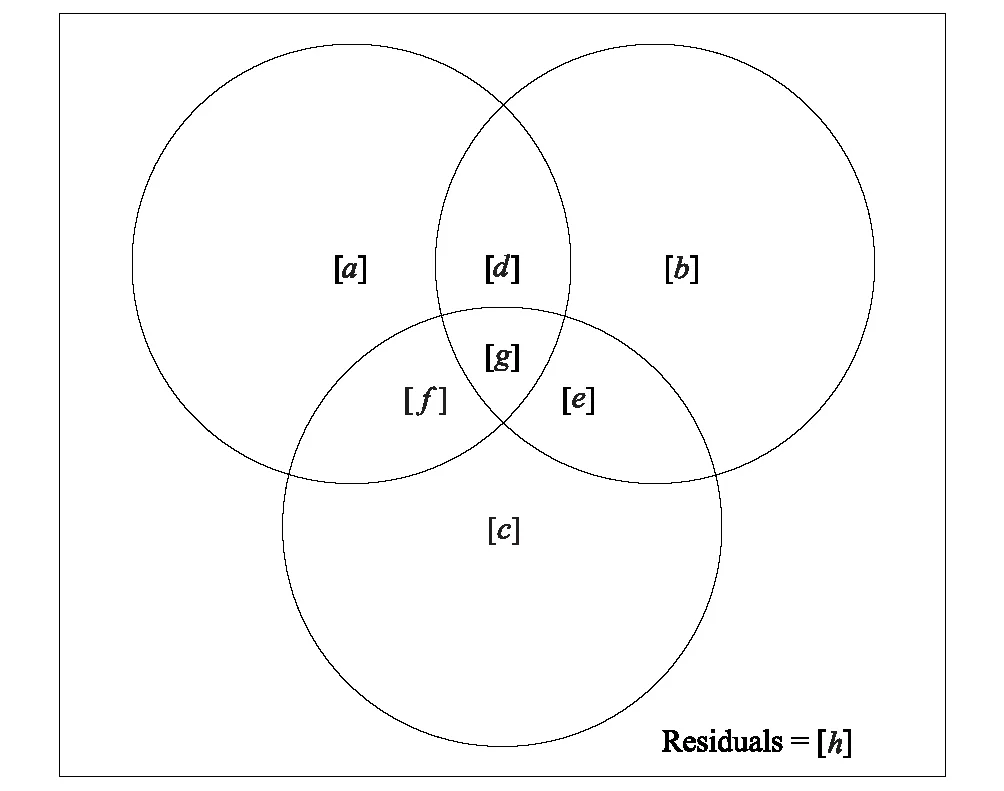
图5 表5中方差分解所对应的文恩图Fig.5 Venn diagram of ANOVA results in Table 5
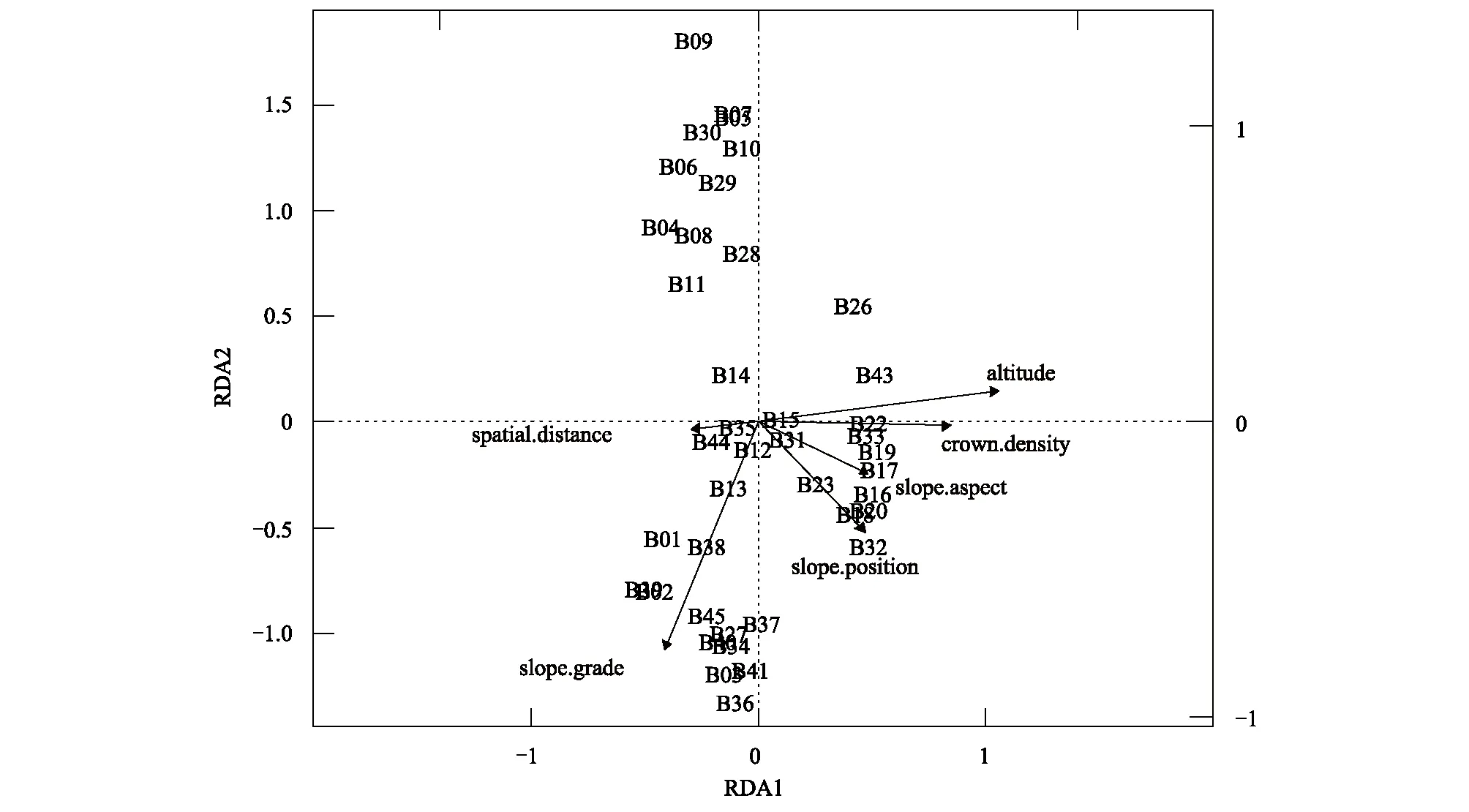
图6 空间距离和环境因子与乔木层相似性的RDA分析Fig. 6 Redundancy analyses (RDA) of the tree layer similarity in relation to spatial distance and environmental variables
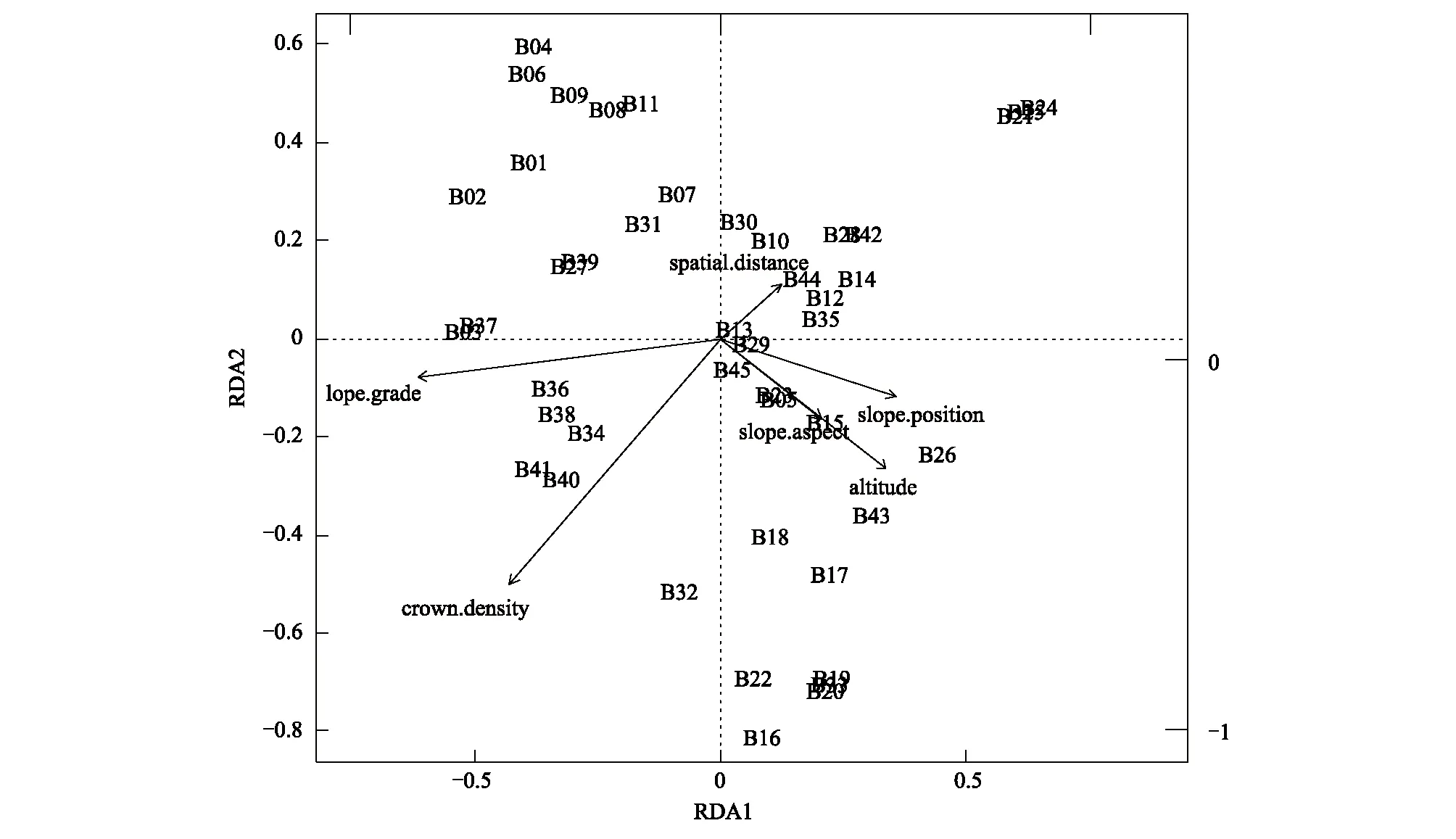
图7 空间距离和环境因子与灌木层相似性的RDA分析Fig. 7 Redundancy analyses (RDA) of the shrub layer similarity in relation to spatial distance and environmental variables

图8 空间距离和环境因子与草本层相似性的RDA分析Fig. 8 Redundancy analyses (RDA) of the herb layer similarity in relation to spatial distance and environmental variables
3 讨论
本研究显示,百山祖自然保护区的植物群落beta多样性同时受生境异质性和扩散限制的影响,这与最近的一些研究结果一致[12,20]。虽然在经典生境异质性理论或群落学中性理论中,beta多样性常被认为由单一的异质性生境或扩散限制引起,但实际群落中尤其是植物群落,更有可能同时受生境异质性和扩散限制的共同作用。例如,很多实验研究表明森林植物群落从种子传播开始就受到扩散限制的影响[17,28],而在随后种子萌发和幼苗生长等阶段都可能受异质性生境的影响[29- 31]。
