
世界蜘蛛的分布格局及其多元相似性聚类分析
2013-12-09 06:01:14申效诚张保石刘新涛
生态学报 2013年21期
申效诚,张保石, 张 锋, 刘新涛
(1. 河南省农科院植物保护研究所, 郑州 450002; 2. 保定学院生化系, 保定 071000;3. 河北大学生命科学学院, 保定 071002)
世界蜘蛛的分布格局及其多元相似性聚类分析
申效诚1,*,张保石2, 张 锋3, 刘新涛1
(1. 河南省农科院植物保护研究所, 郑州 450002; 2. 保定学院生化系, 保定 071000;3. 河北大学生命科学学院, 保定 071002)
蜘蛛是一类种类繁多、数量巨大、分布广泛的捕食性生物。至2012年底,全世界共有蜘蛛43678种(包括亚种),隶属于112科3898属。科、属、种3个分类阶元的分布域非常悬殊,90%的种分布在一个界内,90%的科是跨界分布或全球分布。按行政区域,亚洲种类较多,欧洲较少,南极洲没有蜘蛛记录;按动物地理区域,古北界和新热带界较多,新北界较少。根据地理条件、生态条件和蜘蛛的分布状况,将全球陆地分为53个基础地理单元,用申效诚等新近提出的相似性通用公式和据此创立的多元相似性聚类分析方法,分别对属、种两级的分布进行分析,得到两个聚类结构相同、聚类关系合理的支序图,而且属级的支序图层次更为分明,在相似性水平为0.32时,53个基础地理单元聚为17个小单元群,在0,22水平上,又聚为8个大单元群。这些大、小单元群的组成单元地域相邻相连,生态条件相对一致,可以作为蜘蛛地理区划的界、亚界两个层级。和华莱士主要以哺乳动物建立的世界动物地理区划相比,主要差异是:1)古北界东、西两部分差异显著,可分设两界;2)新西兰和澳洲大陆相似性较低,可单独设界;3)新几内亚和太平洋岛屿与澳洲大陆的关系远于和东洋界的关系,华莱士线在两界间的作用似乎不存在; 4)新热带界的中美地区似乎属于新北界更为合适,并由此产生了南北美洲间的紧密联系;5)新北界与古北界的相似性关系弱于与新热带界的关系,全北界的概念几近消失。前两点差异可以从地球板块构造的变动得到解释,第3、5个差异已在植物和其它生物类群得到佐证,第4个差异尚不稳定,需要更多类群的比较与分析。使用多元相似性聚类分析方法对于如此典型的点状分布的生物类群和如此海量的数据,能够得到如此精细的,既符合地理学、统计学的逻辑,又符合生物学、生态学逻辑的定量分析结果,这在国内外都是首次成功尝试,其简便性和合理性将会促使在其它类群中的应用。
