
云南高原10个湖泊沉水植物的碳、氮、磷化学计量学特征*1
2013-09-25 03:04:50郝贝贝吴昊平刘贵华
湖泊科学 2013年4期
郝贝贝,吴昊平,史 俏,刘贵华,邢 伟**?
(1:中国科学院武汉植物园水生植物与流域生态重点实验室,武汉430074)
(2:中国科学院大学,北京100049)
有机体元素化学特征是有机体漫长演变史的结果,是生态环境条件的统一和反映.有机体对元素的需求特性以及外界环境因素都会影响有机体内部元素化学特征,因此任一元素的缺乏或过量都将导致有机体内部元素比例的失衡,引起有机体生长发育和生态过程的改变[1].生态化学计量学(Ecological stoichiometry)综合了生物学、化学和物理学的基本原理,利用生态过程中多重化学元素的平衡关系,为研究碳、氮、磷等元素在生态系统过程中的耦合关系(主要是C、N、P元素的比例关系)提供了一种综合方法[2-3].
目前,生态化学计量学已经广泛应用于种群动态、生物体营养动态、微生物营养、寄主 病原关系、生物共生关系、消费者驱动的养分循环、限制性元素的判断、生态系统比较分析和森林演替与衰退及全球C、N、P生物地球化学循环等研究中,并取得了许多研究成果[4-6].陆地和水域生态系统都有涉及,研究对象有森林、草地、水蚤、藻类、湿地植物等,但是对于国内湖泊沉水植物的生态化学计量学研究却未见报道.
随着社会与经济的发展,湖泊水体N、P浓度增加,富营养化程度加剧,水生植被尤其是沉水植被衰退和消亡的现象在世界范围内普遍出现.Sagrario等对欧洲22个温带湖泊的调查数据表明,当水体N浓度超过1.2 ~2.0 mg/L、P 浓度高于0.1 ~0.2 mg/L 时,沉水植被就会衰退[7];Jeppesen 等的研究也表明当湖泊无机N∶P小于7、TN浓度在0.5~2.0 mg/L时,沉水植物就会彻底消失[8];Chang等对我国云南省14个湖泊的调查研究表明,在可溶性磷(SRP)高于5 μg/L的水域无沉水植物存在[9].国内外大量的湖泊调查研究表明:当湖泊TN、TP浓度增加到一定值时,沉水植被就会退化甚至消亡.那么,富营养化湖泊沉水植物的化学计量学特征是怎样的?富营养化是如何影响沉水植物元素化学计量特征的?沉水植物内部元素化学特征又是如何适应富营养化环境的?
基于上述3个问题,本文以云南高原10个湖泊的沉水植物为实验对象,研究高原富营养化湖泊沉水植物的元素化学计量学特征,探讨沉水植物在富营养化环境下的退化机理和维持机制,以期为高原湖泊沉水植物的元素化学计量学特征以及高原富营养化湖泊沉水植物的恢复提供基础数据.
1 材料与方法
1.1 样品采集与处理
于2010年8月对云南高原的抚仙湖、剑湖、清水海、洱海、阳宗海、西湖、茈碧湖、星云湖、杞麓湖、长桥海10个湖泊进行沉水植物采集,以采集到相对较多的沉水植物物种为目的.采集沉水植物之前,先采集沉水植物所在区域的水样和底泥样.用采水器采集表层和0.5 m处水样,均匀混合后装入500 ml采样瓶中;用彼得森采泥器采集沉积物,拣出植物根系等后,装入密封袋中;水样和底泥样品先保存于便携式冰箱中,带回实验室后立即测定.采集沉水植物地上部分,去除藻类、浮游动物等附着生物并洗净底泥等杂质,为避免腐烂装入利于水分蒸发的布袋中.所有样品贴上标签,记录样品名称、采样地点和时间.调查的云南高原10个湖泊湖沼学特征见表1.

表1 云南高原10个湖泊的湖沼学特征Tab.1 Limnological characteristics of 10 lakes in Yunnan plateau
1.2 碳、氮、磷的测定
水样元素在测定之前,先经0.45 μm醋酸纤维滤膜过滤.将沉水植物和底泥样品分别在烘箱中烘干至恒重,取出研磨成粉状,干燥保存待测[10].沉水植物、底泥和水体样品的总碳(TC)含量直接使用TOC分析仪(德国耶拿Multi N/C 2100)测定.水体总氮(TN)的测定采用紫外分光光度法;水体总磷(TP)的测定采用抗坏血酸 钼酸铵法[11].沉水植物和底泥样品的TN和TP测定前先进行消解,然后采用靛酚蓝比色法和钼锑抗比色法测定全氮和全磷含量[12].