通过基于距离的偏RDA分析和RDA分析,进一步发现群落郁闭度对beta多样性形成的作用最大,估计是因群落郁闭度影响群落下层光照和其它生境因子分布,及其与群落演替有密切的关系。郁闭度是森林中乔木树冠遮蔽地面的程度,能够反映林分密度以及群落演替的程度,郁闭度大的森林群落中光线较暗、直射到地面的阳光较少,使得幼苗和幼树的成活率较低[32- 33]。百山祖自然保护区郁闭度大的群落中,耐阴性强的乔木幼苗能够较好的生长,例如,对百山祖植物群落alpha多样性的分析发现(未发表),在群落郁闭度较高(gt;0.85)的常绿阔叶林中,优势种甜槠(Castanopsiseyrei)的幼苗因耐阴性强而在林下生长较好,种群能较好地更新,而草本层物种丰富度和物种多样性均较低,主要原因可能是大部分阳光被乔木层吸收和反射,到达地面的阳光较少。在马尾松群落中,由于群落郁闭度仅为0.65,有较多的光线能够透过乔木层到达地面,因此该样地中草本层盖度较大,生长有较多喜阳的蕨类植物。

图9 海拔(a)、群落郁闭度(b)、空间距离(c)对乔木层beta多样性的解释度Fig. 9 Venn diagram of altitude (a), crown density (b) and spatial distance (c) on the explanation of beta diversity of the tree layer
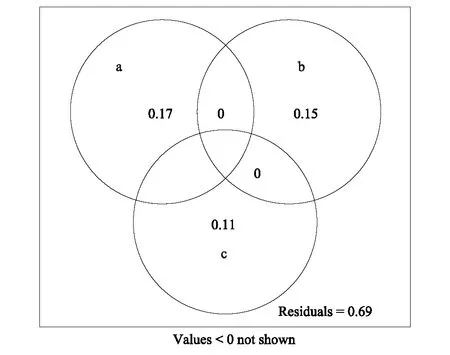
