
基于FVCOM的太湖梅梁湾夏季水温、溶解氧模拟及其影响机制初探*1
2013-09-25 10:19:04欧阳潇然赵巧华魏瀛珠
湖泊科学 2013年4期
欧阳潇然,赵巧华,魏瀛珠
(南京信息工程大学遥感学院,南京210044)
水温和溶解氧是描述湖泊水生生态系统的两个重要水质因子[1].湖泊水温及其分层现象对湖泊中生物和化学过程有着重要的影响,控制着水体中溶解氧、氮循环、初级生产力等水质参量的变化[2].吴军林等[3]通过实验研究表明,太湖水温达到20℃以后,蓝藻生物量随水温的升高会明显增加;26~28℃是太湖蓝藻生长的最适温度.不同于深水湖泊存在着持续时间长的季节性分层现象[4],浅水湖泊受风场影响混合作用较强,水温分层现象极不稳定[5].对于太湖这样的大型浅水湖泊,张玉超等[6-7]通过长期监测发现太湖水温存在着日成层现象,太阳辐射强度和湖面风速等气象条件是影响日成层产生和强弱的主要因素;分层现象的快速形成与消失对水土界面的物质交换产生影响,从而使得营养盐、藻类浓度的空间分布发生变化[8-10].因此,水温对太湖蓝藻暴发预测预警有着重要的意义.佘丰宁等[11]依据水气界面上的热力方程,建立一维太湖水温数值模型,能够较好地模拟太湖平均水温的日变化.陈黎明等[12]运用澳大利亚西澳大学水研究中心开发的一维DYRESM水动力学模型较好地模拟太湖2005全年的水温变化状况.目前,对太湖水温进行三维模拟研究则较为鲜见.
溶解氧的分布和变化受温度、生物、化学及物理过程的综合影响,总体来说,主要受水温控制[13].在水温的控制下,物理和生化过程对溶解氧的影响各不相同,且相互制约[14].Bierman等[15]利用一个粗尺度的三维水质模型对Mississippi河口的溶解氧收支平衡进行研究,Kemp等[16]对Chesapeake湾的底泥耗氧量和浮游植物呼吸量进行现场观测,结果表明生物化学过程对溶解氧的贡献远大于物理过程.但是Zhang等[17]在研究珠江口溶解氧的动力过程时发现,生物化学和物理过程对溶解氧平衡的贡献相当.目前,对溶解氧影响机制的研究主要集中在深水湖泊以及河口这些水体底层易缺氧的区域,而针对太湖这样的大型浅水湖泊中溶解氧的研究相对较为少见.朱广伟等[18]曾对太湖溶解氧浓度做过为期32天的持续性观测,发现在太湖也存在着缺氧现象.在太湖生态水动力模拟方面,逄勇等[19]对太湖地区的大气 水环境综合数值模拟研究以及刘元波等[20]对梅梁湾藻类生态模拟研究,相对系统地反映了太湖的生态系统动力学变化,由于太湖藻类生物量高,二者的研究均以藻类及其相关的营养盐为主,溶解氧只是作为一个变量参与到生态系统的动力学变化中.吴琼[21]利用WASP 5.0模型模拟了太湖流场、总氮、总磷、溶解氧及浮游植物浓度场,但未对太湖溶解氧浓度的影响机制进行深入的研究.
太湖作为我国目前第三大淡水湖泊,位于长江中下游,对周边地区经济发展具有重要作用,其最大的特点便是“大”而“浅”[22].太湖水体流量小、流速慢、置换周期长、自净能力差[23],水体富营养化导致近年来蓝藻水华频发.2007年5月,太湖蓝藻大规模暴发,在蓝藻腐败分解的过程中,大量消耗水中的溶解氧,导致水体严重污染,发黑发臭,并产生蓝藻腐败的特殊异味,从而引发了自来水污染事件和供水危机[24].特别指出,太湖夏季盛行东南风,外太湖的蓝藻容易聚集在位于太湖北部的梅梁湾,水体富营养化使得蓝藻大量繁殖而引起水华暴发,严重影响周围地区的供水,破坏水体景观,制约周围地区的经济发展[25].因此,有必要对太湖梅梁湾的溶解氧影响机制进行深入的研究,为研究生物 化学过程对水生生态系统的影响以及蓝藻水华暴发机理提供参考.
