
2011年秋季中街山列岛邻近海域浮游植物群落的生态研究
2013-10-21 08:31陈应华
浙江海洋大学学报(自然科学版) 2013年3期
阳 丹,陈应华
(国家海洋设施养殖工程技术研究中心,浙江舟山 316004)
在海洋生态系统中,海洋浮游植物作为初级生产者,在物质循环和能量循环中起着至关重要的作用。近年来,随着全球温室效应现象的日益加剧,浮游植物对海洋碳循环和全球气候变化中间的关联作用越来越被受到重视。此外,浮游动物,作为鱼虾贝的直接或间接饵料,其群落组成及丰度为海洋渔业资源环境评估提供重要参考数据。
中街山列岛邻近海域地处我国东海,是著名的舟山渔场重要组成部分。作为国家级海洋特别保护区,其优越的环境曾是大黄鱼,小黄鱼,带鱼及曼氏无针乌贼等海洋经济生物的重要繁衍栖息场所。但近年来,随着我国临海工业,海上交通运输及水产养殖业的飞速发展,由于缺乏完善的管理及专业科学技术,中街山列岛邻近海域的生态环境日益恶化。海水富营化程度逐年加剧,赤潮频发,给该片海域乃至整个舟山渔场生态环境带来了严重影响。海洋浮游植物群落作为海洋生态环境检测的一项重要指标,能间接反映海洋生态系统的受污染程度。为了摸清中街山列岛邻近海域环境现状,2011年秋季在该海域进行了海洋浮游植物群落生长情况的大面站调查。根据调查结果,对该海域的浮游植物群落生长状况及与环境间的关系作了深入的分析评价,以期为中街山列岛邻近海域生态环境的修复提供基本依据。
1 材料与方法
1.1 调查海域和站位
2011年11月16 日至2011年11月19日于中街山列岛邻近海域,共12个站位(图1)按照《海洋调查规范》进行了多学科综合调查。
1.2 样品采集与分析
现场使用YSI自动水质检测仪测定水温(Temperature)、透明度(Transparency)及水体 pH值。水化指标总磷(Total phosphate,TP)、总氮(Total nitrogen,TN)、正磷酸(Orthophosphate,PO4-P)、硝氮(Nitrate nitrogen,NO3-N)、亚硝氮 (Nitrite nitrogen,NO2-N),氨氮(Ammonia nitrogen,NH4-N),按照国家水质标准方法GB3838-2002进行实验。使用浅水型浮游生物网(网口直径37 cm,网长140 cm,筛绢孔径为0.077 mm)由底至表垂直拖网采集,所获标本经5%甲醛现场固定,实验室进行分类鉴定及统计。
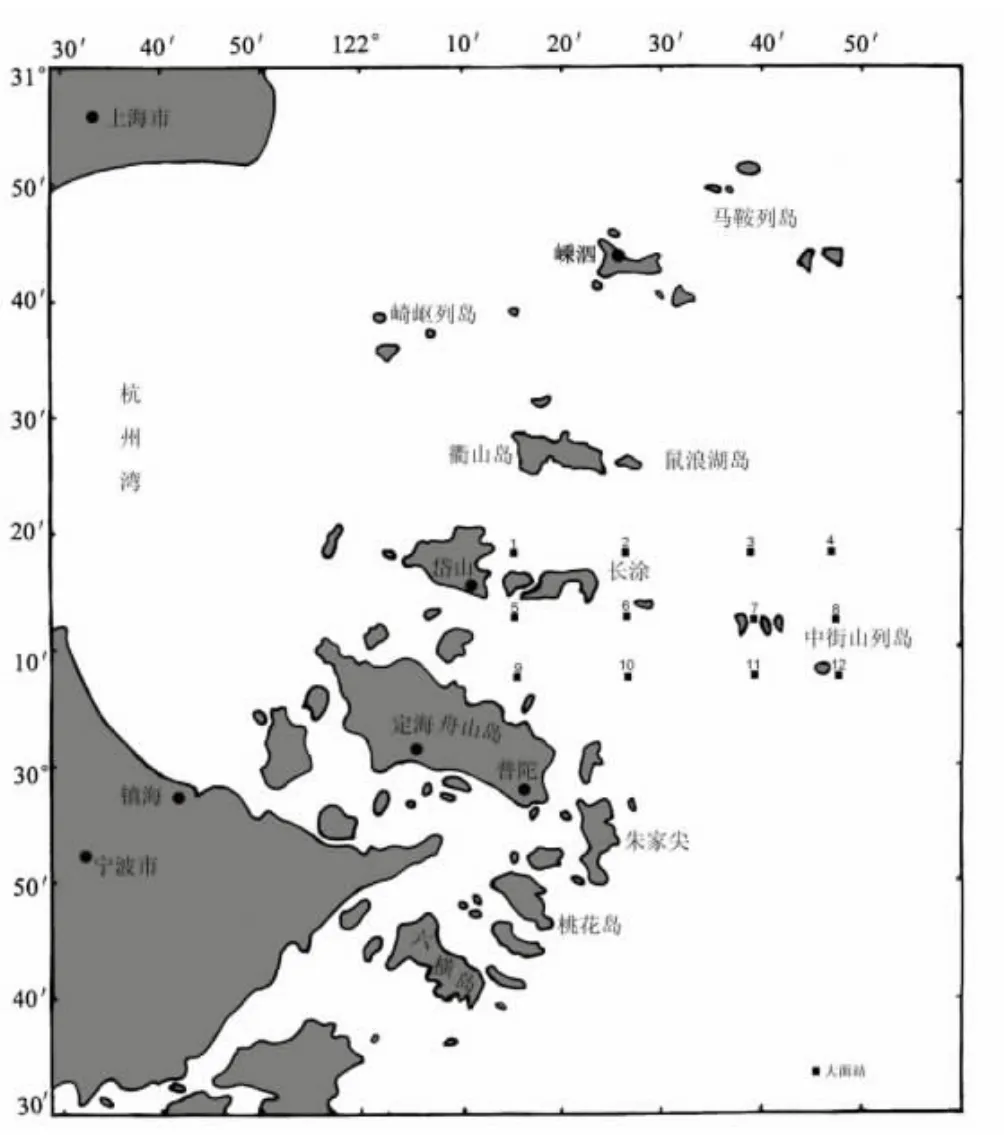
图1 调查站位Fig.1 The sampling locations in Zhongjieshan archipelago waters and its neighborhood