蜘蛛;分布;生物地理;多元相似性聚类分析
蜘蛛是一类种类繁多、数量巨大、分布广泛的捕食性生物,对抑制农林害虫发生、维持生态平衡等方面具有重要作用。对于蜘蛛的研究,西方发展较早,从林奈于1758年建立分类系统以来,已有200多年的历史,有少数经济发展较快、研究较早的国家和地区的蜘蛛资源已基本查清,如北美、西欧和日本等。到了19世纪中期后,蜘蛛研究得到了迅速发展。截止2011年12月31日,全世界共有蜘蛛110科3859属42751种(亚种)。由于全球地理条件、生态条件各异,蜘蛛的分布极不均匀。为了比较世界蜘蛛分布的差异及相互联系的紧密程度,应用作者近年提出的多元相似性系数通用公式[1]和多元相似性聚类分析法[2]对世界不同基础地理单元进行比较,以期为世界蜘蛛地理区划提出定量方案,并和世界高等动物地理区划进行比较与分析。
1 材料和方法
1.1 供分析的蜘蛛种类
蜘蛛的种类和分布资料来源于蜘蛛专业网站、蜘蛛专著、分类区系论文[3- 15]和长期采集调查。汇总全世界的蜘蛛种类及其分布地,剔除没有收集到分布地或分布地存疑的种类,共计110科3823属41992种,占2012年底全世界蜘蛛种类的96.1%,属的98.1%,科的98.2%。
1.2 基础地理单元的划分
根据全球地理条件、生态条件和蜘蛛分布状况,将全球陆地分为53个基础地理单元。各基础地理单元的地理范围见表1。
1.3 数据库的建立
将各个种类的行政区域分布资料转换为基础地理单元分布记录,用微软Access构建数据库,有分布记“1”,无分布不记。并汇总各属、各科的分布信息,以便分别进行各种统计查询和进行属、种两个层次的相似性分析。各基础地理单元的蜘蛛种类见表1。
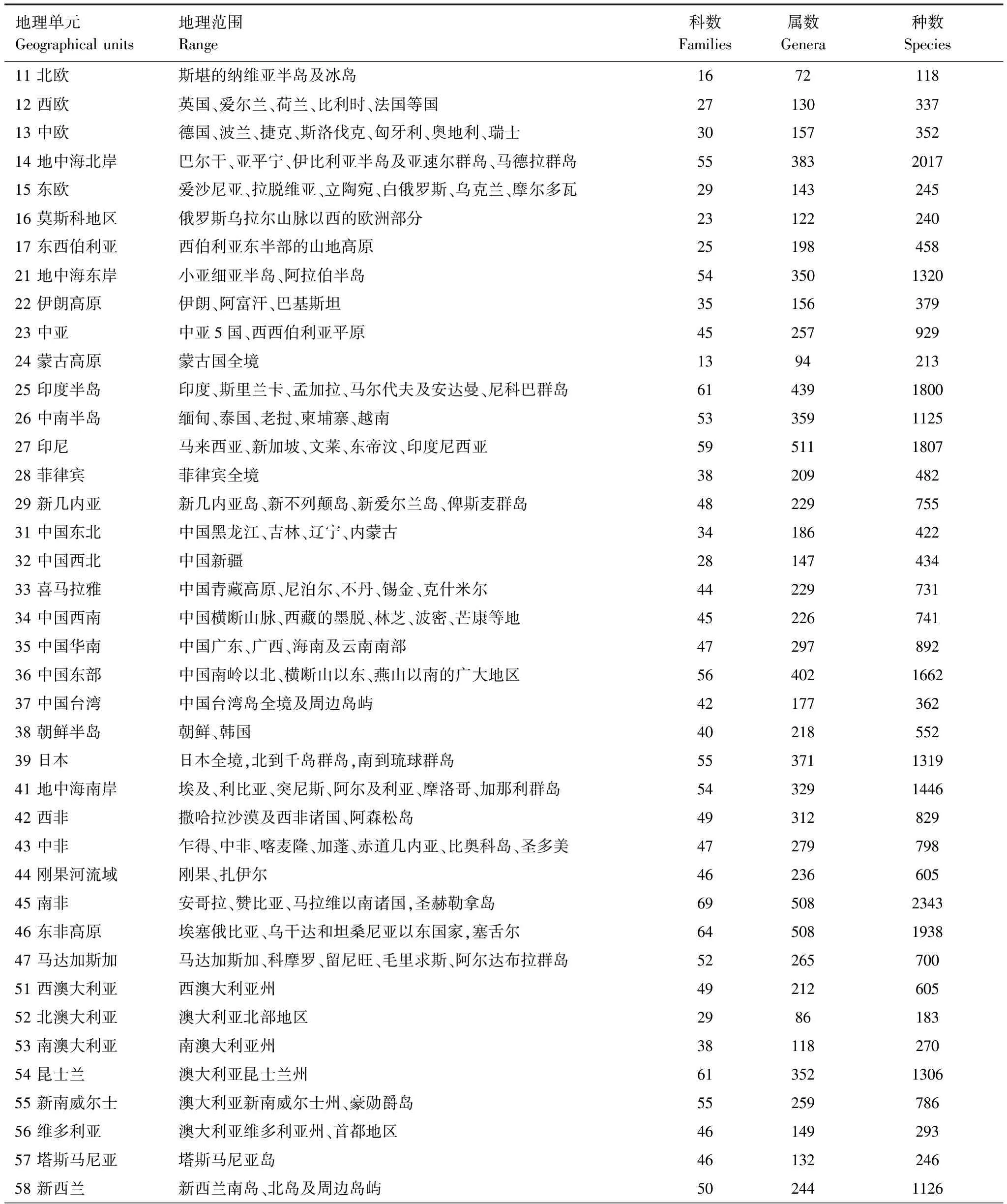
表1 世界蜘蛛基础地理单元的地理范围和蜘蛛种类
续表
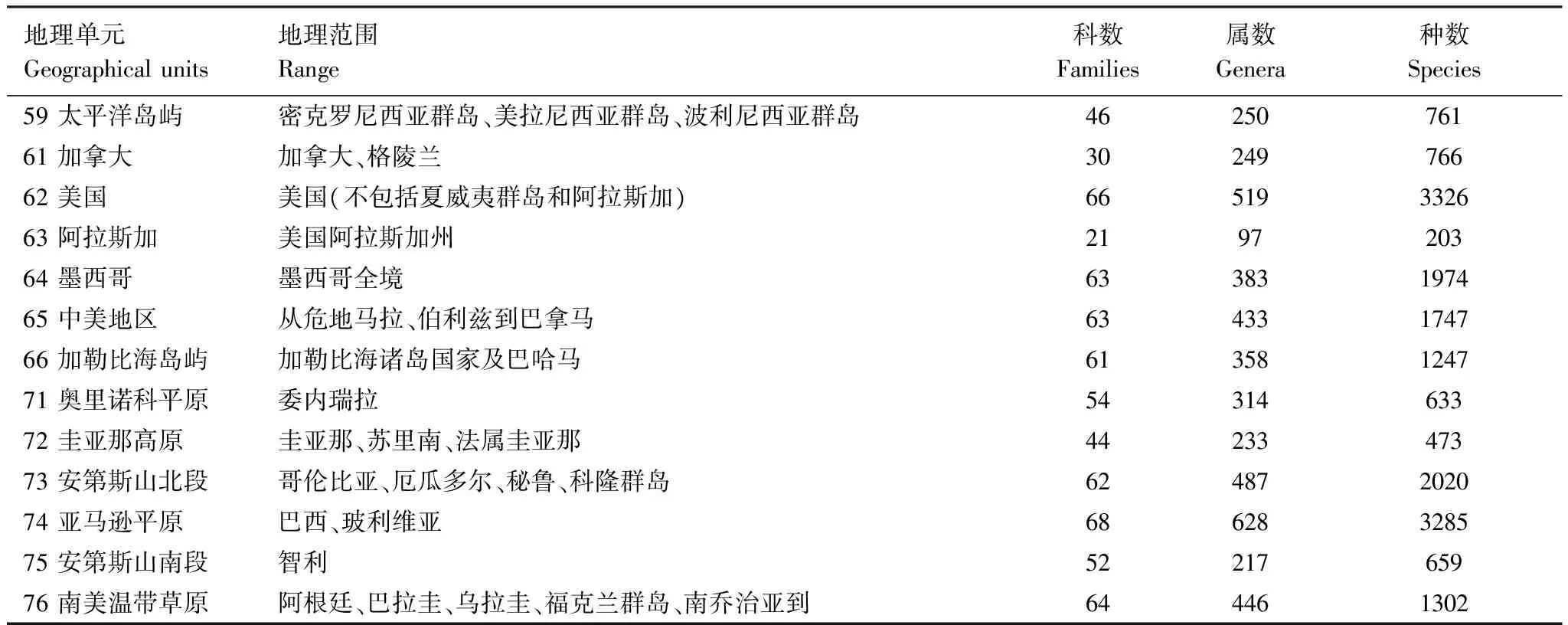
地理单元Geographicalunits地理范围Range科数Families属数Genera种数Species59太平洋岛屿密克罗尼西亚群岛、美拉尼西亚群岛、波利尼西亚群岛4625076161加拿大加拿大、格陵兰3024976662美国美国(不包括夏威夷群岛和阿拉斯加)66519332663阿拉斯加美国阿拉斯加州219720364墨西哥墨西哥全境63383197465中美地区从危地马拉、伯利兹到巴拿马63433174766