图10 海拔(a)、群落郁闭度(b)、空间距离(c)对灌木层beta多样性的解释度Fig.10 Venn diagram of altitude (a), crown density (b) and spatial distance (c) on the explanation of beta diversity of the shrub layer

图11 海拔(a)、群落郁闭度(b)、空间距离(c)对草本层beta多样性的解释度Fig.11 Venn diagram of altitude (a), crown density (b) and spatial distance (c) on the explanation of beta diversity of the herb layer
本研究还发现海拔对百山祖自然保护区内植物群落beta多样性影响也较大,以往许多研究也表明海拔对于群落物种组成是一个重要的影响因素[34- 38]。通常认为海拔对植物群落产生的作用是间接的,主要是通过对气候、土壤等因素的作用对群落产生影响,一般情况下随着海拔的升高,气温会逐渐下降,而降雨量和相对湿度会随着海拔的升高而增加。百山祖自然保护区分为间断的两块区域,调查样地之间海拔落差可达1000 m以上,植物群落的垂直分布较为明显,不同海拔高度上群落的物种组成、外貌和结构都有差异。
对百山祖自然保护区植物群落相似性有显著作用的第3个因素为群落空间距离。中性理论认为群落中各物种具有相同的竞争能力,不同地点的物种组成存在差异是由于物种扩散能力有限造成,故随着研究地点之间空间距离增加,植物群落的相似性表现出逐渐降低[9]。Tuomisto等人对西亚马逊森林植被差异的研究结果显示,靠风传播孢子的蕨类植物和靠鸟取食浆果传播种子的野牡丹科植物,其分布与空间距离的关系不同,野牡丹科植物受到的扩散限制作用要强于蕨类植物[20]。物种有限的扩散能力会导致无法到达离母树较远的地方,因而随着空间距离的增加,群落相似性逐渐降低。