1.2 数据处理方法
浮游植物群落结构分别采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和优势度指数(Y)进行评价分析,公式如下:
Shannon-Wiener多样性指数计算公式:
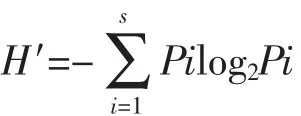
在公式中,S代表此样品的物种数,Pi代表采集样品中第i种的细胞丰度与该样品总细度丰度的比值。
Pielou均匀度指数计算公式:
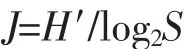
公式中,S代表样品物种数。
优势度指数(Y)计算公式:
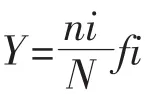
公式中,ni代表第i个种在采集的全部样品中的总细胞丰度,N代表所有才集样品的物种的总细胞丰度,fi代表该物种在所有站位出现的频率。
2 结果与分析
2.1 调查海区环境特征
在图2、3中,分别显示了调查海区表层水体温度,盐度数值的平面分布情况。其中,该海域表层水体温度变化范围介于18.7~25.8℃之间,平均值为21.9℃。由于同时受太阳辐射以及长江冲淡水、沿岸径流的影响,从平面分布图中不难看出,海域表层水体温度在近岸海域基本呈现低值分布,而随着离岸方向,温度大致呈现逐渐上升的趋势,如在离岸较远的St4站,出现本次调查海域温度的最高值25.8。调查海域表层水体盐度的变化范围介于27.8~35.4之间,平均值为31.6。在调查站的St8站,出现了本次调查的盐度最高值。同样受长江冲淡水以及沿岸径流的影响,调查表层水体盐度在近岸海域数值依然较低,随着离岸方向,盐度值基本呈现出越来越高的态势。
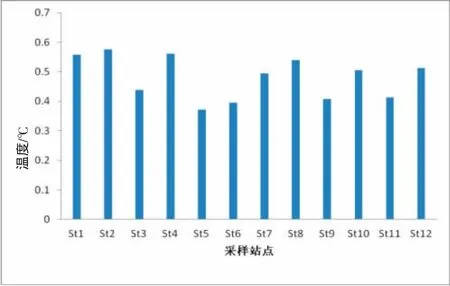
图2 海区表层温度Fig.2 The surface layer temperature in surveyed oceanic area
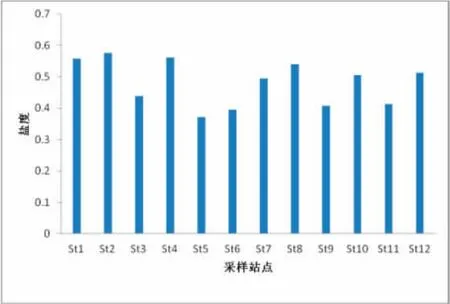
图3 海区表层盐度Fig.3 The surface layer salinity in surveyed oceanic area
2.2 调查海区浮游植物物种组成
分析所获浮游植物样品,共鉴定浮游植物4门49属127种(不包括未定名物种);其中,硅藻38属98种(不包括未定名物种),占所有样品鉴定物种总数的77.2%;甲藻7属24种,占所有物种种数的18.9%;金藻3属4种,蓝藻1属1种。在秋季调查水样中,硅藻和甲藻是主要的浮游植物类群,而硅藻类占据了物种种数的绝大部分。可见中街山列岛邻近海域秋季,浮游植物类群仍以硅藻为主,其次为甲藻。