
贝壳砂生境酸枣叶片光合生理参数的水分响应特征
2013-12-09 06:30王荣荣夏江宝杨吉华刘京涛赵艳云孙景宽
生态学报 2013年19期
王荣荣,夏江宝,杨吉华,刘京涛,赵艳云,孙景宽
(1. 山东农业大学林学院,泰安 271018; 2. 滨州学院山东省黄河三角洲生态环境重点实验室,滨州 256603)
贝壳砂生境酸枣叶片光合生理参数的水分响应特征
王荣荣1,2,夏江宝2,*,杨吉华1,刘京涛2,赵艳云2,孙景宽2
(1. 山东农业大学林学院,泰安 271018; 2. 滨州学院山东省黄河三角洲生态环境重点实验室,滨州 256603)
以黄河三角洲贝壳堤岛优势灌木酸枣(Ziziphusjujubavar.spinosus)为试验材料,模拟贝壳砂生境系列水分条件,测定分析酸枣叶片在系列水分梯度下的光合参数光响应及叶绿素荧光参数,阐明酸枣主要光合生理参数的水分响应特征。结果表明:(1) 酸枣叶片净光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)随土壤水分的增多先增大后减小,Tr对土壤相对含水量(Wr)的敏感程度大于Pn,因而WUE维持在较高水平。(2) 酸枣叶片Pn下降的原因在Wrgt;25%时以气孔限制为主;Wrlt;25%时以非气孔限制为主,光合机构受到不易逆转的破坏。(3) 酸枣叶片最大净光合速率、暗呼吸速率、表观量子效率(AQY)和光饱和点随土壤水分的增多先增大后减小,光补偿点、光抑制项(β)和光饱和项则先减小后增大。(4)在Wr为80%时,酸枣叶片PSⅡ反应中心的光化学转化效率最高。当Wrlt;30%时,AQY和潜在光化学效率迅速减小,β迅速增大,酸枣光抑制明显。当Wrlt;25%时,非光化学淬灭系数迅速减小,初始荧光迅速增大,酸枣PSⅡ受到不可逆的破坏。(5)Wr在11%—25%内为低产低效水,Wr在25%—58%内为中产中效水,Wr在58%—80%内为高产高效水,Wr在80%—95%内为中产低效水。贝壳砂生境酸枣叶片的光合作用对水分逆境具有较强的生理适应性和可塑性,在Wr为58%—80%内,酸枣光合生理活性较高,利于酸枣苗木生长。
贝壳堤岛;气体交换参数;叶绿素荧光参数;光合生产力;水分利用效率;阈值效应
光合作用是植物最重要的生理过程,光合效率水平是植物生产力和作物产量高低的决定性因素[1]。土壤水分是影响植物光合生理过程的重要生态因子,对植物的光合作用、水分代谢及物质转运等生理活动产生重要作用[2]。近年来,随着淡水资源的紧缺和干旱胁迫的加剧,干旱缺水成为制约植被恢复与农林业发展的关键生态因子[3],如何在保证植物维持较高或者中等生产力水平的前提下,提高植物对土壤水分的利用效率成为农林业建设亟需解决的核心问题[4]。目前对植物抗旱光合生理特性的研究主要集中在植物叶片气体交换参数和叶绿素荧光参数等主要光合生理参数对土壤水分变化的响应规律和作用机理方面[5- 6],大多局限于4—8个水分梯度下的试验结果[7- 8],缺乏土壤水分连续变化中的观测研究,从而导致植物光合生理活动对不同水分条件的响应机制及两者的定量关系等问题还不十分清楚。
贝壳堤由生活在潮间带的贝类死亡之后的壳体及其碎屑,通过波浪搬运在高潮线附近堆积而成,是淤泥质或粉砂质海岸所特有的滩脊类型,生态系统较为独特。黄河三角洲贝壳堤岛滩脊地带海拔相对较高,地下水位较低,贝壳砂土壤孔隙度大、粗砂粒含量最高,涵蓄降水能力差,加之该区域蒸降比较大,季节性缺水严重,从而导致该立地条件下植被生长受到干旱缺水的严重限制,植被类型以旱生的灌木和草本植物为主,生态系统脆弱。酸枣(Ziziphusjujubavar.spinosus)是贝壳堤岛滩脊地带的建群种,具有较强的防风固沙和保持水土的功能,是黄河三角洲贝壳堤岛植被恢复与生态重建的优选树种。以往对酸枣的研究主要集中在药用和经济价值[9]、培植技术[10]以及干旱或复水条件下酸枣幼苗生长、生理和生化特性[11- 12]等方面,而贝壳砂生境酸枣叶片光合生理参数与土壤水分的定量关系及响应过程尚不明确,以至于现有的树木抗旱生理研究在应用于贝壳堤岛的适地适树及物种配置中受到较大限制。鉴于此,本研究通过模拟贝壳砂生境系列水分条件,测定分析酸枣叶片主要光合参数的光响应及叶绿素荧光参数,探讨酸枣叶片光合特性对土壤水分的响应规律,阐明贝壳砂生境酸枣生长适宜的水分条件,以期为黄河三角洲贝壳堤岛酸枣的立地选择及水分栽植管理提供理论依据和技术参考。
1 材料与方法
1.1 试验材料
本试验在山东省黄河三角洲生态环境重点实验室的智能科研温室内进行,温室内的光强约为外界自然光强的85%,温度为18—30 ℃,CO2浓度为345—365 μmol/mol,空气相对湿度为41%—65%。盆栽试验土壤于2011年10月取自黄河三角洲贝壳堤岛滩脊地带酸枣群落的贝壳砂,过筛去除直径大于2 mm的砂粒,土壤田间持水量为24%,土壤容重为1.3 g/cm3。以贝壳堤建群种3年生酸枣苗木为试验材料,植株平均地径为0.8 cm,平均树高为0.7 m。于2012年3月15日采用自制塑料箱体(长80 cm×宽40 cm×高80 cm)进行苗木栽植,以避免栽植容器影响酸枣根系生长,共栽植9盆,每盆1株,置温室中正常培植管理。试验植株正常生长121 d后,选取生长健壮、长势基本一致的5株苗木作为观测样株,于2012年7月14日起进行酸枣叶片光合参数的光响应过程及叶绿素荧光参数的测定。
1.2 系列水分梯度的设计