1.3 数据分析
采用Excel 2010和Statistica 8.0对数据进行分析和制图.采用One-Way ANOVA对数据间差异进行显著性比较.采用Spearman秩相关分析方法分析沉水植物化学计量学特征与水体和底泥化学计量学特征的相关关系.
2 结果与分析
2.1 10个湖泊沉水植物物种和环境因子分析
10个湖泊采集到的沉水植物基本上都属于眼子菜科眼子菜属和小二仙草科狐尾藻属.洱海采集到4种沉水植物:光叶眼子菜(Potamogeton lucens)、马来眼子菜(Potamogeton malaianus)、穿叶眼子菜(Potamogeton perfoliatus)、单果眼子菜(Potamogeton acutiflius);茈碧湖1种:穗花狐尾藻(Myriophyllum spicatum);清水海5种:微齿眼子菜(Potamogeton maackianus)、黑藻(Hydrilla verticillata)、穿叶眼子菜、光叶眼子菜、穗花狐尾藻;阳宗海4种:微齿眼子菜、马来眼子菜、蓖齿眼子菜(Potamogeton pectinatus)、穗花狐尾藻;西湖6种:光叶眼子菜、穿叶眼子菜、单果眼子菜、微齿眼子菜、蓖齿眼子菜、穗花狐尾藻;杞麓湖2种:蓖齿眼子菜、穗花狐尾藻;剑湖3种:光叶眼子菜、微齿眼子菜、穗花狐尾藻;星云湖3种:马来眼子菜、蓖齿眼子菜、穗花狐尾藻;长桥海2种:蓖齿眼子菜、光叶眼子菜;抚仙湖4种:穿叶眼子菜、微齿眼子菜、蓖齿眼子菜、穗花狐尾藻.
无论是水体TN还是底泥TN,杞麓湖、星云湖和长桥海都较其他湖泊污染严重,水体和底泥TN平均浓度分别高于4.50 mg/L和4000 mg/kg;清水海水体和底泥的平均TN浓度最低,分别为0.22 mg/L和500 mg/kg(图1).TP浓度在各个湖泊之间的变化较大,茈碧湖和抚仙湖水体平均TP浓度较低,长桥海水体平均TP浓度最高,而星云湖底泥平均TP浓度最高.
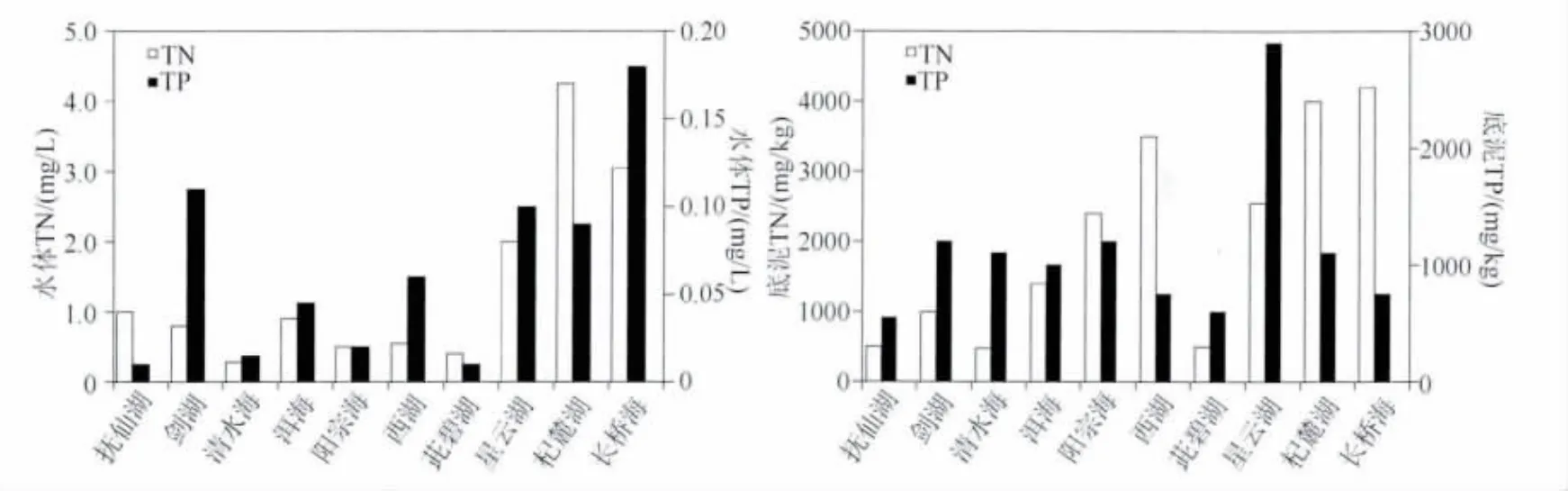
图1 10个湖泊沉水植物采集区域水体和底泥TN、TP浓度Fig.1 Concentrations of total nitrogen and total phosphorous in water and sediment of sampling area in 10 lakes