物种的生态类型主要可分为广温广盐近岸类群,代表种为角毛藻Chaetoceros、角藻Ceratium、布氏双尾藻Ditylum bright wellii、海洋原甲藻Prorocentrum micans、夜光藻Noctiluca scintillans等;以及暖温近岸低盐类群,其代表藻种有中肋骨条藻Skeletonema costatum、尖刺拟菱形藻Pseudo-nitzschia pungens等。本次对该海域的调查中,浮游植物中高温高盐类群,高温次高盐类群以及偏低温高盐类群所有物种总数中的比例不大。
调查海域优势种见表1。中街山列岛邻近海域优势藻种同以往对该海域调查结果相比,微型的中肋骨条藻,小型的佛氏海线藻Thalassionema frauenfeldii、琼氏圆筛藻Coscinodiscus jonesianus、角毛藻等仍是该片海域的优势藻种,其藻细胞丰度的比例之和高达79.8%,出现频率指数也均较高,以上优势藻种都属于该片海域常见的赤潮原因种。但较以往不同的是,本次调查优势种中出现了泰晤士扭鞘藻Helicotheca tamesis、颤藻Oscillatoria,其藻种丰度比例及出现频率都较高。由优势种分析结果可见,该海域的生态环境或许在一定程度上发生了改变,并进而影响到了浮游植物群落优势种的构成。
2.3 调查海区浮游植物的细胞丰度特征
调查海域浮游植物的细胞丰度介于 0.373×103~199.325×103cell/L,平均丰度值为137.458×103cell/L。浮游植物群落中,硅藻在丰度值上占有绝对的优势,丰度比例为总细胞丰度值的81.2%,其丰度值介于0.482×103~142.475×103cell/L,平均丰度值为83.683×103cell/L。甲藻所占丰度比例其次,数值为 16.9%。丰度值介于 0.660×103~103.115×103cell/L,平均丰度值为47.265×103cell/L。与以往对该海域的调查结果相比,此次调查的丰度值略偏低。这可能与本次海洋调查的站点设置覆盖范围较小,加之使用拖网采集样品的方法,丰度值测算结果易造成较大的误差有关;另外,与浮游植物细胞丰度较大的时空变异特性也有着密切关联(图4)。
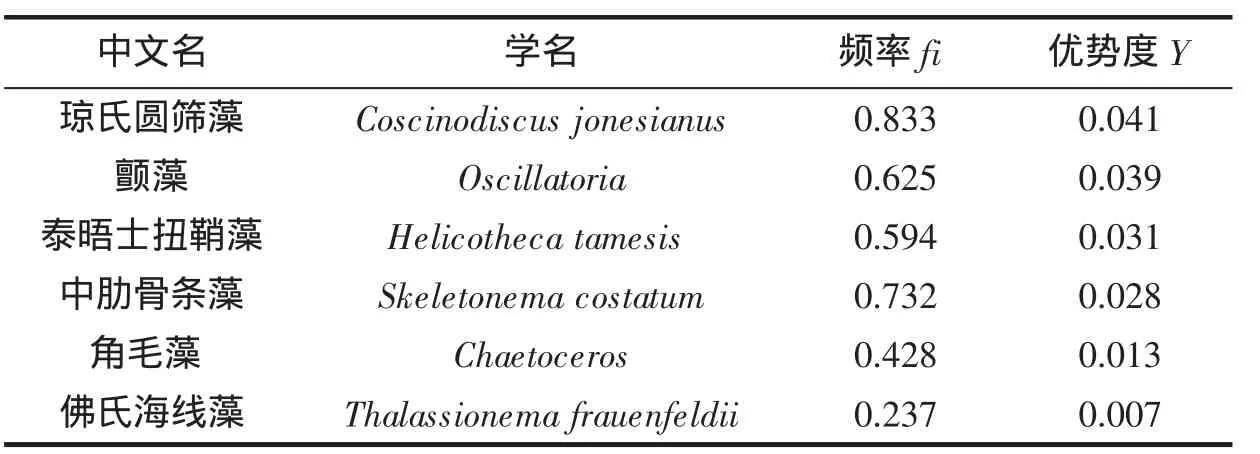
表1 调查海区浮游植物优势种Tab.1 Dominant species of phytoplankton in surveyed oceanic area
2.3.1 表层浮游植物细胞丰度的特征