采用人工给水和自然耗水相结合的方法获取系列土壤水分梯度。2012年7月12日和13日,对5盆观测样株的土壤充分供水使之饱和,7月14日开始停止供水,土壤水分通过植株蒸腾耗水逐渐减少,同时将土壤表面覆塑料薄膜以防止土壤水分蒸发,减缓土壤水分下降速度,于每个晴朗天气(每隔1—2d)采用烘干法测定5盆土壤的重量含水量,每盆3个重复,8月11日测定结束。根据重量含水量和田间持水量的比值计算土壤相对含水量(Wr),求取Wr平均值,获得Wr由95%降至11%的18个水分梯度。
1.3 测定内容与方法
在测定每个土壤水分梯度的当天9:00—11:30,从每个观测样株中选取中上部健康、成熟叶3片,利用CIRAS- 2型便携式光合仪(PP Systems,USA)对酸枣叶片光合参数的光响应过程进行测定。利用人工光源将光合有效辐射(PAR, μmol·m-2·s-1)控制在由1600 μmol·m-2·s-1降至20 μmol·m-2·s-1的13个梯度内,每个光强控制120 s,3次重复读数。通过测定和计算,得到酸枣叶片的净光合速率(Pn, μmol·m-2·s-1)、蒸腾速率(Tr, mmol·m-2·s-1)、水分利用效率(WUE=Pn/Tr, μmol/mmol)[13]、气孔导度(Gs, mmol·m-2·s-1)、胞间CO2浓度(Ci, μmol/mol)和气孔限制值(Ls=1-Ci/Ca,其中,Ca为大气CO2浓度)[14]等主要光合参数的光响应过程。
利用FMS- 2型便携脉冲调制式荧光仪(Hansatech, UK)对酸枣叶片的叶绿素荧光参数进行测定,与光合参数光响应测定同步进行。每株选取中上部健康、成熟叶3片,叶片暗适应1 h后测定初始荧光(Fo)和黑暗中最大荧光(Fm),待叶片在自然光下活化1 h后,与太阳光保持垂直,测定稳态荧光(Fs)和光下最大荧光(Fm′)。通过计算,得到酸枣叶片的潜在光化学效率(Fv/Fm=(Fm-Fo)/Fm)、实际光化学效率(ΦPSⅡ=(Fm′-Fs)/Fm′)和非光化学淬灭系数(NPQ=(Fm-Fm′)/Fm′)等主要荧光参数[15]。
1.4 数据处理
采用SPSS 18.0,根据直角双曲线修正模型[16- 17]对酸枣叶片的Pn光响应过程进行非线性回归分析,得到Pn拟合值以及最大净光合速率(Pmax, μmol·m-2·s-1)、暗呼吸速率(Rd, μmol·m-2·s-1)、表观量子效率(AQY)、光补偿点(LCP, μmol·m-2·s-1)、光饱和点(LSP, μmol·m-2·s-1)、光抑制项(β)和光饱和项(γ)等光响应特征参数。采用Origin 7.5对不同水分条件下酸枣叶片光合参数的光响应过程进行三维绘图,对酸枣Pn光响应特征参数和叶绿素荧光参数与土壤水分的定量关系进行多项式回归分析并对拟合函数进行积分求解。采用SPSS 18.0对土壤水分点进行系统聚类分析,对不同处理间数据的差异显著性进行单因素方差分析和多重比较。
2 结果与分析
2.1 酸枣叶片光合参数与土壤水分的关系
2.1.1 不同水分条件下酸枣叶片气体交换参数的光响应
直角双曲线修正模型对不同水分条件下酸枣叶片的Pn光响应过程拟合效果较好,决定系数(R2)位于0.993—0.999之间,故采用Pn拟合值代替Pn实测值对Pn光响应过程进行研究。研究发现,PAR≤400 μmol·m-2·s-1时,酸枣叶片光合作用主要受低光强抑制,受土壤水分的影响不明显,强光下酸枣叶片的气体交换参数对土壤水分的响应呈现一定的规律性(图1)。酸枣叶片Pn、Tr和WUE均随PAR的增大先迅速上升后缓慢上升,随Wr的增大先增大后减小,均在Wr=73%时达到最大值,可见水分过高或过低均会抑制酸枣的光合作用、蒸腾作用及水分利用效率。酸枣WUE对土壤水分的变化幅度小于Pn和Tr,最大WUE在Wr为32%—56%和65%—79%之间差异均不显著(Pgt;0.05),始终保持较高水平,说明酸枣Tr对土壤水分的敏感程度大于Pn,水分逆境中,酸枣能够通过敏感的气孔开闭机制协调碳同化与水分消耗之间的关系,调节WUE。
弱光(PAR≤200 μmol·m-2·s-1)、干旱(Wr为11%—55%)及渍水(Wr为85%—95%)等条件对酸枣叶片Gs均具有一定的抑制作用,适宜的光照和水分条件有利于Gs的增大,Gs在Wr=74%,PAR=1600 μmol·m-2·s-1时达到最大值。酸枣叶片Ci随PAR的增大而减小,随Wr的增大呈“减小—增大—平稳—减小”的趋势,酸枣叶片Ls对光强和土壤水分的响应规律与Ci基本相反。根据气孔限制理论[18],当Wr为55%—80%时,酸枣Pn和Gs较高,Ci和Ls较稳定,表明酸枣光合作用的限制因素较少,土壤水分条件较适宜;当Wr为25%—55%或80%—95%时,Pn、Gs和Ci减小,Ls增大,表明酸枣光合作用在轻度干旱或渍水胁迫时主要受气孔限制;当Wr为11%—25%时,Pn、Gs和Ls减小,Ci增大,表明酸枣光合作用在严重干旱胁迫时主要受非气孔限制。
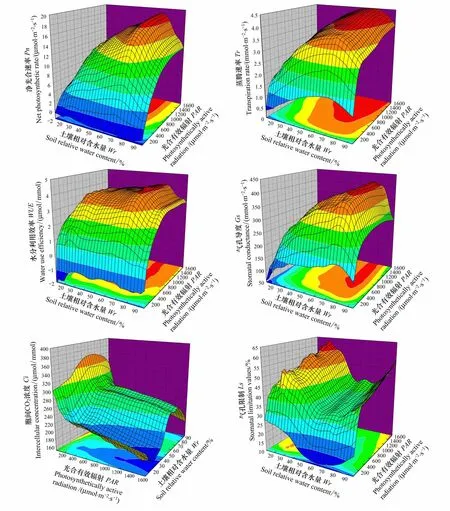