但本研究结果中环境因子也对植物群落组成有显著作用,说明完全中性的群落存在的可能性较低,植物群落的beta多样性更可能是受扩散限制和环境因子共同作用。当然,这里空间距离对beta多样性解释的部分并不可能仅仅由扩散限制造成,其它诸如未观测到的且具有空间结构的环境变量等也可能增加这部分的解释度。
更重要的是,有高达64%的beta多样性格局无法被5个环境因子和群落空间距离解释。产生这一结果的原因可能有很多,例如其他无空间特异性的生物因子(类似随机的二次种子扩散)及在极小空间尺度上变化的非生物环境因子(如pH值的变化等)对beta多样性格局的作用[39]。此外,纯随机过程(例如台风干扰,冰冻灾害等)也可能是一个重要原因[40- 41]。群落学中性理论指出,无论是否存在扩散限制,随机漂变仍是群落构建的重要力量。因此,这些高比例未解释的beta多样性格局可能反映了随机过程在百山祖自然保护区植物群落构建中的重要作用,或者其他未考虑到的生态过程。此外,本研究中并未发现坡向、坡位和坡度对百山祖自然保护区植物群落beta多样性有显著影响,但一般认为坡位和坡度也会影响植物的分布乃至植物群落的组成[42]。有可能它们的作用与随机过程的作用强度相当,这需要进行更多的验证。当然,生境异质性和扩散限制并不是维持植物群落beta多样性唯一的两大因素,其他一些因素,例如种间相互作用、物种库等因素,都可能显著影响植物群落beta多样性格局[43- 44]。
致谢:浙江大学郑朝宗教授在植物分类方面提供了帮助,丽水学院赵鹂教授和谭毓佳、吴香花、杭州师范大学郑毅、赵银佳和杨王伟等,浙江大学美国进修生Lauren Barry、董钢、黄翔参加了野外调查、数据录入工作,特此致谢。
[1] Whittaker R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213- 251.