如图5所示,在调查海区表层水体中,浮游植物细胞丰度值介于 0.862×103~201.473×103cell/L 间,其平均丰度值为148.643×103cell/L。该水层浮游植物群落仍然主要由硅藻和甲藻组成,二者的细胞丰度比例占据了所有细胞丰度的99.7%。硅藻细胞丰度同样在调查海区表面水体中占具大优势,其丰度比例达到了83.7%,丰度值介于 0.862×103~146.573×103cell/L,平均值为89.853×103cell/L。甲藻细胞丰度在表层水体中仅占 16.0%,丰度值介于 0.660×103~107.384×103cell/L间,平均值为49.315×103cell/L。细胞丰度在该调查海域存在2个高值区,其中最大高值区位于岱山岛邻近的St1站,此处站点海域靠近港口码头,且有大量的工业废水,居民生活污水排入,海水富营养化程度较高;次高值区位于中街山列岛养殖海区的St7站,该站位所在海域因常年养殖海洋经济生物,富营化程度相对较高。与此同时,调查海域表层细胞丰度呈现在近岸海域数值高,离岸海域数值低的特征。
2.3.2 表层优势种细胞丰度的特征
如图6所示,中肋骨条藻在调查海域的大部分站位均有分布,其表层丰度值介于4.455×103~124.245×103cell/L之间,平均丰度值为 58.372×103cell/L,其高值区主要分布在岱山岛、长凃岛附近的近岸海域。琼氏圆筛藻在分布站位的丰度值均较高,介于4.125×103~146.573×103cell/L之间,平均丰度值为93.703×103cell/L。峰度值位于中街山岛养殖海区的St7站位,且在该养殖区域的St5、St6站,琼氏圆筛藻丰度值仍然很高。这与此片海域常年进行水产养殖所导致的水体富营养化有直接关系。同以往历史资料相比,此次对该海域的调查,优势种中新增了隶属于蓝藻门的颤藻,其表层丰度值介于0.932×103~201.473×103cell/L之间,平均丰度值为151.040×103cell/L。其在St1及St7站位出现的峰度值已远远超过其它优势藻种。颤藻在此次调查优势种中的出现,不仅与中街山列岛邻近海域地处长江口区域有关,而且也说明近年来,该海域随着水体富营养化的加剧,浮游植物群落生长也在逐步发生变化。
2.3.3 细胞丰度的垂直分布
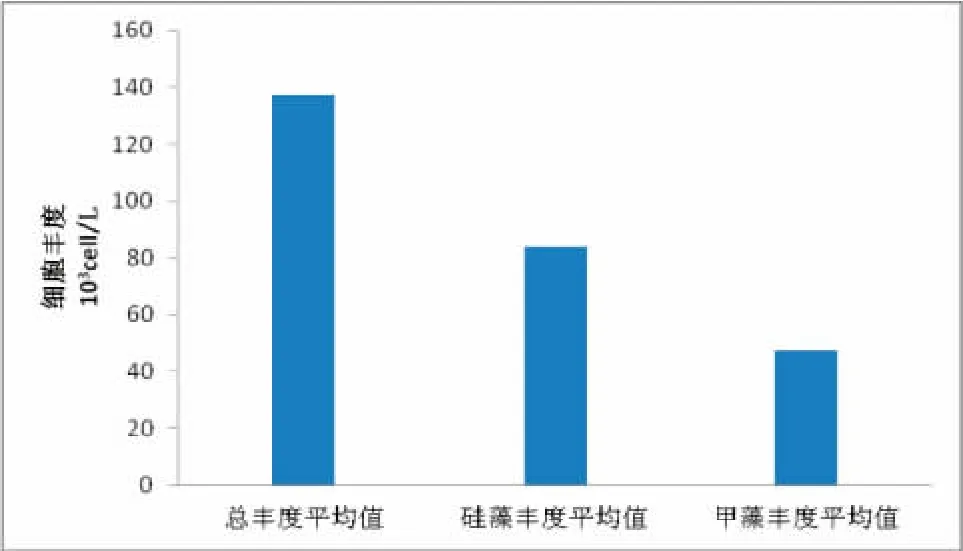
图4 调查海区浮游植物的细胞丰度特征Fig.4 The cell abundance characteristics of phytoplankton in surveyed oceanic area
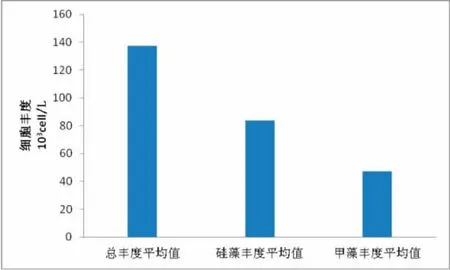
图5 调查海区浮游植物表层的丰度特征Fig.5 The surface layer abundance chrarcteristics of phytoplankton in surveyed oceanic area