图1 不同水分条件下酸枣叶片的净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、气孔导度(Gs)、胞间CO2浓度(Ci)和气孔限制值(Ls)的光响应过程Fig. 1 Light response processes of net photosynthetic rate (Pn), transpiration rate (Tr), water use efficiency (WUE), stomatal conductance (Gs), intercellular CO2 concentration (Ci) and stomatal limitation values (Ls) of Ziziphus jujuba var. spinosus under different water content
2.1.2 酸枣叶片Pn光响应特征参数与土壤水分的关系
酸枣叶片Pn光响应特征参数与土壤水分定量关系的拟合中,通过对比发现四次方程的拟合效果较好,R2位于0.945—0.989之间。随土壤含水量的逐步增多,酸枣的Pmax、Rd、AQY和LSP呈先增大后减小的趋势,LCP、β和γ呈先减小后增大的趋势,不同参数对土壤水分的响应阈值存在一定差异(图2)。根据拟合函数求解极值,可知酸枣Pmax在Wr=73%时达到最大值18.2 μmol·m-2·s-1。根据拟合函数的积分结果求得试验土壤水分范围(Wr为11%—95%)内Pmax的平均值为14.6 μmol·m-2·s-1,对应的Wr为39%和88%,可知Pmax在Wr为39%—88%内维持较高水平,超出此水分范围后,Pmax随干旱或渍水胁迫的加剧迅速减小,酸枣光合潜力减弱。酸枣Rd、AQY和LSP分别在Wr为67%、72%和73%时达到最大值3.1 μmol·m-2·s-1、0.056和1704 μmol·m-2·s-1,三参数均随干旱(Wrlt;40%)或渍水(Wrgt;90%)胁迫的加剧而减小,表明酸枣的呼吸代谢活性以及对光能的利用能力均受到水分胁迫的限制。酸枣LCP、β和γ分别在Wr为58%、76%和72%时降至最小值36 μmol·m-2·s-1、0.56×10-4和1.08×10-3,当Wrlt;30%时,随土壤含水量的减少,LCP较缓慢地上升,β和γ大幅度地上升,表明随着干旱胁迫的加剧,酸枣对弱光的适应性减弱,同时在高光强下极易达到光饱和并出现光抑制[19]。酸枣光响应特征参数均在Wr为58%—76%内达到最适程度,表明适宜水分条件有利于增强酸枣叶片的光合活性。
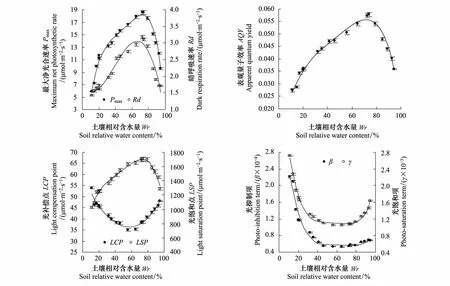
图2 不同水分条件下酸枣叶片Pn光响应特征参数的实测点与拟合曲线(平均值±标准误)Fig.2 Measured points and fitting curves of net photosynthetic rate-light response parameters of Ziziphus jujuba var. spinosus under different water conditions (mean±SE)
2.2 酸枣叶片叶绿素荧光参数与土壤水分的关系
对酸枣叶片的Fv/Fm、ΦPSⅡ、Fo和NPQ与土壤水分定量关系的拟合同样采用四次方程,R2位于0.689—0.952之间(图3)。酸枣ΦPSⅡ的变化幅度大于Fv/Fm,数值小于Fv/Fm,两指标均在Wr为80%时达到最大值,此时酸枣PSⅡ光化学效率最高。Fv/Fm在Wr为32%—95%内保持较高水平,差异不显著(Pgt;0.05),ΦPSⅡ在Wr为30%—80%或80%—95%内随干旱或渍水胁迫的加剧小幅度减小,表明ΦPSⅡ的降低并非由PSⅡ受损引起,而是酸枣光合作用减弱后对PSⅡ反馈抑制的结果。Fv/Fm和ΦPSⅡ均在Wrlt;30%时随干旱胁迫的加剧迅速减小,表明酸枣PSⅡ受到一定程度的损伤,光化学效率降低。Fo表征PSⅡ反应中心完全开放时的荧光强度,PSⅡ反应中心受到不易逆转的破坏会引起Fo的增加[20]。酸枣Fo在Wrlt;25%时随干旱胁迫的加剧迅速增大,表明酸枣PSⅡ在严重干旱胁迫时受到不易逆转的破坏。NPQ表征PSⅡ天线色素吸收的光能中以热的形式耗散掉的部分,热耗散是植物在光合作用达到光饱和、光合机构无法吸收过多光能情况下的一种保护机制[21]。酸枣NPQ在Wr为30%—80%或80%—95%的范围内随干旱或渍水胁迫的加剧而增大,表明酸枣能够通过热耗散消耗过剩光能实现自身光保护。NPQ在Wrlt;25%时随干旱胁迫的加剧而减小,表明酸枣热耗散能力减弱,PSⅡ受到不可逆的破坏。
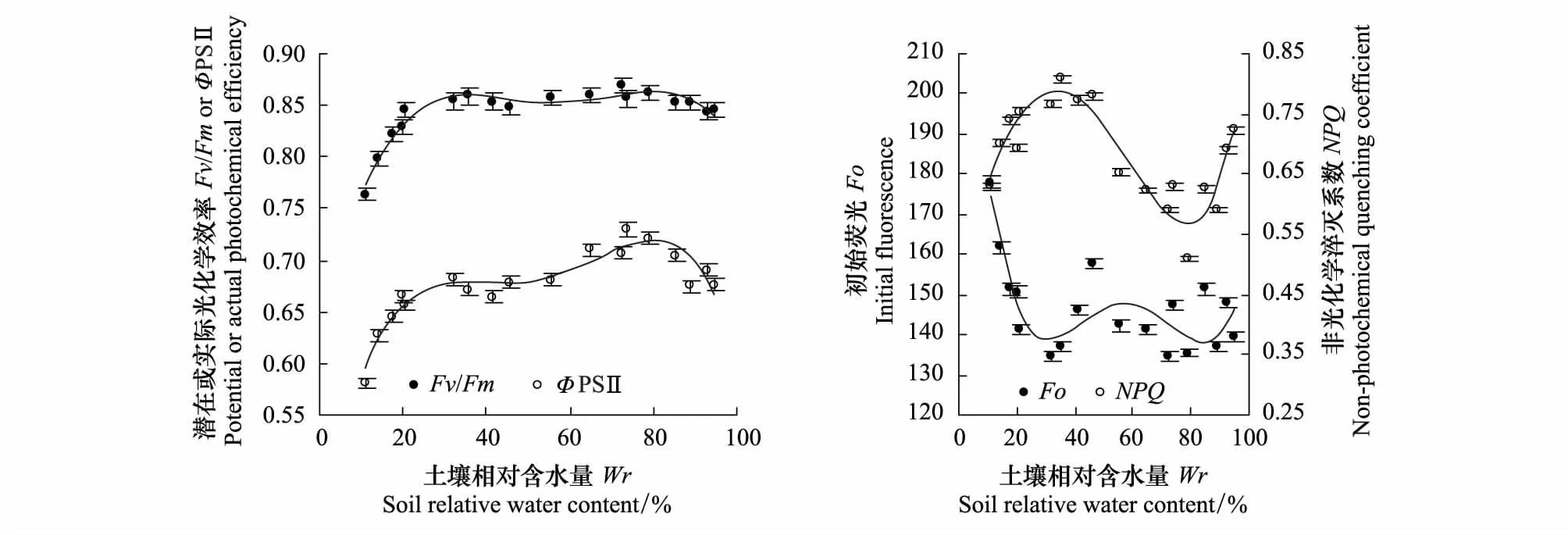
图3 不同水分条件下酸枣叶片主要叶绿素荧光参数的实测点与拟合曲线(平均值±标准误)Fig.3 Measured points and fitting curves of main chlorophyll fluorescence parameters of Ziziphus jujuba var. spinosus under different water conditions (mean ± SE)
2.3 酸枣叶片土壤水分生产力分级