加勒比海岛屿加勒比海诸岛国家及巴哈马61358124771奥里诺科平原委内瑞拉5431463372圭亚那高原圭亚那、苏里南、法属圭亚那4423347373安第斯山北段哥伦比亚、厄瓜多尔、秘鲁、科隆群岛62487202074亚马逊平原巴西、玻利维亚68628328575安第斯山南段智利5221765976南美温带草原阿根廷、巴拉圭、乌拉圭、福克兰群岛、南乔治亚到644461302
1.4 多元相似性聚类分析(MSCA)
首先,用Jaccard的二元相似性系数计算公式算出两两单元间的相似性系数,从最大的相似性系数开始,用申效诚等的多元相似性公式,计算3个单元、多个单元、以至全部53个单元的相似性系数,作出支序图。
Jaccard的公式:
SI=C/(A+B-C)
式中,SI是两地理单元间的相似性系数,A、B分别是两单元的物种数,C是两单元的共有物种数。
申效诚等的SGF:
SIn= ∑Hi/nSn= ∑(Si-Ti)/nSn
式中,SIn是n个地理单元间的相似性系数,Si,Hi,Ti分别是i单元的种类数、共有种类数、独有种类数,满足Hi=Si-Ti,Sn是n个单元的总种类数。因此,公式可表述为:多个地理单元的相似性系数是各自共有种类的平均数占总种类数的比例。
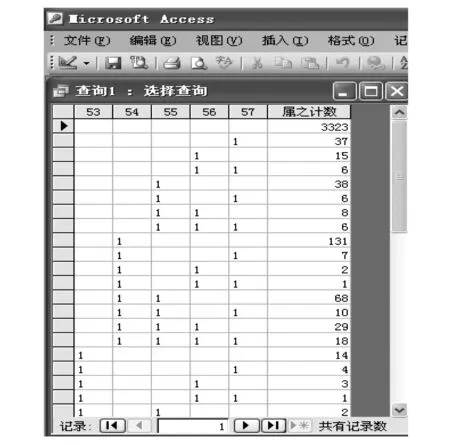
图1 5个地理单元的蜘蛛属的分布查询表 Fig.1 The distribution inquiry table of spider genera for 5 geographical units
上述两个公式的不同在于,Jaccard的公式是申效诚等的公式在n为2时的一个特例,后者可代替前者,前者不能代替后者。
计算多个地区的相似性系数所需的参数都可以从数据库的查询表中得到。如要计算第53—57号5个地理单元的蜘蛛属相似性系数,直接建立查询表如图1。
由表1已知,数据库的总属数为3823属,5个地理单元的蜘蛛属数分别为118,352,259,149,132,其和为1010,图1第1行的3323属是5个地理单元都没有的属,这些单元的总属数为3823-3323=500属;图中各列第1条记录的属数是各单元的独有属数,分别是14,131,38,15,37,其和为235,5个地理单元的相似性系数为(1010-235)/5×500=0.31。按左上方的视图键后,即可返回查询设计视图,随意更改、增添或删除小区,直到所有参与小区聚类完毕。新的MSCA法与传统的相似性聚类分析法的根本区别在于多元相似性系数的直接计算和摒弃计算过程中的合并环节。