本研究将随时间变化、空间均匀的实际气象资料作为驱动场,利用FVCOM中水动力模型和水质模型的耦合,对2008年8月16--20日的太湖水温以及各水质参量进行了三维时空模拟;在验证模式有效性的基础上,分析水温的影响因子,并对溶解氧的“源”和“汇”进行定量分析,讨论太湖梅梁湾夏季溶解氧的影响机制.
1 数据与方法
1.1 数据
太湖水域面积2338 km2,平均水深1.89 m,最大水深2.60 m.监测点(见图1)位于太湖梅梁湾东南沿岸的太湖湖泊生态系统研究站(31°25'08″N,120°12'47″E),距岸边150 m.水温记录采用美国 NexSense公司的TS110型水温温度链,温度传感器使用范围为-10~70℃,精度为+/-0.1℃.垂向温度链含有5层温度传感器,深度分别为 0.10、0.63、1.10、1.63、2.13 m.
气象要素观测采用全球湖泊生态观测网络(GLEON)站点Vaisala WXT520自动气象站,其位置在水面上方7 m处,监测指标主要包括风速、风向、气温、相对湿度、气压、降水及短波辐射等.气象数据截取时间为2008年8月16日0:00至8月20日23:00.数据采集频率为10 min/次,本次研究采用整点数据.同时利用邻近气象站点(宜兴市气象站点离湖区12 km)所观测的逐时云量及蒸发量数据.
考虑到太湖面积很大、水体流速较小,且不同湖区承担的功能不同,各湖区的水质情况差异性较大,如若将水质初始场设为常数并对其进行全湖模拟是不符合实际情况的.因此,本研究针对太湖的水质情况将太湖划分为6个区域(图1).水质参量包括溶解氧(DO)、碳化需氧量(CBOD)、浮游生物量(PHYT)、铵态氮(-N)、硝酸态及亚硝酸态氮(-N+-N)、有机氮(ON)、磷酸盐(-P)以及有机磷(OP).其中,溶解氧采样为水下50 cm,其他水质参量上中下三层混合采样;水质初始场数据是由8月15日所采样得到的 1、3、4、5、6、7、10、13、14、16、17、18、31、32号点的水质参量,并且考虑到湖区水质分布特点以及采样点所处地理位置(图1),结合8月16日采样所得的 12、24、25、26、27 号点,插值到网格点上所得;其余的采样点用做检验点.水质采样数据来源于太湖湖泊生态系统研究站;另外,初始场中需要的参数为CBOD,在实际采样过程中所得的参数为化学需氧量(COD),在一些研究[26-28]中认为,CBOD/COD 的比值介于 0.4 ~0.6之间,结合太湖水体的实际特点,CBOD的取值为COD浓度的0.5倍.
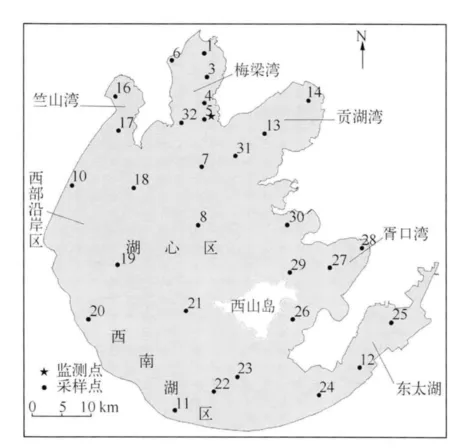
图1 太湖监测点、采样点分布Fig.1 Location of monitoring and sampling sites in Lake Taihu
1.2 方法
为探讨太湖梅梁湾溶解氧的时空变化规律及其影响机制,本次研究利用美国麻省大学海洋科学技术学院海洋生态模型实验室和美国伍兹霍尔海洋研究所于2000年成功建立的非结构网格海洋环流与生态模型——FVCOM模拟太湖水动力情况及溶解氧浓度,代码在http://fvcom.smast.umassd.edu/FVCOM/index.html申请获取.该模式的特点在于[29]:(1)在水平方向上采用无结构三角网格对水平计算区域进行离散,以便更好地拟合边界;(2)在垂向上利用σ坐标系或者S坐标系对不规则的底部地形进行拟合;(3)数值计算采用有限体积积分方式,有限体积法结合了有限元法的自由几何拟合特性和有限差分法的离散结构简单及计算高效的特性,能够更好地保证复杂几何结构的河口海湾及海洋计算中的质量、动量、盐度、温度及热量的守恒性;(4)在物理和数学上使用Mellor-Yamada 2.5阶垂向湍流闭合子模型以及Smagorinsky水平湍流闭合子模型,对方程组进行闭合,且可以将水平方向的湍流扩散系数和垂直方向的扩散系数设为函数,克服了以往太湖水动力学模式中将其设为常数的不足;(5)采用内外膜分离的计算方法,内膜计算与密度场有关的相对较慢的运动,外膜计算与表面重力波相关的快速运动.