分别根据酸枣在饱和光强下的Pn和WUE对所测土壤水分点进行聚类分析,设定聚类数为3,依据类平均值将酸枣的Pn和WUE划分为高、中和低3个等级,将Pn和WUE赋予“产”和“效”的光合生产力意义[22],结合酸枣主要光合生理参数对土壤水分的阈值效应,建立酸枣土壤水分生产力分级体系。依据酸枣Pn对土壤水分点聚类,分为11%—21%,32%—56%和85%—95%,以及65%—79%3类,相应为低产、中产和高产,差异显著(Plt;0.05),其中,低产和中产的Pn聚类均值分别为高产的0.45倍和0.77倍。依据酸枣WUE对土壤水分点聚类,分为11%—21%和85%—95%,32%—56%,以及65%—79%3类,相应为低效、中效和高效,差异显著(Plt;0.05),其中,低效和中效的WUE聚类均值分别为高效的0.75倍和0.92倍。酸枣叶片主要光合参数和叶绿素荧光参数均在Wr为58%—80%的范围内达到最适程度,因此,将Wr为58%作为划分中(高)产水、中(高)效水的临界点,将Wr为80%作为划分高(中)产水、高(低)效水的临界点。酸枣Pn下降原因由气孔限制转变为非气孔限制为主的土壤水分点出现在Wr为25%时,因此,将Wr为25%作为划分低(中)产水、低(中)效水的临界点。综合上述聚类水分点及光合生理参数土壤水分关键临界点,采用数学交集求解原理,确定出贝壳砂生境酸枣土壤水分生产力级别:Wr在11%—25%内为低产低效水,Wr在25%—58%内为中产中效水,Wr在58%—80%内为高产高效水,Wr在80%—95%内为中产低效水(表1)。
表1酸枣土壤水分生产力分级标准
Table1GradingcriterionofsoilmoistureproductivityforZiziphusjujubavar.spinosus.Differentlowercaselettersindicatesignificantdifferenceamongdifferentclustersofthesameparameterat0.05level
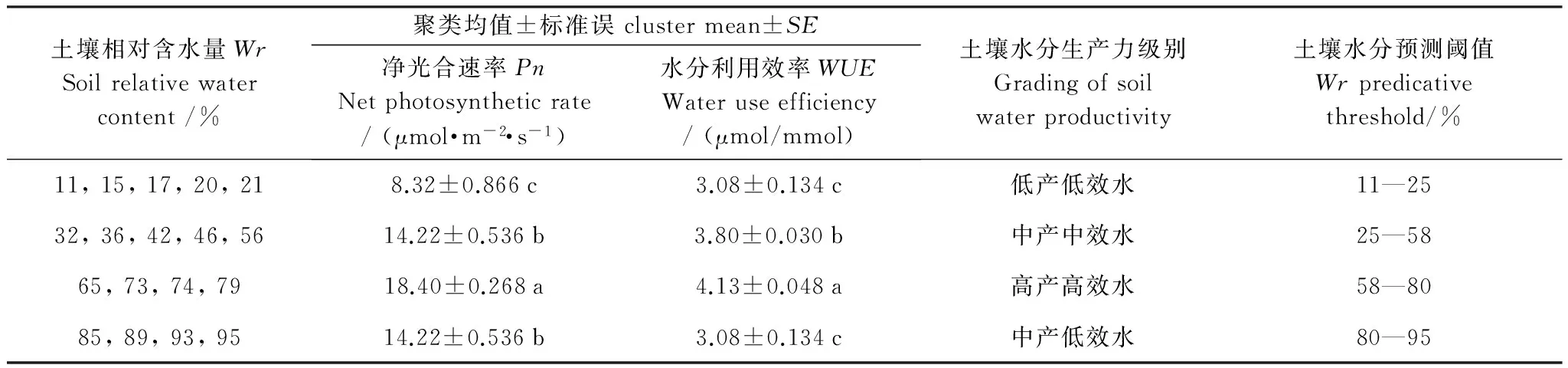
土壤相对含水量WrSoilrelativewatercontent/%聚类均值±标准误clustermean±SE净光合速率PnNetphotosyntheticrate/(μmol·m-2·s-1)水分利用效率WUEWateruseefficiency/(μmol/mmol)土壤水分生产力级别Gradingofsoilwaterproductivity土壤水分预测阈值Wrpredicativethreshold/%11,15,17,20,218.32±0.866c3.08±0.134c低产低效水11—2532,36,42,46,5614.22±0.536b3.80±0.030b中产中效水25—5865,73,74,7918.40±0.268a4.13±0.048a高产高效水58—8085,89,93,9514.22±0.536b3.08±0.134c中产低效水80—95
同一参数不同类群之间的不同小写字母表示差异显著(Plt;0.05)
3 讨论
3.1 酸枣叶片的光合作用效率
光合作用气孔限制转折点的判定对于确定酸枣光合生理需水的下限具有重要意义。气孔限制理论[18]认为,限制植物光合作用的因素分为气孔因素和非气孔因素,前者指叶片气孔的关闭程度及由此引起的气体交换难易程度,对光合作用的影响是可逆的;后者指光合机构的受损程度及由此引起的光合活性下降,对光合作用的影响是不可逆的。植物通过气孔限制降低光合作用,是其耐旱策略之一,而非气孔限制决定了光合作用的实际状态和潜力[23]。本研究发现,酸枣在轻度干旱或渍水胁迫时,光合作用主要受气孔限制,在严重干旱胁迫时,光合作用主要受非气孔限制,其转折点出现在Wr为25%时,金矮生苹果(Maluspumilacv. Goldspur)[3]和沙棘(HippophaerhamnoidesLinn.)[24]的转折点分别出现在Wr为48%和39%时,可见不同树种发生气孔限制转折的水分条件不尽相同,但酸枣的转折点出现在严重干旱胁迫时,表明其光合机构对干旱胁迫的适应能力较强,光合潜力较大。
Pmax是衡量植物叶片光合作用潜力的重要指标,在最适环境中,表示叶片的最大光合能力[25],酸枣Pmax的最大值为18.2 μmol·m-2·s-1,大于山杏和沙棘的最大值(13.3—15.5 μmol·m-2·s-1)[8, 24],表明酸枣的光合潜力较高。植物的呼吸作用是植物生长所必需的生理代谢活动[26],酸枣Rd的最大值为2.36 μmol·m-2·s-1,大于山杏[8]、杠柳(Periplocasepium)[17]和辽东楤木(Araliaelata)[27]的最大值(0.86—2.08 μmol·m-2·s-1),表明酸枣呼吸代谢活性高于一般灌木。AQY反映植物光合作用对光能的利用效率,酸枣AQY在Wr为72%时达到最大值(0.056),高于一般植物AQY范围(0.03—0.05)的上限[28],在干旱或渍水胁迫时降低,表明酸枣光合作用的光能利用效率较高,但对土壤水分响应敏感。在Wr为58%—73%内,酸枣LCP降至最小值,LSP达到最大值,利用弱光和强光的能力均较高,超出此水分范围后,随干旱或渍水胁迫的加剧,LCP增大,LSP减小,酸枣光照生态幅变窄。酸枣AQY、LCP和LSP对土壤水分的阈值效应表明酸枣在水分逆境中对光照的变化较敏感,具有通过降低光能利用率适应水分逆境的生理策略。