2 结果与分析
2.1 蜘蛛的分布格局
世界陆地除南极洲没有蜘蛛记录外,6个洲中,亚洲蜘蛛种类最多,欧洲最少;6个动物地理界中,古北界和新热带界最多,新北界最少(表2)。

表2 各洲、各界的蜘蛛种类
蜘蛛的集中产地有亚马逊河流域、东南非高原、地中海和东亚等地,蜘蛛分布最多的国家有中国、美国、澳大利亚、巴西等国,面积很小的岛屿蜘蛛种类也非常丰富(表3)。
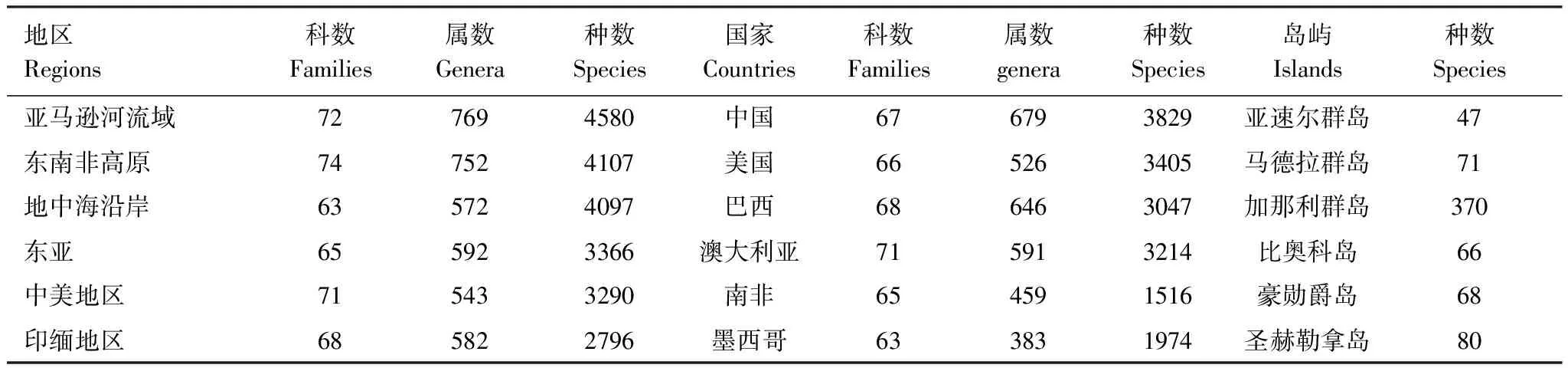
表3 蜘蛛种类最丰富的地区、国家和岛屿
中国是世界上蜘蛛种类最多的国家,占世界蜘蛛种类的9.1%,其中古北种类650种,占中国种类的17.7%,东洋种类312种,占中国种类的8.1%,跨古北、东洋两界的种类57种,占中国种类的1.5%;中日种类228种,占中国种类的6.0%,其余2582种是中国特有种类,占中国种类的67.4%。
2.1.2 各分类阶元分布域的宽度
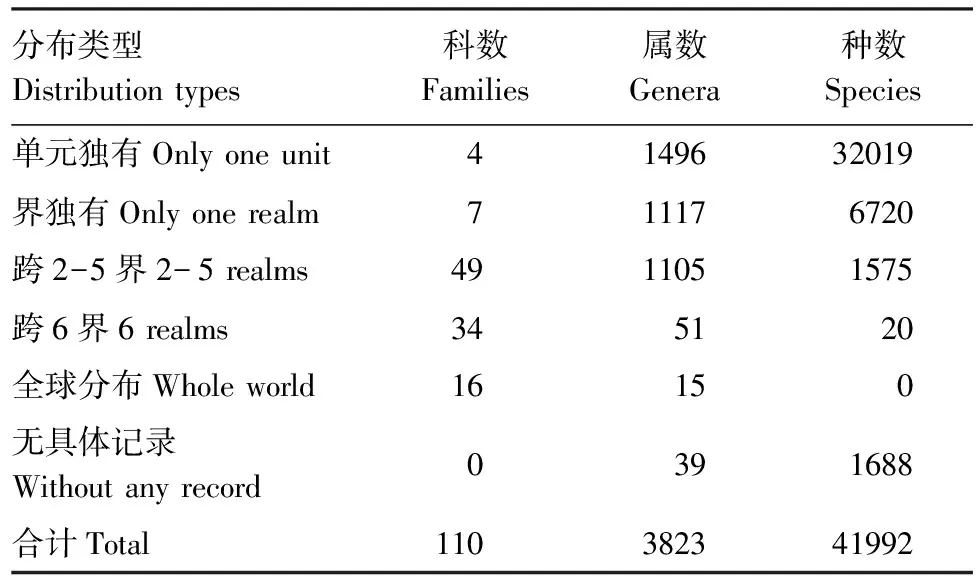
表4 不同阶元的分布状况
蜘蛛没有翅,自然的扩散迁移能力远不如昆虫,因此,种的分布域非常狭窄,约3/4的种类局限在1个地理单元内,还有1/6的种类局限在1个动物界内,只有1500余种是跨界分布,而且多是跨2、3界,能够跨6界分布的只有20种(表4)。平均每种分布1.2个地理单元。
蜘蛛属的分布域比种大为扩展,局限在1个地理单元内的属不到40%,单界分布的不到30%,有超过30%的属跨界分布,其中跨6界的属有66个,更有15属是绝大部分单元都有分布的可称之为全球性分布的属。平均每属分布3.8个地理单元。
蜘蛛的科级分布域极为宽阔,45%的科跨6界分布,其中卵形蛛科Oonopidae, 球蛛科Theridiidae, 皿蛛科Linyphiidae, 肖蛸科Tetragnathidae, 圆蛛科Araneidae, 狼蛛科Lycosidae, 漏斗蛛科Agelenidae, 栅蛛科Hahniidae, 卷叶蛛科Dictynidae, 米图蛛科Miturgidae, 管巢蛛科Clubionidae, 圆颚蛛科Corinnidae, 拟平腹蛛科Zodariidae, 平腹蛛科Gnaphosidae, 蟹蛛科Thomisidae, 跳蛛科Salticidae等16科几乎分布于所有地理单元;有30%的科为跨2—5界分布,只有7个科分别局限在澳洲界或新北界内,而Penestomidae和Chummidae共2科2属11种仅局限在南非地理单元,Huttoniidae只有1属1种,仅局限在新西兰地理单元,2008年建立的新科Sinopimoidae仅含1属1种,是在中国西南地理单元采到的。平均每科分布22.3个地理单元。