模式采用三维原始方程,并且包含了多种物理、水质、生态计算模块.FVCOM模型以其在数值处理方法和岸线地形拟合上的优势在河口海岸地带得到了广泛应用[30-32].国内学者将FVCOM模型用于太湖研究并非首次,逄勇等[33]曾经利用FVCOM较好地模拟了太湖流场和悬浮物浓度分布;赵巧华等[34]利用FVCOM对太湖水温以及表层混合层的时空分布进行模拟.本次模拟将太湖划分为19854个三角网格单元和10313个三角网格节点;在垂直方向上采用σ坐标,分为14层;外膜时间步长设为6 s,内膜时间步长设为30 s,积分120 h.公式σ=中,D表示整体水柱深度,H为海底深度(相对于平均海面z=0),ζ为自由面高度(相对于z=0);模式中采用σ坐标系,本文中以sigma 1~sigma 14表示14个σ层.本文数据后期处理使用的是MATLAB的插值及作图函数.
1.2.1 水体所获净热通量计算的改进 根据Churchill等[35-38]计算水气交换过程中水体所获得的净热通量,包括短波辐射、长波辐射、感热及潜热.
1)短波辐射能量计算公式为:式中,Qs0为Vaisala WXT520自动气象站所观测到的短波辐射能量(W/m2),α为水面反照率,Qs为进入水体中的短波辐射(W/m2).

2)长波辐射的计算公式为:

式中,Hlw为长波辐射通量(W/m2);ε为湖水的发射率;K 为斯蒂芬-波尔兹曼参数;Ts和Ta分别为湖面水体温度和大气温度(K);ea为水汽压(hPa);Bc为云的订正因子,Bc=(1-0.72C),其中C为云量(成数).
3)感热和潜热的计算公式为:

式中,Hsen、Hlat分别为感热和潜热通量(W/m2);ρa为大气密度;cpa为大气的热容;Csen为感热转换系数;Clat为潜热转换系数;qa和qsat分别为比湿以及饱和比湿;Le表示蒸发潜热;U为风速.
1.2.2 溶解氧的计算 FVCOM采用的是修改后的WASP 5水质模型,其控制方程参考Zheng等[39]的详细描述.有5个水质参量直接参与到溶解氧的平衡:PHYT、-N、-N+-N、CBOD 和 DO 本身.溶解氧的计算公式为:

式中,C1、C2、C3、C4分别代表着 DO、CBOD、PHYT和-N 浓度(mg/L),T代表水温(℃).方程右侧第1项为大气复氧过程,Kreae为20℃复氧速率,是关于流速和水深的函数;Cs为饱和溶解氧,是关于温度和盐度(在太湖中盐度设为0)的函数.右侧第2项为碳化需氧过程.第3项为硝化作用的耗氧过程.第4项为浮游植物耗氧过程,DPP为浮游植物耗氧率,与浮游植物呼吸率以及死亡率有关,是有关水温的函数.第5项为底泥需氧,SODD为底泥需氧速率.第6项为浮游植物光合作用制氧过程(毛初级生产力模型),GPP为浮游植物生长速率,是关于浮游植物生长率、光照和营养盐限制的函数;Rnc为浮游植物碳氮比;PNH3为氨优先(Ammonia Preference).第7项为细菌呼吸耗氧.参数的取值主要依据实测及WASP 5 模型[40]和相关文献(吴琼[21]、Zheng等[39]),模式通过参数率定及适用性调整.研究中参与到溶解氧平衡的主要参数见表1.由于太湖为藻型湖泊,高等水生植物相对较少,少量的草型区也存在较为严重的退化趋势[41],因而模式中涉及光合作用的过程主要是浮游植物的光合作用及耗氧作用.