植物叶绿素吸收的光能主要通过光合电子传递、叶绿素荧光发射和热耗散3种途径,它们之间存在着此消彼长的关系[15],叶绿素荧光分析可以间接反映植物PSⅡ反应中心对光能的吸收、传递和转化的情况,同时叶绿素荧光技术因具有快速、灵敏、无损伤的特征而逐渐受到重视[29]。研究表明,非环境胁迫条件下Fv/Fm比较恒定,一般位于0.80—0.85之间[1],酸枣Fv/Fm在Wr为80%时达到最大值0.866,高于一般植物;酸枣ΦPSⅡ在Wr为80%时达到最大值0.718,大于沙棘的最大值(0.640)[24],以及大叶型丹参(Salviamiltiorrhizacv. Sativa)和小叶型丹参(Salviamiltiorrhizacv. Silcestris)的最大值(0.450—0.490)[30],表明酸枣PSⅡ对光能的吸收、传递和转化效率较高。当Wrlt;30%时,酸枣Fv/Fm和AQY作为表征光抑制的重要指标[31],均随干旱胁迫的加剧迅速减小,同时光抑制项β迅速增大,表明酸枣出现明显光抑制。当Wrlt;25%时,随干旱胁迫的加剧,酸枣Fo增大,NPQ减小,表明酸枣PSⅡ受到不可逆的破坏,与酸枣光合作用发生非气孔限制的水分阈值一致。
3.2 酸枣叶片光合作用的水分有效性
植物土壤水分生产力分级建立在植物水分生理学基础上,赋予Pn和WUE“产”和“效”的光合生产力意义,是植物光合生理水分有效性研究的重要手段。目前,不同植物在土壤水分生产力分级方法上存在一定差异。张光灿等[22]和陈建等[27]通过非线性回归分析、积分求解等方法确定植物主要光合参数(Pn、WUE、Tr和Ls等)与土壤水分的定量关系及各参数的土壤水分关键临界点(极值、平均值、补偿点和转折点等),根据各临界点将土壤水分划分为不同的生产力级别。夏江宝等[8]和张淑勇等[32]依据植物主要光合参数(Pn、WUE和Tr等)对系列土壤水分点进行聚类分析,结合主要参数对土壤水分的阈值效应,建立植物土壤水分生产力分级标准。本文依据酸枣叶片Pn和WUE分别对土壤水分点进行聚类分析,比依据植物主要光合参数共同对土壤水分点聚类的结果更加详细,并在重点分析Pn和WUE的同时兼顾酸枣叶片光合参数和叶绿素荧光参数的水分响应性,采用数学交集求解的方法确定酸枣土壤水分生产力级别,但各种分级方法适用于何种试验结果有待深入研究。
根据酸枣土壤水分生产力分级标准,当Wr为11%—25%时为低产低效水,酸枣光合生产力和水分利用效率均处于较低水平,同时光合作用受到非气孔限制,以PSⅡ反应中心为主的光合机构受到不可逆的破坏,严重影响酸枣苗木正常生长。当Wr为25%—58%时为中产中效水,Wr为80%—95%时为中产低效水,在这两个水分范围内酸枣光合生产力因受到气孔因素的可逆胁迫而处于中等水平,水分利用效率随干旱或渍水胁迫的加剧存在不同程度地减弱。当Wr为58%—80%时为高产高效水,酸枣主要光合生理参数均达到最适水平,光合生产力和水分利用效率处于较高水平,此范围是贝壳砂生境酸枣生长较为适宜的土壤水分条件。在干旱缺水地区,为有效提高水分利用效率,往往选择高产高效水或中产高效水作为植被生长较为适宜的土壤水分条件,而非高产中效水,如将Wr为48%—64%和41%—52%分别作为刺槐(Robiniapseudoacacia)和侧柏(Platycladusorientalis)适宜的土壤水分条件[22]。维持其他植物适宜生长的Wr为:金矮生苹果,60%—71%[3];山杏,47%—75%[8];辽东楤木,44%—85%[27];小叶扶芳藤(Euonymusfortuneivar.radicansSieb.),44%—72%[32]。其中,乔木刺槐和侧柏以及果树金矮生苹果的水分生态幅较窄,刺槐和侧柏适宜于较干旱的生境,金矮生苹果适宜于水分较充足的生境;灌木酸枣、山杏和辽东楤木以及藤本小叶扶芳藤的水分生态幅较宽,对不同水分生境具有较好的适应性。可见,植物适宜生长的土壤水分范围因植物种类和生境条件的不同而有较大差异。
综上所述,贝壳砂生境中酸枣不仅具有较高的光合能力,而且对水分逆境具有较强的生理适应性和可塑性。水分适宜(Wr为58%—80%)时,酸枣光合生理活性较高,利于酸枣苗木较好地生长;在轻度干旱(Wr为25%—58%)及渍水(Wr为80%—95%)胁迫时,酸枣光合作用主要受气孔限制,光合生理活性减弱;在严重干旱胁迫(Wr为11%—25%)时主要受非气孔限制,光合机构受到不易逆转的破坏,严重影响酸枣苗木的正常生长。
[1] Xu D Q. Photosynthetic Efficiency. Shanghai: Shanghai Science and Technology Press, 2002: 32- 33, 39- 52.
[2] Sofo A, Dichio B, Montanaro G, Xiloyannis C. Photosynthetic performance and light response of two olive cultivars under different water and light regimes. Photosynthetica, 2009, 47(4): 602- 608.
[3] Zhang S Y, Zhang G C, Gu S Y, Xia J B, Zhao J K. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation in semiarid loess hilly area. Photosynthetica, 2010, 48(4): 589- 595.
[4] Du Y D, Song L L, Liu Z X. An overview on theoretic research of high efficient water use in agriculture. Chinese Journal of Applied Ecology, 2003, 14(5): 808- 812.