[2] Kimmins J P. Forest Ecology. New York: Macmillan Publishing Company, 1987.
[3] Condit R, Pitman N, Leigh E G Jr, Chave J, Terborgh J, Foster R B, Núez P, Aguilar S, Valencia R, Villa G, Muller-Landau H C, Losos E, Hubbell S P. Beta-diversity in Tropical forest trees. Science, 2002, 295(5555): 666- 669.
[4] Margules C R, Pressey R L. Systematic conservation planning. Nature, 2000, 405(6783): 243- 253.
[5] Samuel B, James A R, Jose F R. Biogeographic patterns, β-diversity and dominance in the cerrado biome of Brazil. Biodiversity and Conservation, 2004, 13(12): 2295- 2318.
[6] Wiersma Y E, Urban D L. Beta diversity and nature reserve system design in the Yukon, Canada. Conservation Biology, 2005, 19(4): 1262- 1272.
[7] Jacquemyn H, Honnay O, Pailler T. Range size variation, nestedness and species turnover of orchid species along an altitudinal gradient on Réunion Island: implications for conservation. Biological Conservation, 2007, 136(3): 388- 397.
[8] Whittaker R H. Vegetation of the Siskiyou mountains, Oregon and California. Ecological Monographs, 1960, 30(3): 279- 338.
[9] Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton: Princeton University Press, 2001.
[10] Pitman N C A, Terborgh J W, Silman M R, Núez P, Neill D A, Cerón C E, Palacios W A, Aulestia M. Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecology, 2001, 82(8): 2101- 2117.
[11] Leibold M A, Mcpeek M A. Coexistence of the niche and neutral perspectives in community. Ecology, 2006, 87(6): 1399- 1410.
[12] Legendre P, Mi X C, Ren H B, Ma K P, Yu M J, Sun I F, He F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology, 2009, 90(3): 663- 674.
[13] Chen S B, Ouyang Z Y, Xu W H, Xao Y. A review of beta diversity studies. Biodiversity Science, 2010, 18(4): 323- 335.
[14] Tuomisto H, Ruokolainen K, Kalliola R, Linna A, Danjoy W, Rodriguez Z. Dissecting Amazonian biodiversity. Science, 1995, 269(5220): 63- 66.
[15] Nekola J C, White P S. The distance decay of similarity in biogeography and ecology. Journal of Biogeography, 1999, 26(4): 867- 878.
[16] Valencia R, Foster R B, Villa G, Condit R, Svenning J C, Hernández C, Romoleroux K, Losos E, Magård E, Balslev H. Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador. Journal of Ecology, 2004, 92(2): 214- 229.
[17] Seidler T G, Plotkin J B. Seed dispersal and spatial pattern in tropical trees. PloS Biology, 2006, 4(11): 2132- 2137.
[18] Qian H. Beta diversity in relation to dispersal ability for vascular plants in North America. Global Ecology and Biogeography, 2009, 18(3): 327- 332.
[19] Holland J D. Isolating spatial effects on beta diversity to inform forest landscape planning. Landscape Ecology, 2010, 25(9): 1349- 1362.
[20] Tuomisto H, Ruokolainen K, Yli-Halla M. Dispersal, environment, and floristic variation of western Amazonian forests. Science, 2003, 299(5604): 241- 244.
[21] Balvanera P, Lott E, Segura G, Siebe C, Islas A. Patterns ofβ-diversity in a Mexican tropical dry forest. Journal of Vegetation Science, 2002, 13(2): 145- 158.
[22] Chust G, Chave J, Condit R, Aguilar S, Lao S, Pérez R. Determinants and spatial modeling of tree β-diversity in a tropical forest landscape in Panama. Journal of Vegetation Science, 2006, 17(1): 83- 92.
[23] Bin Y, Wang Z G, Wang Z M, Ye W H, Cao H L, Lian J Y. The effects of dispersal limitation and topographic heterogeneity on beta diversity and phylobetadiversity in a subtropical forest. Plant Ecology, 2009, 209(2): 237- 256.