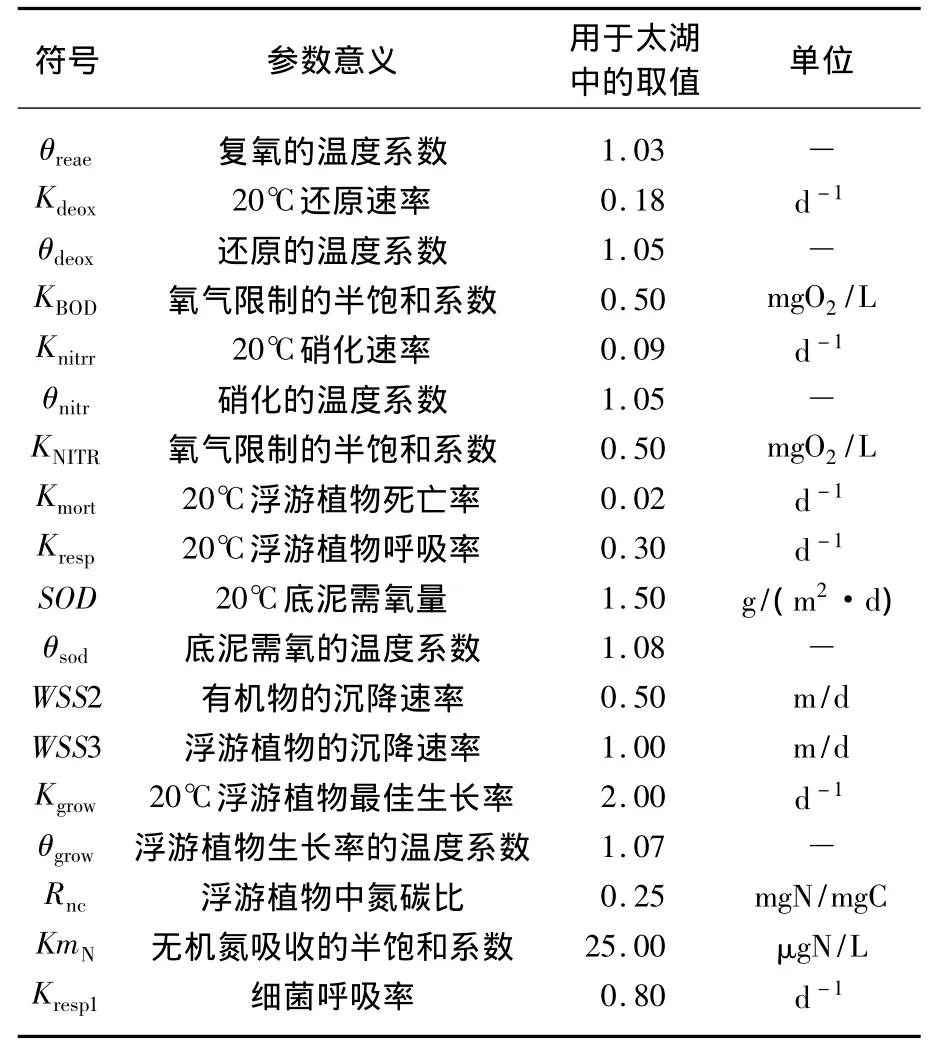
表1 参与溶解氧平衡的参数Tab.1 Parameters participating in the dissolved oxygen balance
2 结果验证与分析
2.1 水温模拟
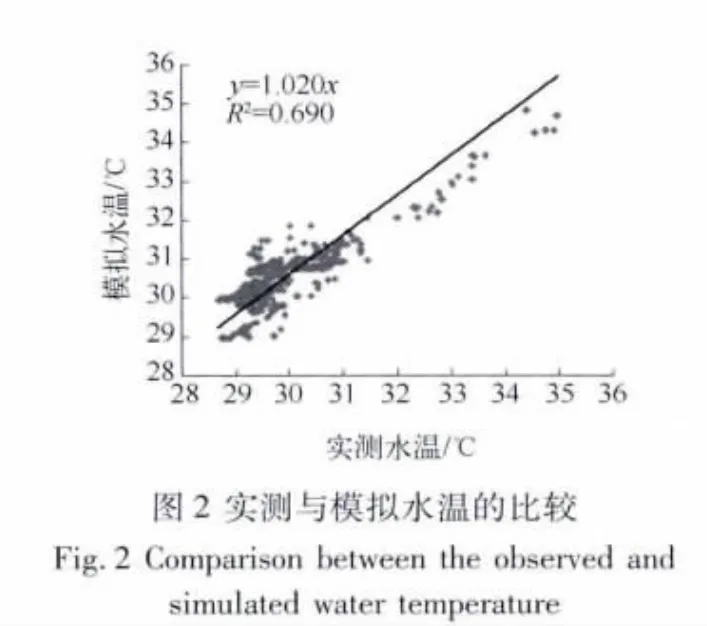
2.1.1 水温模拟结果验证 利用自动气象站所观测到的风速风向、短波辐射、相对湿度、气压及湖边气象站所观测的云量、蒸发量等气象要素作为模式驱动场,模拟太湖三维水温随时间的变化,最后将模拟结果插值到监测点位的相应实测水深处,验证模拟结果以便对太湖水温深度廓线的变化规律进行分析.通过建立相同深度的模拟水温与实测水温散点关系图(图2)可以看出,两者基本紧密散布在y=1.020x的直线附近,R2为0.690(n=480),平均相对误差为1.3%,最大相对误差为6.3%,进而证明了该模式对太湖三维水温有较好的模拟能力.通过水下0.63 m处模拟水温与实测水温的比较可以发现(图3),该模式较好地模拟了太湖水温的日变化趋势,但是存在着一定的误差.误差的主要来源主要可能有两个方面:第一,模式输入所需数据中,气温、短波辐射等气象数据均是整点数据,在两两之间整点时间段内,模式中所用的线性插值并不能完整描述气象场的变化;第二,水体反照率在模式中设定为常数,而反照率受大气浊度和太阳高度角的影响,其中大气浊度通过影响短波辐射中直射与漫射的比例,进而影响反照率;太阳高度角存在明显的日变化,因而反照率也存在日变化,而非常量[34].

2.1.2 水温模拟结果分析 由于太阳辐射强度的变化和水体在水深方向上的增温和冷却强度不一,使得水温呈现出分层现象;水温的状况及其分层现象直接或间接地影响湖库水环境中的各种物理、化学和生物过程[42].从溶解氧的物理过程来看,水体的垂直温度分层直接决定着溶解氧的扩散系数[43];从溶解氧的生物化学过程来看,水温直接控制着溶解氧的“源”和“汇”(公式5).