[5] Wang Y, Chen J W, Di X Y. Characterization of the responses of photosynthetic and chlorophyll fluorescence parameters to water stress in seedlings of six provenances of Chinese Pine (PinustabulaeformisCarr.). Acta Ecologica Sinica, 2011, 31(23): 7031- 7038.
[6] Wang Z X, Chen L, Ai J, Qin H Y, Liu Y X, Xu P L, Jiao Z Q, Zhao Y, Zhang Q T. Photosynthesis and activity of photosystem II in response to drought stress in Amur Grape (VitisamurensisRupr.). Photosynthetica, 2012, 50(2): 189- 196.
[7] Gao L, Yang J, Liu R X. Effects of soil moisture levels on photosynthesis, transpiration, and moisture use efficiency of female and male plants ofHippophaerhamnoidesssp.sinensis. Acta Ecologica Sinica, 2009, 29(11): 6025- 6034.
[8] Xia J B, Zhang G C, Sun J K, Liu X. Threshold effects of photosynthetic and physiological parameters inPrunussibiricato soil moisture and light intensity. Chinese Journal of Plant Ecology, 2011, 35(3): 322- 329.
[9] Li H J, Li P, Yu G D. Advance of research and prospect of development ofZizyphusjujubavar.spinosa(Bunge) Hu ex H F Chow. Chinese Wild Plant Resources, 1999, 18(3): 15- 19.
[10] Cui X D. Research on the fine individual selection and fast propagation technology of wild jujube resources. Journal of Anhui Agricultural Sciences, 2011, 39(8): 4464- 4466.
[11] He S X, Liang Z S, Yu L Z, Zhou Z Y. Growth and physiological characteristics of wild sour jujube seedlings from two provenances under soil water stress. Acta Botanica Boreali-Occidentalia Sinica, 2009, 29(7): 1387- 1393.
[12] Zhou Z Y, Liang Z S, Li S, Lai W F. Effect of water stress and re-watering on relative water content, protective enzyme and photosynthetic characteristics of wild jujube. Chinese Journal Eco-Agriculture, 2011, 19(1): 93- 97.
[13] Prior S A, Runion G B, Rogers H H, Arriaga F J. Elevated atmospheric carbon dioxide effects on soybean and sorghum gas exchange in conventional and notillage systems. Journal of Environmental Quality, 2010, 39(2): 596- 608.
[14] Berry J A, Downton W J S. Environmental regulation of photosynthesis // Govindjee, ed. Photosynthesis, VolumeⅡ. New York: Academic Press, 1982: 263- 342.
[16] Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis inOryzasativa. Photosynthetica, 2007, 45(4): 637- 640.