2.2 10个湖泊沉水植物化学计量学特征及其与环境因子的关系
10个湖泊沉水植物的 TC 含量范围为170.22 ~373.14 mg/g,TN 含量范围为0.90 ~9.70 mg/g,TP 含量范围为0.36 ~4.30 mg/g(图2).其中,星云湖沉水植物的碳、氮、磷平均含量均较高,分别为 361.12、9.70、4.30 mg/g;杞麓湖、抚仙湖沉水植物的碳、氮、磷含量平均值次之;茈碧湖沉水植物的碳、氮、磷平均含量均最低.C∶N、C∶P 和 N∶P 范围分别为 37.21(星云湖)~188.47(茈碧湖)、83.18(杞麓湖)~468.02(茈碧湖)、1.57(杞麓湖)~4.90(清水海).
从单一物种来看,TC、TN、TP最高含量分别出现在马来眼子菜(368.41 mg/g)、光叶眼子菜(7.60 mg/g)和蓖齿眼子菜(3.26 mg/g)(表2).10个湖泊中光叶眼子菜、马来眼子菜、穿叶眼子菜、穗花狐尾藻、微齿眼子菜、蓖齿眼子菜6种沉水植物的平均C∶N、C∶P和N∶P范围分别为40.43(蓖齿眼子菜)~68.93(微齿眼子菜)、93.94(蓖齿眼子菜)~224.57(光叶眼子菜)、2.32(蓖齿眼子菜)~4.82(光叶眼子菜).
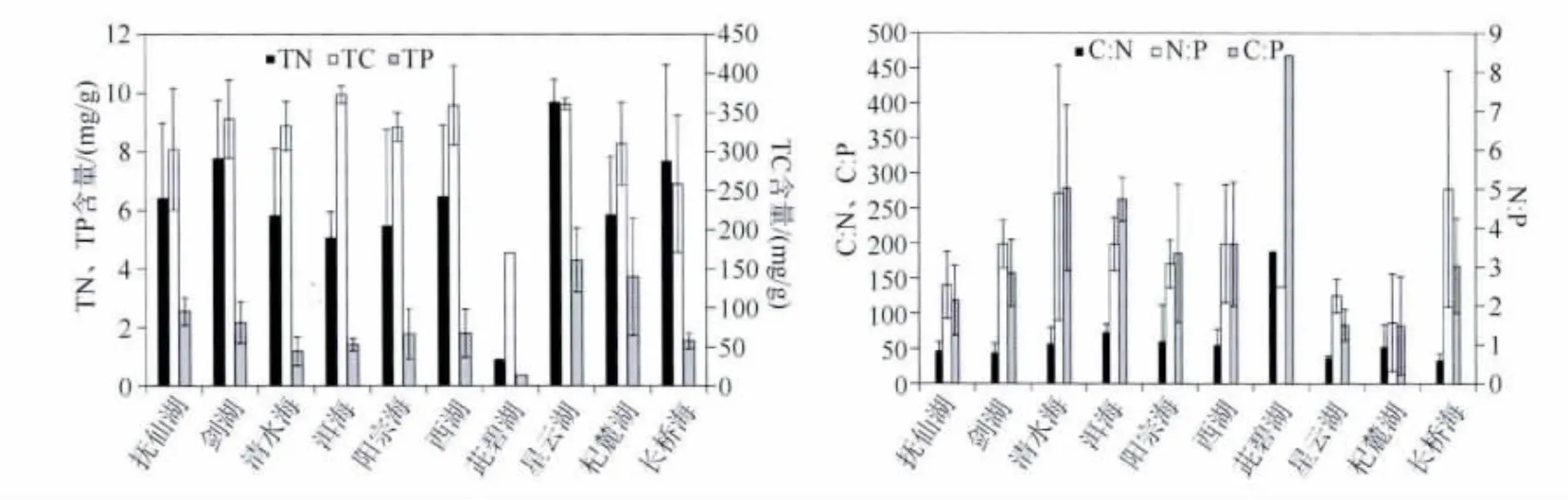
图2 10个湖泊沉水植物碳、氮、磷化学计量学特征Fig.2 Stoichiometric characteristics of C,N and P of submerged macrophytes in 10 lakes
沉水植物碳、氮、磷化学计量学特征与湖泊特征和理化参数的Spearman秩相关分析结果表明(表3),沉水植物TN含量与湖泊最大水深呈显著负相关,与水体TP呈显著正相关(P<0.05);而沉水植物TP含量却与水体TN浓度呈显著正相关(P<0.05);沉水植物C∶P与水体TN呈极显著负相关(P<0.01);C∶N分别与水体TN和TP浓度呈显著负相关(P<0.05);N∶P与湖泊特征和理化参数没有显著相关性(P>0.05).