[24] Jin X F, Ding B Y, Zheng C Z, Ye Z L, Chen X R. The floristic analysis of seed plants in Baishanzu Nature Reserve from Zhejiang Province. Acta Botanica Yunnanica, 2004, 26(6): 605- 618.
[25] Xu M, Luo Z R, Yu M J, Ding B Y, Wu Y G. Floristic composition and community structure of mid-montane evergreen broad-leaved forest in north slope of Baishanzu Mountain. Journal of Zhejiang University: Agriculture and Life Sciences, 2007, 33(4): 450- 457.
[26] Chao A, Chazdon R L, Colwell R K, Shen T J. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecology Letters, 2005, 8(2): 148- 159.
[27] Legendre P, Legendre P L F. Numerical Ecology. New York: Elsevier Science, 1998.
[28] Standish R J, Cramer V A, Wild S L, Hobbs R J. Seed dispersal and recruitment limitation are barriers to native recolonization of old-fields in western Australia. Journal of Applied Ecology, 2007, 44(2): 435- 445.
[29] Collins S L, Good R E. The seedling regeneration niche: habitat structure of tree seedlings in an Oak-Pine Forest. Oikos, 1987, 48(1): 89- 98.
[30] Schupp E W. Seed-seedling conflicts, habitat choice, and patterns of plant recruitment. American Journal of Botany, 1995, 82(3): 399- 409.
[31] Sternberg M, Yu S L, Bar P. Soil seed banks, habitat heterogeneity, and regeneration strategies in a Mediterranean coastal sand dune. Israel Journal of Plant Sciences, 2004, 52(3): 213- 221.
[32] Augspurger C K. Seedling survival of tropical tree species: interactions of dispersal distance, light-gaps, and pathogens. Ecology, 1984, 65(6): 1705- 1712.
[33] Denslow J S, Ellison A M, Sanford R E. Treefall gap size effects on above-and below-ground processes in a tropical wet forest. Journal of Ecology, 1998, 86(4): 597- 609.
[34] Shen Z H, Hu H F, Zhou Y, Fang J Y. Altitudinal patterns of plant species diversity on the southern slope of Mt. Shennongjia, Hubei, China. Biodiversity Science, 2004, 12(1): 99- 107.
[35] Zhu B, Chen A P, Liu Z L, Li G Z, Fang J Y. Changes in floristic composition, community structure, and tree species diversity of plant communities along altitudinal gradients on Mt. Mao′er, Guangxi, China. Biodiversity Science, 2004, 12(1): 44- 52.
[36] Zhu B, Chen A P, Liu Z L, Fang J Y. Plant community composition and tree species diversity on eastern and western Nanling Mountains, China. Biodiversity Science, 2004, 12(1): 53- 62.
[37] Tang Z Y, Ke J H. Altitudinal patterns of plant species diversity in Mt. Niubeiliang, Qinling Mountains. Biodiversity Science, 2004, 12(1): 108- 114.
[38] Tang Z Y, Fang J Y, Zhang L. Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains. Biodiversity Science, 2004, 12(1): 115- 122.
[39] Gabriel D, Roschewitz I, Tscharntke T, Thies Carsten. Beta diversity at different spatial scales: plant communities in organic and conventional agriculture. Ecological Applications, 2006, 16(5): 2011- 2021.
[40] Legendre P. Studying beta diversity: ecological variation partitioning by multiple regression and canonical analysis. Journal of Plant Ecology, 2008, 1(1): 3- 8.
[41] Wang Q, Yu D, Li Z Q, Wang L G. The effect of Typhoons on the diversity and distribution pattern of aquatic plants on Hainan Island, south China. Biotropica, 2008, 40(6): 692- 699.
[42] Song Y C. Vegetation Ecology. Shanghai: Press of East China Normal University, 2001: 174- 175.
[43] Caswell H, Cohen J E. Disturbance, interspecific interaction and diversity in metapopulations. Biological Journal of the Linnean Society, 1991, 42(1/2): 193- 218.
[44] Connell J H, Hughes T P, Wallace C C, Tanner J E, Harms K E, Kerr A M. A long-term study of competition and diversity of corals. Ecological Monographs, 2004, 74(2): 179- 210.
参考文献:
[13] 陈圣宾, 欧阳志云, 徐卫华, 肖燚. Beta多样性研究进展. 生物多样性, 2010, 18(4): 323- 335.