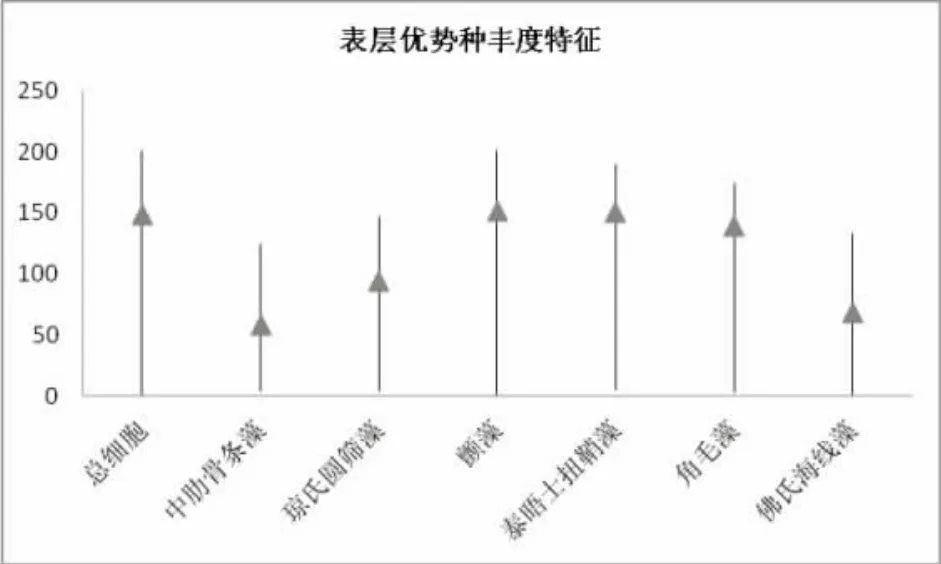
图6 表层优势种细胞丰度的特征Fig.6 The cell abundance characteristics of dominant species of phytoplankton in surface layer
如图7所示,显示了调查海区浮游植物群落在垂直方向上的分布特点。不难看出,该片海区浮游植物群落整体上有随海水深度的增加,群落细胞丰度逐渐减小的趋势。在调查海区表层,浮游植物细胞丰度达到了相比其它水层的最大值。但对于各藻种单独而言,并不完全是随水深深度增加丰度值渐减。此次调查,在优势种中,中肋骨条藻的细胞丰度在水下30 m的水层才达到最大值,而在水体表层与次表层中,其细胞丰度相差不大。琼氏圆筛藻在表层水体中细胞丰度达到最大值,但在水下30 m的细胞丰度却大于次表层。优势种中新出现的蓝藻门中的颤藻,在表层出现其细胞丰度的最大值,且随着水体深度的加深,其细胞丰度大致有逐减变小的趋势。
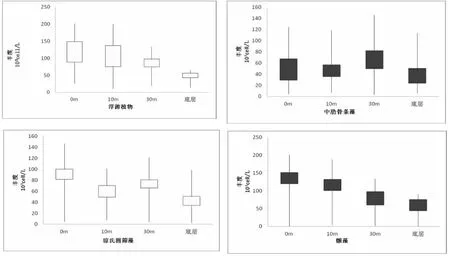
图7 细胞丰度的垂直分布Fig.7 The vertical distribution of the cell abundance
2.4 群落多样性
群落多样性指数能客观地反应出群落的分布特征以及群落的稳定性,通常情况下,对于调查区群落多样性指数的测定,一般选用Shannon-Wiener多样性指数和Pielou均匀度指数来进行综合评估。Shannon-Wiener多样性指数一般用于反映调查区群落结构的复杂性程度,而Pielou指数则多用于来客观反映群落的均匀度。本次调查海域表层浮游植物群落的以上两种参数值平面分布特征见图7。
调查海域表层浮游植物群Shannon-Wiener多样性指数(图8)介于2.594~4.028之间,平均值为3.393。其中,高值区分布在长涂岛附近的St2站及离岸海域的St4站,由于上述两站位均分布在长江口邻近海域,受长江冲淡水的影响,所在海区营养盐丰富,且水体透明度较好,易于浮游植物群落正常光合作用的同时吸收大量营养盐。故浮游指物多样性指数出现最高值。低值区出现在岱山岛附近海域的St5站,由于该调查站位处于近岸海域,水体浑浊,透明度很低,除近岸浮游植物群落外,并不利于浮游植物各物种的大量生长,故相关指数出现最低值。从分析数据结果来看,该海域的多样性指数整体均偏高,这可能是由于调查区所在海域隶属于舟山渔场海域,营养盐水平较高,加之该区域浮游植物物种丰富,使得浮游植物群落大量生长,多个优势种并存,并最终导致了调查海区整体上多样性指数偏高,群落结构复杂。