2.2 种、属的多元相似性聚类分析结果
蜘蛛种和属的MSCA结果分别为图2和图3。
比较图2和图3,所聚成的群数和各群的组成完全相同,群内各单元的聚类顺序也大体相同,不同的是属级阶元的聚类,相似性水平要比种级阶元高,而且能够提供更多的聚类信息。
在图2的0.025的相似性水平上和图3的0.17水平上,53个地理单元聚成了相同的5个大单元群,A、B、C、D分别相当于世界动物区划中的古北界、东洋界、非洲界和澳洲界,E群则是南北美洲。
在图3的0.22水平上,聚成8个单元群,即A群分为东、西两部分,D群中新西兰与澳大利亚分开,E群分为南、北两部分。
在图3的0.32水平上,聚为17个小单元群,基本相当于各界下的亚界,1—4小群相当于古北界的欧洲平原亚界、地中海亚界(包括中亚亚界)、西伯利亚亚界和东亚亚界;5小群相当于东洋界,6、7小群原属于澳洲界,但和澳洲界关系较远,接近于东洋界;8、9、10小群可作为非洲平原丘陵亚界、东南非高原亚界和马达加斯加亚界;11、12小群分别是西澳、东澳亚界,13小群即新西兰界;14小群相当于新北界,15小群中美地区原属于新热带界,现和新北界关系密切,16、17小群是南美洲的平原高原和智利山地两部分。
蜘蛛的上述聚类结果和哺乳动物的地理区划的差异,是真实和客观存在的。蜘蛛是节肢动物门的一个成员,进化历史已有3亿多年,比只有6500万年的哺乳动物经历过多得多的历史事件;蜘蛛是变温动物,受环境影响比哺乳动物大得多;蜘蛛多以昆虫为食,昆虫又大多以植物为食,食物链的关系决定蜘蛛在哺乳动物、植物和昆虫之间,会与昆虫最密切,再与植物为接近,和哺乳动物最远。况且,目前仍然沿用的19世纪对哺乳动物和有花植物的阐释[16]都是定性研究的结果,各大地理区域间的关系还必须接受定量分析的检验,因此,如此的差异是合乎生物学逻辑和统计学逻辑的,如果没有差异,倒是有违自然规律。