[17] Wang R R, Xia J B, Yang J H, Zhao Y Y, Liu J T, Sun J K. Comparison of light response models of photosynthesis in leaves ofPeriplocasepiumunder drought stress in sand habitat formed from seashells. Chinese Journal of Plant Ecology, 2013, 37(2): 111- 121.
[18] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33(1): 317- 345.
[19] Ye Z P, Kang H J. Study on biological significance of coefficients in modified model of photosynthesis-irradiance. Journal of Yangzhou University: Agricultural and Life Science Edition, 2012, 33(2): 51- 57.
[20] Krause G H. Photoinhibition of photosynthesis: an evaluation of damaging and protective mechanisms. Physiologia Plantarum, 1988, 74(3): 566- 574.
[21] Gilmore A M, Yamamoto H Y. Zeaxanthin formation and energy-dependent fluorescence quenching in pea chloroplasts under artificially mediated linear and cyclic electron transport. Plant Physiology, 1991, 96(2): 635- 643.
[22] Zhang G C, Xia J B, Shao H B, Zhang S Y. Grading woodland soil water productivity and soil bioavailability in the semi-arid Loess Plateau of China. Clean-Soil, Air, Water, 2012, 40(2): 148- 153.
[23] Li Q, Wang M, Wang W W, Zhang R C, Yue M. Response of photosynthetic characteristics ofPsathyrostachyshuashanicaKeng to drought stress. Acta Ecologica Sinica, 2012, 32(13): 4278- 4284.
[24] Pei B, Zhang G C, Zhang S Y, Wu Q, Xu Z Q, Xu P. Effects of soil drought stress on photosynthetic characteristics and antioxidant enzyme activities inHippophaerhamnoidesLinn. seedings. Acta Ecologica Sinica, 2013, 33(5): 1386- 1396.
[25] Tartachnyk I I, Blanke M M. Effect of delayed fruit harvest on photosynthesis, transpiration and nutrient remobilization of apple leaves. New Phytologist, 2004, 164(3): 441- 450.
[26] Tjoelker M G, Oleksyn J, Lee T D, Reich P B. Direct inhibition of leaf dark respiration by elevated CO2is minor in 12 grassland species. New Phytologist, 2001, 150(2): 419- 424.
[27] Chen J, Zhang G C, Zhang S Y, Wang M J. Response processes ofAraliaelataphotosynthesis and transpiration to light and soil moisture. Chinese Journal of Applied Ecology, 2008, 19(6): 1185- 1190.
[28] Li H S. Modern Plant Physiology. Beijing: Higher Education Press, 2002.
[29] Zhang R H, Zheng Y J, Ma G S, Zhang X H, Lu H D, Shi J T, Xue J Q. Effects of drought stress on photosynthetic traits and protective enzyme activity in maize seeding. Acta Ecologica Sinica, 2011, 31(5): 1303- 1311.
[30] Luo M H, Hu J Y, Wu Q G, Yang J T, Su Z X. Effects of drought stress on leaf gas exchange and chlorophyll fluorescence ofSalviamiltiorrhiza. Chinese Journal of Applied Ecology, 2010, 21(3): 619- 623.
[31] Demmig-Adams B, Adams W W III. Photoprotection and other responses of plants to high light stress. Annual Review of Plant Physiology and Plant Molecular Biology, 1992, 43(1): 599- 626.
[32] Zhang S Y, Zhou Z F, Xia J B, Zhang G C. The responses ofEuonymusfortuneivar.radicansSieb. leaf photosynthesis to light in different soil moisture. Acta Botanica Boreali-Occidentalia Sinica, 2007, 27(12): 2514- 2521.
参考文献:
[1] 许大全. 光合作用效率. 上海: 上海科学技术出版社, 2002: 32- 33, 39- 52.
[4] 杜尧东, 宋丽莉, 留作新. 农业高效用水理论研究综述. 应用生态学报, 2003, 14(5): 808- 812.
[5] 王琰, 陈建文, 狄晓燕. 不同油松种源光合和荧光参数对水分胁迫的响应特征. 生态学报, 2011, 31(23): 7031- 7038.
[7] 高丽, 杨劼, 刘瑞香. 不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征. 生态学报, 2009, 29(11): 6025- 6034.
[8] 夏江宝, 张光灿, 孙景宽, 刘霞. 山杏叶片光合生理参数对土壤水分和光照强度的阈值效应. 植物生态学报, 2011, 35(3): 322- 329.
[9] 李会军, 李萍, 余国奠. 酸枣的研究进展及开发前景. 中国野生植物资源, 1999, 18(3): 15- 19.
[10] 崔向东. 野生酸枣资源选优与快速繁殖技术研究. 安徽农业科学, 2011, 39(8): 4464- 4466.
[11] 贺少轩, 梁宗锁, 蔚丽珍, 周自云. 土壤干旱对2个种源野生酸枣幼苗生长和生理特性的影响. 西北植物学报, 2009, 29(7): 1387- 1393.
[12] 周自云, 梁宗锁, 李硕, 来威锋. 干旱- 复水对酸枣相对含水量、保护酶及光合特征的影响. 中国生态农业学报, 2011, 19(1): 93- 97.
[17] 王荣荣, 夏江宝, 杨吉华, 赵艳云, 刘京涛, 孙景宽. 贝壳砂生境干旱胁迫下杠柳叶片光合光响应模型比较. 植物生态学报, 2013, 37(2): 111- 121.
[19] 叶子飘, 康华靖. 植物光响应修正模型中系数的生物学意义研究. 扬州大学学报: 农业与生命科学版, 2012, 33(2): 51- 57.
[23] 李倩, 王明, 王雯雯, 张瑞昌, 岳明. 华山新麦草光合特性对干旱胁迫的响应. 生态学报, 2012, 32(13): 4278- 4284.
[24] 裴斌, 张光灿, 张淑勇, 吴琴, 徐志强, 徐萍. 土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响. 生态学报, 2013, 33(5): 1386- 1396.
[27] 陈建, 张光灿, 张淑勇, 王梦军. 辽东楤木光合和蒸腾作用对光照和土壤水分的响应过程. 应用生态学报, 2008, 19(6): 1185- 1190.
[28] 李合生. 现代植物生理学. 北京: 高等教育出版社, 2002.