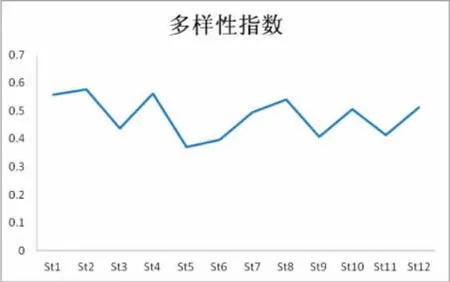
图8 Shannon-Wiener多样性指数Fig.8 Shannon-Wiener′s diversity indeX
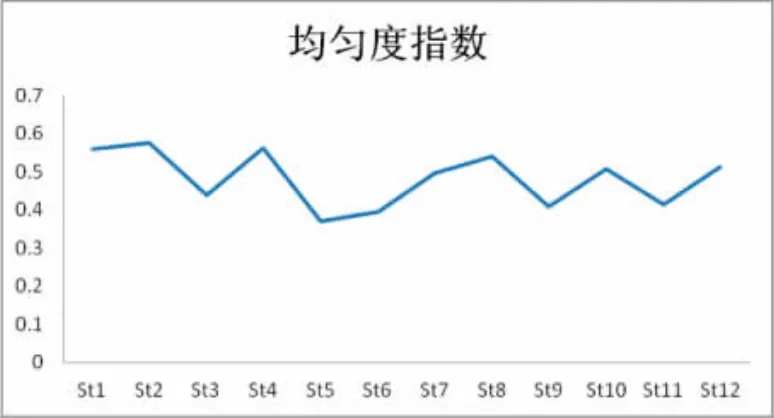
图9 Pielou均匀度指数Fig.9 Pielou′s evenness index
调查海域表层浮游植物群落Pielou均匀度指数(图9)介于0.371~0.576之间,平均值为0.485。其中高值区出现在长涂岛附近海域的St2站位,而低值区出现在岱山岛附近的St5站,最低值为0.371。由数据分析结果及平面分布图可知,该调查海区Shannon-Wiener多样性指数与Pielou均匀度指数出现相似的分布特点,即在浮游植物群落多样性指数高的海区,其均匀度指数也同样较高;而均匀度指数较低的区域,如St5站,浮游植物群落多样性指数亦低。
3 结论
在2011年秋季对中街山列岛邻近海域的海洋调查中,对所获浮游植物样品进行分析,共鉴定浮游植物4门49属127种(不包括未定名物种)。硅藻仍是该海区的主要浮游植物类群,其次为甲藻。物种的生态类型主要可分为广温广盐近岸类群,代表种为角毛藻、角藻、布氏双尾藻、海洋原甲藻、夜光藻等;以及暖温近岸低盐类群,其代表藻种有中肋骨条藻、尖刺拟菱形藻等。此次海洋调查的优势种同以往对该海区的调查结果相比,新增了颤藻、泰晤士扭鞘藻。造成原因或许是因为该片海域的生态环境在一定程度上发生了改变,并进而影响到了浮游植物群落优势种的构成。
调查海域浮游植物的细胞丰度介于0.373×103~199.325×103cell/L,平均丰度值为137.458×103cell/L。与以往对该海域的调查结果相比,此次调查的丰度值略偏低。这可能与本次海洋调查的站点设置覆盖范围较小,加之使用拖网采集样品的方法,丰度值测算结果易造成较大的误差有关,另外,与浮游植物细胞丰度较大的时空变异特性也有着密切关联。调查海域表层细胞丰度呈现在近岸海域数值高,离岸海域数值低的特征。浮游植物群落在垂直方向上的分布特点,大致表现为随海水深度的增加,群落细胞丰度逐渐减小的特点。
调查海域表层浮游植物群Shannon-Wiener多样性指数介于2.594~4.028之间,平均值为3.393。其均匀度指数介于0.371~0.576之间,平均值为0.485。该调查海区Shannon-Wiener多样性指数与Pielou均匀度指数出现相似的分布特点,即在浮游植物群落多样性指数高的海区,其均匀度指数也同样较高;而均匀度指数较低的区域,浮游植物群落多样性指数亦低。