从8月16日- 8月20日不同σ层模拟水温随时间的变化可以清楚地看出,水温的日变化以表层最为明显,随着深度的加深,日变化逐渐减弱(图4).另外,不同深度处的水温极值出现的时间也不一致,表现出随着深度的加深最高温度出现的时间逐渐滞后:表层最高温度一般出现在14:00--15:00,随着深度的增加,在湖面以下1.5 m处,最高温度出现在傍晚18:00--19:00.这是由于水温的变化主要取决于热量收支各要素间的平衡,热量由湖面向湖底传递需要时间.同时夜间水温随深度变化不大、基本相同,这说明夜间水体基本混合均匀,而白天水温最大相差4℃.这说明太湖这样的大型浅水湖泊水温分层现象在白天可能存在,但是在夜间基本消失、水体混合基本均匀.
2.2 溶解氧模拟
2.2.1 溶解氧模拟结果验证 利用8月15日所有采样点以及8月16日部分采样点进行插值作为水质参量的初始场,通过参数率定,最后将模拟结果插值到剩余采样点水下50 cm特定采样时间上,验证溶解氧模拟结果(表2).溶解氧模拟值与采样实测值相近,判断系数R2为0.76,最大相对误差为12.88%,平均相对误差为3.53%,证明该水质模型能够较好地模拟溶解氧.同时,将溶解氧30个采样点进行空间插值与8月20日23:00的最终模拟结果进行比较(图5),可以看出两者在空间分布趋势上基本一致,溶解氧浓度总体上表现出由东向西减少的趋势.梅梁湾属于藻型湖湾,浮游植物生物量较大,在朱广伟等[18]对太湖为期32 d的研究中,有18 d梅梁湾监测点处溶解氧的浓度大于12 mg/L,远远超过了溶解氧在特定水温下的饱和度(水温 30℃,101.325 kPa,纯水中溶解氧的饱和溶解度为7.55 mg/L);东太湖为草型湖区,分布着大量沉水植物,采样期间,东太湖25#采样点的叶绿素a 浓度(58.03 μg/L)高于往常,浮游植物光合作用制氧量较大,因此造成该采样点在模拟时间段内溶解氧浓度较高.