[24] 金孝锋, 丁炳扬, 郑朝宗, 叶珍林, 陈小荣. 浙江百山祖自然保护区种子植物区系分析. 云南植物研究, 2004, 26(6): 605- 618.
[25] 徐敏, 骆争荣, 于明坚, 丁炳扬, 吴友贵. 百山祖北坡中山常绿阔叶林的物种组成和群落结构. 浙江大学学报: 农业与生命科学版, 2007, 33(4): 450- 457.
[34] 沈泽昊, 胡会峰, 周宇, 方精云. 神农架南坡植物群落多样性的海拔梯度格局. 生物多样性, 2004, 12(1): 99- 107.
[35] 朱彪, 陈安平, 刘增力, 李光照, 方精云. 广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局. 生物多样性, 2004, 12(1): 44- 52.
[36] 朱彪, 陈安平, 刘增力, 方精云. 南岭东西段植物群落物种组成及其树种多样性垂直格局的比较. 生物多样性, 2004, 12(1): 53- 62.
[37] 唐志尧, 柯金虎. 秦岭牛背梁植物物种多样性垂直分布格局. 生物多样性, 2004, 12(1): 108- 114.
[38] 唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释. 生物多样性, 2004, 12(1): 115- 122.
[42] 宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001: 174- 175.
BetadiversityofplantcommunitiesinBaishanzuNatureReserve
TAN Shanshan1, YE Zhenlin2, YUAN Liubin3, ZHOU Rongfei2, HU Guang1, JIN Xiaofeng4, YU Mingjian1,*
1CollegeofLifeSciences,ZhejiangUniversity,Hangzhou310058,China2ManagementofBaishanzu,Fengyangshan-BaishanzuNationalNatureReserve,Qingyuan323800,China3CollegeofEcology,LishuiUniversity,Lishui323000,China4CollegeofLifeandEnvironmentalScience,HangzhouNormalUniversity,Hangzhou310036,China
In this study, we analyzed the beta diversity pattern of forest plant communities in Baishanzu Nature Reserve by typical quadrat methods. The beta diversity pattern was described by Chao′s index based on 45 20m×20m plots in this area. Correlations between the beta diversity and five environmental factors (coverage of tree layer, elevation, slope grade, slope aspect and slope position) and spatial distances of communities were examined by Mantel test, Distance-based Redundancy analysis and ANOVA. The results showed that, beta diversity increased with spatial distance and environmental difference among plots, but only 36% of its variance can be explained by the joint effects of spatial distance and environmental difference. Specifically, only coverage of tree layer and elevation of the local community significantly affected on beta diversity at Baishanzu Nature Reserve. The impact of coverage of tree layer (explained 20.0% variance of the beta diversity) on beta diversity was also slightly larger than that of elevation (18.0%), and evidently lager than spatial distance (only 9.0%). Our study revealed the pattern of beta diversity and its relationship with environment and spatial distance of plant communities in Baishanzu Nature Reserve, and the results supported the joint effect hypothesis of habitat heterogeneity and dispersal limitation on beta diversity.
Baishanzu Nature Reserve;Beta diversity;environmental factors;spatial distance;forest plant community
中央高校基本科研业务费专项资助项目
2012- 07- 01;
2013- 05- 13
*通讯作者Corresponding author.E-mail: fishmj202@hotmail.com
10.5846/stxb201207010920
谭珊珊,叶珍林,袁留斌,周荣飞,胡广,金孝锋,于明坚.百山祖自然保护区植物群落beta多样性.生态学报,2013,33(21):6944- 6956.
Tan S S, Ye Z L, Yuan L B, Zhou R F, Hu G, Jin X F, Yu M J.Beta diversity of plant communities in Baishanzu Nature Reserve.Acta Ecologica Sinica,2013,33(21):6944- 6956.
猜你喜欢
党政干部论坛(2022年3期)2022-04-20 10:13:34
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
福建林业(2020年5期)2020-03-18 08:23:02
热带林业(2019年4期)2019-03-05 09:53:58
森林工程(2018年3期)2018-06-26 03:40:46
中国环境监察(2016年5期)2016-10-24 05:26:00
现代农业(2016年5期)2016-02-28 18:42:42
水生生物学报(2015年1期)2015-02-28 16:01:05
农民致富之友(2014年7期)2014-10-21 20:06:25