此次对中街山列岛邻近海域的调查,能够为该海域生态环境的恢复提供一定的理论依据和参考数据。但由于此次调查站点设置较少,覆盖面不够全面,以致调查结果有一定局限性,有待在今后对该海域的调查工作中进一步完善与加强。
[1]全为民,沈新强.长江口及邻近水域富营养化现状及变化趋势的评价与分析[J].海洋环境科学,2005,24(3):13-16.
[2]赵卫红,李金涛,王江涛.夏季长江口海域浮游植物营养限制的现场研究[J].海洋环境科学,2004,23(4):1-5.
[3]王云龙,袁 骐,沈新强.长江口及邻近水域春季浮游植物的生态特征[J].中国水产科学,2005,12(3):300-306.
[4]顾新根,袁 骐,杨蕉文.长江口羽状锋海区浮游植物的生态研究[J].中国水产科学,1995,2(1):1-15.
[5]王金辉.长江口邻近水域的赤潮生物[J].海洋环境科学,2002,21(2):37-41.
[6]国家技术监督局.海洋调查规范:海洋生物调查[M].北京:中国标准出版社,1992,17-20.
[7]孙 军,刘东艳.中国海区常见浮游植物种名更改初步意见[J].海洋与湖沼,2002,33(3):271-286.
[8]罗民波,陆健健,王云龙,等.东海浮游植物数量分布与优势种[J].生态学报,2007,27(12):5 076-5 085.
[9]刘子琳,宁修仁,蔡昱明.杭州湾舟山渔场秋季浮游植物现存量和初级生产力[J].海洋学报,2001,23(2):93-99.
[10]徐兆礼,白雪梅,袁 骐.长江口浮游植物的生态研究[J].中国水产科学,1999,5(5):52-54.
[11]王金辉,刘阿成,房建孟.长江河口水域浮游植物动态分布与环境作用[C]//海峡两岸水资源暨环境保护上海论坛论文集.西安:陕西人民出版社,2002.284-292.
[12]顾新根,袁 骐,杨蕉文.长江口羽状锋海区浮游植物的生态研究[J].中国水产科学,1995,2(1):1-15.
[13]洪君超,黄秀清,蒋晓山,等.长江口中肋骨条藻赤潮发生全过程调查报告Ⅰ营养盐状况[J].海洋与湖沼,1994,25(2):179-184.
猜你喜欢
海洋信息技术与应用(2021年3期)2022-01-17
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
河南畜牧兽医(2020年21期)2020-01-10
流行色(2019年10期)2019-12-06
中国农村水利水电(2018年3期)2018-04-13
北京航空航天大学学报(2017年3期)2017-11-23
中国篆刻·书画教育(2017年5期)2017-06-08
纺织检测与标准(2016年3期)2016-12-16
知识窗(2013年7期)2013-05-14