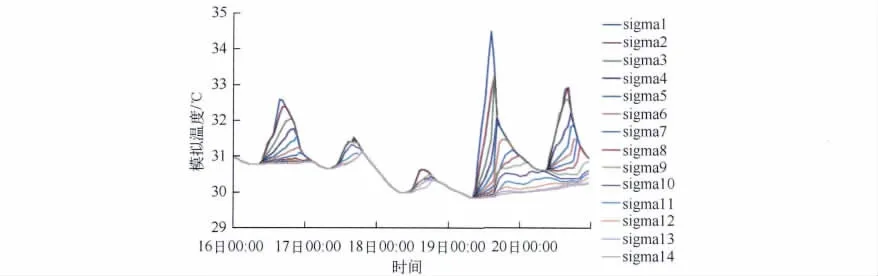
图4 不同深度模拟水温随时间的变化Fig.4 The simulated temperature at different depths varying from hours
2.2.2 溶解氧模拟结果分析 以8月20日为例,将溶解氧模拟值插值到梅梁湾处的监测点上,并求取水柱层平均溶解氧浓度,溶解氧浓度随着时间变化大致呈现出“双峰双谷”的趋势(图6).水柱层中溶解氧浓度表现出了垂直差异(图6a),这可能是由水温以及光合作用产氧量的垂直差异造成的.其一,结合图4可以发现,8月20日日出前与日落后,水温有着不大明显的垂直分层,日出后至日落前这段时间的垂直分层现象较为明显;在图6a中,溶解氧浓度也呈现出与水温相似的垂直分层现象.由公式(5)可知,水温控制着溶解氧的各个过程,进而影响着溶解氧的变化.Vassilis等[1]和张玉超等[6]研究也认为水温分层对湖泊中的生物化学过程有着重要的影响,水温分层现象的存在不利于表层溶解氧向下传输,溶解氧浓度表现出随着深度越深逐渐下降的现象.其二,由于光线的水下衰减引起光合作用的垂直分层,可能也会引起溶解氧的垂直分层现象,这将在下文讨论中详细解释.同时表层与底层溶解氧浓度最大差值仅为2 mg/L左右(图6a),水体并没有像其他深水湖泊一样表现出底层缺氧的现象,这可能是由于水深较浅,光线能够较好地进入水下,有利于水下浮游植物光合作用的顺利进行.秦伯强等[44]的研究也表明,像太湖这样的浅水湖泊,水土界面上的溶解氧供应充分,水体溶解氧浓度随深度的变化很小.

表2 太湖溶解氧模拟结果验证Tab.2 The validation of the simulated dissolved oxygen in Lake Taihu

图5 溶解氧浓度实测值(a)和模拟值(b)空间分布趋势Fig.5 The spatial distribution of observed(a)and simulated(b)dissolved oxygen
梅梁湾溶解氧浓度呈现出“双峰双谷”的趋势明显(图6b).极小值出现在日出前4:00左右和太阳辐射最为强烈的午后14:00左右.这主要是因为夜间没有光合作用,而浮游植物以及其他无机物和有机物消耗了溶解氧,使得日出前溶解氧浓度达到极小值.午后由于光强较大,光抑制现象导致上层水体光合作用产氧量在午后达到最低;同时由于水温的不断升高,此时浮游植物的呼吸率和死亡率随之升高(公式5),水体中溶解氧的消耗在此时段达到一天中的最大值;并且此时Cs逐渐减小,水体中的溶解氧不断向大气逸散,上述原因使得午后溶解氧的浓度达到另一个极小值.极大值出现在日出后9:00以及日落时18:00左右,这是因为日出后随着光照增强,光合作用也随之增强,但中午前后光照最强产生光抑制现象,午后随着光强减弱至适宜光合作用发生,达到一天中的第2次最大值.上述分析表明,水温和光照可能是影响溶解氧浓度变化的重要因子.