进一步剖析这些差别的原因,蜘蛛在古北界东、西两部分的相似性存在较大差异,可能是两个地理区域之间在2.1亿年前到6500万年前,由鄂毕海相隔将近1.5亿年,占蜘蛛进化历史的将近一半,这应该在蜘蛛的区系构成上留下痕迹;新西兰和澳大利亚大陆虽然地理距离较近但生物关系较远,是由于它们先后脱离冈瓦纳大陆以后,一直没有接触的历史,较大的生物区系差异早已是生物地理学者的共识,只是没人定量地进行比较过;新几内亚及大洋洲岛屿与东洋界保持较近的关系而远离澳大利亚大陆,早已在植物界是不争的事实,也必然影响到昆虫和蜘蛛,最近,蚁科昆虫的分析也予以证明[17];南美洲和北美洲的密切关系也已在哺乳动物和有花植物中得到证实[18]。
3 结论与讨论
蜘蛛是种类繁多、全世界广泛分布、但属种的分布又非常狭窄的生物类群,特有性突出,各地分布差异悬殊,亚洲和古北界、新热带界种类较多,欧洲和新北界较少;亚马逊河流域、东南非高原、地中海沿岸以及中国、美国、澳大利亚、巴西是蜘蛛的集中产地。
对蜘蛛的属、种两个阶元进行多元相似性聚类分析结果,与世界哺乳动物区划6个界的定性划分相比,各动物界之内和之间的定量关系有所差异。这是生物进化与地球板块结构共同作用的结果,也是低等动物与高等动物分布格局不同的具体体现。诚然,这仅仅是一个蜘蛛目的数据,但至少已表明不能把华莱士主要用哺乳动物做出的世界动物地理区划来外延到整个动物界。若要对包括低等动物在内的世界动物分布格局进行定量性的评价,尚需更多的动物类群参与,但这已是一个良好的开端。

图2 蜘蛛种级阶元的聚类支序图Fig.2 The MSCA tree for spider species
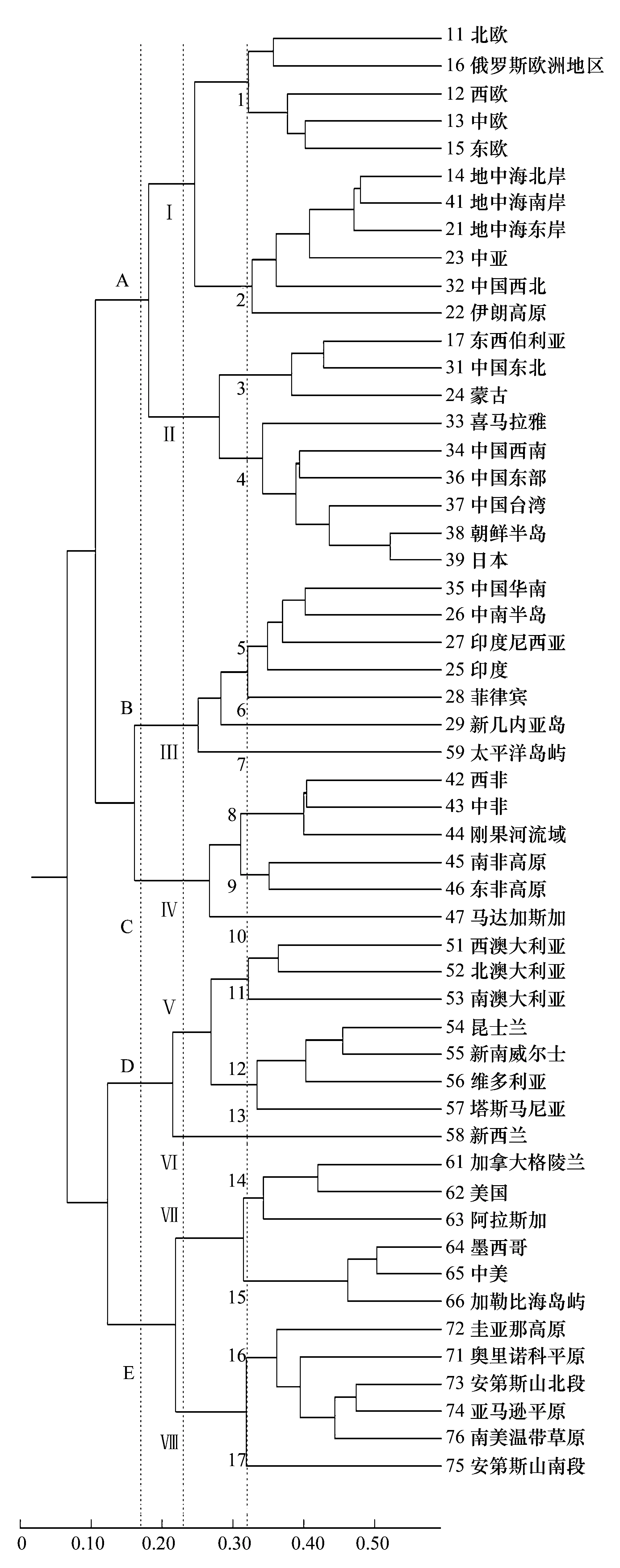
图3 蜘蛛属级阶元的聚类支序图Fig.3 The MSCA tree for spider genera
本研究所使用的多元相似性聚类分析技术是我们近年创建的新方法,对于如此典型的点状分布的生物类群和如此海量的数据,能够得到如此精细的,既符合地理学、统计学的逻辑,又符合生物学、生态学逻辑的定量分析结果,在世界上尚难见到。之前的诸多以“合并降阶”为特征的传统聚类分析法均难以获得如此的结果。这预示着生物地理研究从定性研究进入定量研究将会加速。
[1] Shen X C, Wang A P. A simple formula for multivariate similarity coefficient and its contribution rate in analysis of insect fauna. Journal of Henan Agricultural Sciences, 2008, (7): 67- 69.