表2 10个湖泊单一沉水植物物种的碳、氮、磷化学计量学特征Tab.2 Stoichiometric characteristics of C,N and P of submerged macrophytes in 10 lakes

表3 沉水植物碳、氮、磷化学计量学特征与湖泊特征和理化参数的Spearman秩相关分析Tab.3 Spearman rank correlations between stoichiometric characteristics of C,N and P of submerged macrophytes and environmental parameters
3 讨论
在调查的云南高原10个湖泊中,抚仙湖和剑湖为I类水质,洱海和清水海为Ⅱ类水质,阳宗海为Ⅲ类水质,西湖和茈碧湖为Ⅳ类水质,星云湖、杞麓湖和长桥海为劣Ⅴ类水质(表1).沉水植物的物种数随着水质恶化而减少,星云湖、杞麓湖、长桥海沉水植物的物种数仅有2~3种,其他水质较好湖泊则都不低于4种.这些结果说明过高的N、P营养盐严重影响湖泊沉水植物群落的组成结构[8,13].
植物体内的元素含量与许多生物和非生物因子有关,其中最主要的因子是生境条件[6,14-15].沉水植物从水体和沉积物中吸收营养,当水体和沉积物营养浓度增加时,沉水植物体内元素含量也会在一定范围内增加,此时生物体必须改变它们的C∶N∶P比值以适应生长速率的改变[2].富营养化湖泊中沉水植物固然不会受到外界环境N、P营养的限制,但是其体内任一元素的过量也必将通过生长速率理论的控制导致其余元素的消耗,造成元素的相对限制[16-18],从而引发植物体内部元素比例的失衡、动态平衡的打破[19],进而引起生长发育和生态过程的改变[1].而本文在云南高原10个湖泊中调查得到的沉水植物组成结构也恰恰证实了这一点.
植物生物量中的N∶P比值是一个具有重要生态意义的指标,一方面可以反映植被的结构和功能特征,同时也反映群落水平的营养限制状况.当生境中磷过剩而氮不足时,植物过量吸收磷,干生物量中的N∶P比值通常小于14,植物生长表现为氮限制;反之,则植物干生物量中的N∶P比值大于16,植物生长表现为磷限制[10,14].本研究中 10 个湖泊的沉水植物 N∶P 比值平均为 3.5,比梁子湖沉水植物 N∶P 比平均值(5.5)[20]以及洱海沉水植物的N∶P比平均值(7.1)[10]低很多,表明云南高原10个湖泊中沉水植物都是磷过量而氮相对不足,而且这种情况随着水质恶化会更加严重.
Spearman秩相关分析结果表明,植物体N∶P比值与外界环境(水和底泥N、P浓度)没有显著相关性,可能与富营养化湖泊N、P浓度过高有关;而植物体C∶N和C∶P比值分别与水体TP和水体TN呈显著负相关(表3),原因还有待于进一步的实验来解释.研究表明,植物体C∶N和C∶P比值表述了C(生物量)与养分的比值关系,即养分利用效率[21].从单一物种来看,植物体C∶N和C∶P比值最高值分别出现在微齿眼子菜和光叶眼子菜上,表明微齿眼子菜和光叶眼子菜分别对N、P的利用效率高,这与童昌华等[22]的研究结果一致,同时表明微齿眼子菜和光叶眼子菜可能更适合作为云南高原富营养化湖泊沉水植物恢复的物种.