[29] 张仁和, 郑友军, 马国胜, 张兴华, 路海东, 史俊通, 薛吉全. 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响. 生态学报, 2011, 31(5): 1303- 1311.
[30] 罗明华, 胡进耀, 吴庆贵, 杨敬天, 苏智先. 干旱胁迫对丹参叶片气体交换和叶绿素荧光参数的影响. 应用生态学报, 2010, 21(3): 619- 623.
[32] 张淑勇, 周泽福, 夏江宝, 张光灿. 不同土壤水分条件下小叶扶芳藤叶片光合作用对光的响应. 西北植物学报, 2007, 27(12): 2514- 2521.
ResponsecharacteristicsofphotosyntheticandphysiologicalparametersinZiziphusjujubavar.spinosusseedlingleavestosoilwaterinsandhabitatformedfromseashells
WANG Rongrong1,2, XIA Jiangbao2,*, YANG Jihua1, LIU Jingtao2, ZHAO Yanyun2, SUN Jingkuan2
1CollegeofForestry,ShandongAgriculturalUniversity,Taian,Shandong271018,China2BinzhouUniversity,ShandongProvincialKeyLaboratoryofEco-EnvironmentalScienceforYellowRiverDelta,Binzhou,Shandong256603,China
Three-year-old seedlings ofZiziphusjujubavar.spinosus, a dominant shrub in shell ridge island of China′s Yellow River Delta, were used as experimental materials. A series of soil water content in sand formed from seashells were simulated by artificial feeding firstly and then plant transpiring. Light responses of photosynthesis and chlorophyll fluorescence parameters in leaves ofZ.jujubavar.spinosusunder series water content were measured with CIRAS-2 portable photosynthesis system and FMS-2 portable fluorescence system, respectively. The threshold effects of the main photosynthetic and physiological parameters in leaves ofZ.jujubavar.spinosusto soil water and their response relationships were elucidated, and the soil water availability onZ.jujubavar.spinosusleaves′ photosynthesis was studied. The results showed that: (1) The net photosynthetic rate (Pn), transpiration rate (Tr) and water use efficiency (WUE) in leaves ofZ.jujubavar.spinosusfirst increased and then decreased with the soil water increasing, whileTrwas more sensitive to soil water variation thanPn, thus causingWUEat a higher level. (2) The main reason ofPndeclining changed from stomatal limitation to non-stomatal limitation in leaves ofZ.jujubavar.spinosuswhen soil relative water content (Wr) reduced to 25%,Pnwas dominated by stomatal limitation whenWrgt; 25%, and non-stomatal limitation whenWrlt; 25%, meanwhile photosynthetic apparatus was damaged irreversibly. (3) The maximum net photosynthetic rate, dark respiration rate, apparent quantum yield (AQY) and light saturation point in leaves ofZ.jujubavar.spinosusfirst increased and then decreased, while light compensation point, photo-inhibition term (β) and photo-saturation term first decreased and then increased with the soil water increasing. ThePnlight response parameters above all reached the optimum levels whenWrvaried from 58% to 76%, meanwhile the photosynthetic activity was higher. (4) The potential photochemical efficiency (Fv/Fm) and actual photochemical efficiency ofZ.jujubavar.spinosusfirst increased and then decreased with the soil water increasing, the photochemical transformation efficiency of PSⅡ reaction center reached maximum whenWrwas 80%. Non-photochemical quenching coefficient (NPQ) increased gradually with the aggravation of drought or waterlogging stress whenWrvaried from 80% to 30% or from 80% to 95%,Z.jujubavar.spinosusPSⅡ had strong ability of heat dissipation and could prevent photosynthetic apparatus from damage by dissipating excessive light energy.AQYandFv/Fmdecreased rapidly andβincreased rapidly whenWrlt; 30%, photo-inhibition occurred inZ.jujubavar.spinosusleaves significantly.NPQdecreased rapidly and initial fluorescence increased rapidly whenWrlt; 25%,Z.jujubavar.spinosusPSⅡ was damaged irreversibly. (5) 11%—25% ofWrwas low productivity and lowWUEforZ.jujubavar.spinosus, 25%—58% was middle productivity and middleWUE, 58%—80% was high productivity and highWUE, 80%—95% was middle productivity and lowWUE. In summary, the photosynthesis ofZ.jujubavar.spinosushad great physiological adaptability and plasticity to soil water stress in sand formed from seashells. The photosynthetic and physiological activity ofZ.jujubavar.spinosusmaintained at a relatively high level whenWrwas between 58% and 80%, meanwhileZ.jujubavar.spinosusseedling could grow well, and the soil water maximum deficit level allowed in sand habitat forZ.jujubavar.spinosusoccurred whenWrwas 25%.
shell ridge island; gas exchange parameter; chlorophyll fluorescence parameter; photosynthetic productivity; water use efficiency; threshold effect
国家自然科学基金资助项目(31100468, 41201023);山东省高校科研发展计划资助项目(J13LC03, J13LE57)
2013- 05- 16;
2013- 07- 15
*通讯作者Corresponding author.E-mail: xiajb@163.com
10.5846/stxb201305161088
王荣荣,夏江宝,杨吉华,刘京涛,赵艳云,孙景宽.贝壳砂生境酸枣叶片光合生理参数的水分响应特征.生态学报,2013,33(19):6088- 6096.
Wang R R, Xia J B, Yang J H, Liu J T, Zhao Y Y, Sun J K.Response characteristics of photosynthetic and physiological parameters inZiziphusjujubavar.spinosusseedling leaves to soil water in sand habitat formed from seashells.Acta Ecologica Sinica,2013,33(19):6088- 6096.
猜你喜欢
今日农业(2022年13期)2022-09-15
中成药(2021年5期)2021-07-21
趣味(语文)(2021年12期)2021-04-19
河北果树(2020年1期)2020-02-09
小哥白尼(野生动物)(2019年6期)2019-10-10
读友·少年文学(清雅版)(2018年4期)2018-01-14
高原山地气象研究(2016年2期)2016-11-10
学苑创造·A版(2016年5期)2016-06-21
塔里木大学学报(2014年3期)2014-03-11
新疆农垦科技(2014年7期)2014-02-28