[2] Shen X C, Ren Y D, Wang A P, Zhang S J. A multivariate similarity clustering analysis for geographical distribution of insects, spiders and mites in Henan province. Acta Ecologica Sinica, 2010, 30(16): 4416- 4426.
[3] Platnick N I. The World Spider Catalog, Version 13.5. 2013, [2013- 05- 06]. http://research.amnh.org/iz/spiders/catalog/ INTRO1.html
[4] Song D X, Zhu M S, Chen J. The Spiders of China. Shijiazhuang: Hebei Science and Technology Publishing House, 1999: 32- 564.
[5] Yin C M, Wang J F, Zhu M S, Xie L P, Peng X J, Bao Y H. Fauna Sinica, Arachnida Araneae: Araneidae. Beijing: Science Press, 1997: 57- 403.
[6] Zhu M S. Fauna Sinica Arachnida Araneae Theridiidae. Beijing: Science Press, 1998: 17- 348.
[7] Song D X, Zhu M S. Fauna Sinica Arachnida: Araneae Thomisidae, Philodromidae. Beijing: Science Press, 1997: 19- 214.
[8] Zhu M S, Song D X, Zhang J X. Fauna Sinica: Invertebrata, Volume 35: Arachnida Araneae Tetragnathidae. Beijing: Science Press, 2003: 41- 314.
[9] Chen H M, Jia Q, Wang S J. A revision of the genusQianleptoneta(Araneae: Leptonetidae). Journal of Natural History, 2010, 44(47/48): 2873- 2915.
[10] Han G X, Zhu M S. Taxonomy and biogeography of the spider genusEriovixia(Araneae: Araneidae) from Hainan Island, China. Journal of Natural History, 2010, 44(43/44): 2609- 2635.
[11] Lin Y C, Li S Q. Long-legged cave spiders (Araneae: Telemidae) from Yunnan-Guizhou Plateau, southwestern China. Zootaxa 2010, 2445: 1- 34.
[12] Lin Y C, Li S Q. Leptonetid spiders from caves of the Yunnan-Guizhou Plateau, China (Araneae: Leptonetidae). Zootaxa, 2010, 2587: 1- 93.
[13] Song Y J, Li S Q. The spider generaAraeoncusSimon, 1884 andDiplocephalusBertkau, 1883 (Arachnida, Araneae, Linyphiidae) of China. Zoosystema, 2010, 32(1): 117- 137.
[14] Zhang B S, Zhu M S. Two new species of the genusStorenomorphaSimon from China (Araneae: Zodariidae). Acta Arachnologica, 2009, 58(2): 91- 95.
[15] Zhang F, Zhu M S. A review of the genusPholcus(Araneae: Pholcidae) from China. Zootaxa, 2009, 2037: 1- 114.
[16] Cox C B, Moore P D. Biogeography: An Ecological and Evolutionary Approach. 7th ed. Oxford: Blackwell Publishing Ltd, 2005.
[17] Bolton B. A New General Catalogue of the Ants of the World. Cambridge: Harvard University Press, 1995.
[18] Heywood V H. Flowering Plants of the World. Oxford: Oxford University Press, 1978.
参考文献:
[1] 申效诚, 王爱萍. 昆虫区系多元相似性的简便计算公式及其贡献率. 河南农业科学, 2008, (7): 67- 69.
[2] 申效诚, 任应党, 王爱萍, 张书杰. 河南昆虫、蜘蛛、蜱螨地理分布的多元相似性聚类分析. 生态学报, 2010, 30(16): 4416- 4426.
[5] 尹长民, 王家福, 朱明生, 谢莉萍, 彭贤锦, 鲍幼惠. 中国动物志 蛛形纲 蜘蛛目 园蛛科. 北京: 科学出版社, 1997: 57- 403.
[6] 朱明生. 中国动物志 蛛形纲 蜘蛛目 球蛛科. 北京: 科学出版社, 1998: 17- 348.
[7] 宋大祥, 朱明生. 中国动物志 蛛形纲 蜘蛛目 蟹蛛科 逍遥蛛科. 北京: 科学出版社, 1997: 19- 214.