[1]Méndez M,Karlsson PS.Nutrient stoichiometry in Pinguicula vulgaris:nutrient availability,plant size and reproductive status.Ecology,2005,86:982-991.
[2]Sterner RW,Elser JJ.Ecological stoichiometry:the biology of elements from molecules to the biosphere.Princeton:Princeton University Press,2002.
[3]贺金生,韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论.植物生态学报,2010,34:2-6.
[4]曾德慧,陈广生.生态化学计量学:复杂生命系统奥秘的探索.植物生态学报,2005,29:1007-1019.
[5]Sardans J,Rivas-Ubach A,Peñuelas J.The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function:a review and perspectives.Biogeochemistry,2011.DOI 10.1007/s10533-011-9640-9.
[6]Sardans J,Rivas-Ubach A,Peñuelas J.The C∶N∶P stoichiometry of organisms and ecosystems in a changing world:a review and perspectives.Perspectives in Plant Ecology,Evolution and Systematics,2012,14:33-47.
[7]Sagrario G,Mańa A,Jeppesen E et al.Does high nitrogen loading prevents clear-water conditions in shallow lakes at intermediate high phosphorus concentrations.Freshwater Biology,2005,50:27-41.
[8]Jeppesen E,Søndergaard M,Meerhoff M et al.Shallow lake restoration by nutrient loading reduction—some recent findings and challenges ahead.Hydrobiologia,2007,584:239-252.
[9]Chang FY,Pan XJ,Kang LJ et al.Phosphorus bioavailability in relation to submerged macrophyte species and biomass in fourteen temperate lakes,China.Fresenius Environmental Bulletin,2010,19:50-56.
[10]鲁 静,周虹霞,刘贵华等.洱海流域44种湿地植物的氮磷含量特征.生态学报,2011,31(3):709-715.
[11]国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法:第4版.北京:中国环境科学出版社,2006.
[12]董 明,蒋高明,孔繁志等.陆地生物群落调查观测与分析.北京:中国标准出版社,1996.
[13]许秋瑾,金相灿,颜昌宙.中国湖泊水生植被退化现状与对策.生态环境,2006,15(5):1126-1130.
[14]Koerselman W,Meuleman AFM.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation.Journal of Applied Ecology,1996,33:1441-1450.
[15]Demars BOL,Edwards AC.Tissue nutrient concentrations in aquatic macrophytes:comparison across biophysical zones,surface water habitats and plant life forms.Chemistry and Ecology,2008,24:413-422.
[16]Güsewell S,Koerselman W,Verhoeven J.Biomass N∶P ratios as indicators of nutrient limitation for plant populations in wetlands.Ecological Applications,2003,13:372-384.
[17]Güsewell S.N∶P ratios in terrestrial plants:variation and functional significance.New Phytologist,2004,164:243-266.
[18]Yoshida T.Ecological stoichiometry and the shape of resource-based tradeoffs.Oikos,2006,112:406-411.
[19]Schimel DS.All life is chemical.Bioscience,2003,53:521-524.
[20]熊汉锋,黄世宽,陈治平等.梁子湖湿地植物的氮磷积累特征.生态学杂志,2007,26(4):466-470.
[21]Vitouseck PM.Nutrient cycling and nutrient use efficiency.American Naturalist,1982,119:553-572.
[22]童昌华,杨肖娥,濮培民.富营养化水体的水生植物净化试验研究.应用生态学,2004,15(8):1447-1450.
猜你喜欢
绿色天府(2023年5期)2023-06-27 09:26:56
四川畜牧兽医(2023年2期)2023-02-20 02:47:56
云南化工(2021年5期)2021-12-21 07:41:34
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:32:44
现代园艺(2018年1期)2018-03-15 07:56:06
中国草食动物科学(2016年5期)2016-11-24 11:19:13
创新科技(2015年9期)2015-12-15 07:04:09
微型小说选刊(2015年16期)2015-11-17 17:03:05
参花(下)(2015年8期)2015-06-11 10:50:05
小小说月刊(2015年11期)2015-05-14 14:55:32