[8] 朱明生, 宋大祥, 张俊霞. 中国动物志 无脊椎动物 第三十五卷 蛛形纲 蜘蛛目 肖蛸科. 北京: 科学出版社, 2003: 41- 314.
Worldwidedistributionandmultivariatesimilarityclusteringanalysisofspiders
SHEN Xiaocheng1,*,ZHANG Baoshi2, ZHANG Feng3, LIU Xintao1
1InstituteofPlantProtection,HenanAcademyofAgriculturalSciences,Zhengzhou450002,China2DepartmentofBiochemistry,BaodingUniversity,Baoding071000,China3CollegeofLifeSciences,HebeiUniversity,Baoding071002,China
Spiders have the greatest number of individuals and species, and the widest distribution, of any predacious animals. As of December 31, 2012, a total of 43,678 spider species and subspecies belonging to 3898 genera and 112families have been reported worldwide. With respect to the three categories of species, genus, and family, distribution areas differ greatly: 90% of species are limited to a single realm, whereas 90% of families are found in at least two realms, with many distributed worldwide. Continental distributions also vary. Asia has the greatest number of spider species, with a smaller number in Europe and none in Antarctica. The greatest number of species is found in Palaearctic and Neotropical realms, the fewest in the Nearctic realm. In this study, we divided world land areas into 53 basic geographic units based on geographical conditions, ecological conditions, and distribution patterns of all known spiders. We then analyzed species and genus distributions using a similarity general formula (SGF) and multivariate similarity clustering analysis (MSCA). In the resulting genus-based tree, all basic geographic units are clustered into 8 groups and 17 subgroups at similarity levels of 0.22 and 0.32, respectively. The eight groups are designated as East Palaearctic, West Palaearctic, Oriental, Afrotropical, Australian, New Zealandian, Nearctic, and Neotropical kingdoms. The 17 subgroups, or subkingdoms, are distributed as follows: East Palaearctic kingdom (Siberian and East Asian subkingdoms), West Palaearctic kingdom (European and Central Asian subkingdoms), Oriental kingdom (Southeast Asian, New Guinea, and Pacific Ocean Islands subkingdoms), Afrotropical kingdom (West African, Southeast African, and Madagascar subkingdoms), Australian kingdom (East Australian and West Australian subkingdoms), New Zealandian kingdom (New Zealandian subkingdom), Nearctic kingdom (North American and Central American subkingdoms), and Neotropical kingdom (Amazon and Andes subkingdoms). Spider geographical divisions differ from those observed for mammals in several ways: 1) the Palaearctic realm is easily divided into eastern and western components; 2) there is a low similarity level between New Zealand and the Australian mainland, such that New Zealand can be considered to constitute its own kingdom; 3) New Guinea and Pacific Islands subkingdoms are more similar to the Oriental realm than to the Australian mainland, with no evident division between the two realms corresponding to Wallace′s line; 4) Central America is more appropriately classified as belonging to the Nearctic realm; and 5) the Nearctic realm is more strongly related to the Neotropical realm than to the Palaearctic realm. The first and second observed differences can be explained by earth plate tectonics and differing evolutionary histories of spiders and mammals. The third and fifth geographical distributional patterns by which spiders differ from mammals are similar to those uncovered from biogeographical studies of plants and other groups. Analyses with additional examples are needed to elucidate the reason for the fourth difference. The application in this study of SGF and MSCA—techniques developed in recent years by the first author—represents the first reported quantitative analysis of typical point-forms and vast amounts of distribution information to successfully generate results that are statistically, geographically, ecologically, and biologically logical. This approach should be applicable to additional animal groups.
spider; distribution; biogeography; multivariate similarity clustering analysis
河南省基础和前沿技术基金资助项目(082300430370)
2012- 07- 08;
2013- 05- 13
*通讯作者Corresponding author.E-mail: shenxiaoc@126.com
10.5846/stxb201207080951
申效诚,张保石, 张锋, 刘新涛.世界蜘蛛的分布格局及其多元相似性聚类分析.生态学报,2013,33(21):6795- 6802.
Shen X C,Zhang B S, Zhang F, Liu X T.Worldwide distribution and multivariate similarity clustering analysis of spiders.Acta Ecologica Sinica,2013,33(21):6795- 6802.
猜你喜欢
数学物理学报(2022年5期)2022-10-09 08:56:44
河北画报(2020年8期)2020-10-27 02:54:20
收藏界(2018年1期)2018-10-10 05:23:08
创新作文(小学版)(2018年31期)2018-05-16 03:00:30
小朋友·快乐手工(2018年3期)2018-04-22 11:48:52
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
小学阅读指南·低年级版(2017年6期)2017-06-12 08:22:47
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:51
小学阅读指南·低年级版(2016年6期)2016-05-14 21:39:20
小朋友·快乐手工(2015年1期)2015-03-13 00:05:24