图6 8月20日监测点溶解氧的变化趋势(a:垂直分布,b:水柱层平均)Fig.6 The change trend of dissolved oxygen at the monitoring site on August 20th(a:vertical distribution,b:average of the water column)
朱广伟等[18]通过对梅梁湾监测点处溶解氧浓度进行连续观测发现,其浓度呈现出“单峰单谷”的变化趋势,谷值出现在日出前4:00 -7:00之间,峰值出现在14:00--18:00之间.本文分析结果与其观测结果部分相似,即其中一个谷值和一个峰值出现的时间是一致的,但是朱广伟等的研究中并没有在午后出现另一个谷值,这可能是由当时的主导风向、风速及气温等气象条件和藻类等水质参量浓度的不同所引起的.关于气象条件对溶解氧的影响将在今后进行进一步的探讨.
3 讨论
3.1 水温的影响因子
赵林林等[4]和张玉超等[6]认为,影响太湖水温日成层现象的因素主要有太阳辐射、气温、风速.赵巧华等[34]认为,水温的分层及日变化主要取决于水气热量交换过程中水体所获得的净热通量;当净热通量为正时,水体吸收热量致水温上升.因此,这里讨论太阳辐射、风速对水温日成层现象的影响.
其一,太阳辐射.结合图4,太阳辐射与水温表现出相似的日变化.这是因为随着太阳辐射的逐渐增强至约700 W/m2,表层水体不断吸收太阳辐射,水温变化较为剧烈,衰减作用使得底层水温变化较小.通过实测的太阳辐射数据与模拟的水温相比较,可以发现这与前人的研究结果[4,6]一致.同时发现,太阳辐射强的时候(8月18日中午),其水温(30.5℃)不一定大于太阳辐射相对较弱时候(8月16日中午)的水温(33℃).这可能与当日的水-气热量交换及风速引起的动力作用有关,8月18日最高气温(29℃)低于8月6日最高气温(32℃),并且8月18日的风速较8月16日相对剧烈(图7a).
其二,风速.8月19日、8月20日的气温极值以及太阳辐射与8月17日相当(图7a、图7b),但是水体温度分层则表现出较大的差异,并且水温极值的大小差异也较大;其中8月17日水温较低且分层现象不明显,另8月17日风速较大(图7c).因而8月17日水温分布成因可能是由风场造成的水体流速切变引发的不稳定作用所致.在风速较大的情况下(8月17日、8月18日),由于风的驱动作用使得水体水平运动垂直切变大于温度的垂直梯度,加强水体的混合作用,上下层的热量交换加速,水温的垂向差异变得很小,造成白天水体分层现象难以形成,中午11:00后才出现微弱的温差,直至傍晚18:00左右,此时的太阳辐射已经十分微弱,分层现象消失.
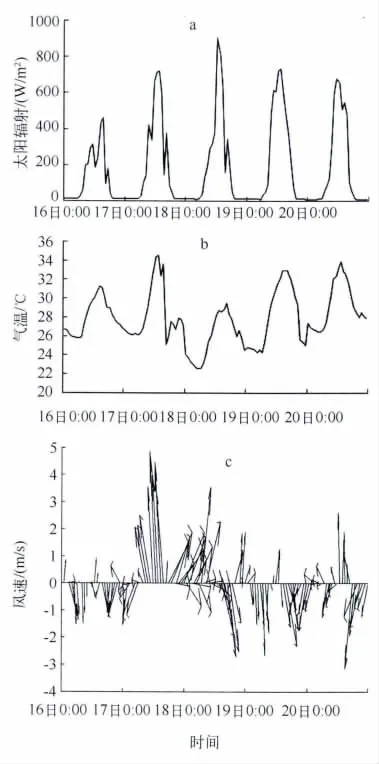
图7 模拟时间段内实际太阳辐射(a)、气温(b)和风速(c)变化Fig.7 Solar radiation(a),air temperature(b)and wind speed(c)during simulated hours
3.2 溶解氧的可能影响机制
在对太湖水动力及溶解氧模拟结果进行验证和分析后,尝试着主要对梅梁湾溶解氧的“源”、“汇”进行探讨.如公式(5)所示,直接影响溶解氧生物-化学机制的动力过程主要有7个:复氧、碳化需氧、硝化作用耗氧、浮游植物耗氧、底泥耗氧、浮游植物制氧以及细菌呼吸耗氧过程.考虑到模式的稳定时间,水质模型的结果从2008年8月16日6:00开始输出,本文选取2008年8月17日- 8月20日的结果对梅梁湾太湖湖泊生态系统研究站监测点进行讨论.
浮游植物光合作用制氧通常用来作为评估初级生产力(这里指毛初级生产力)的依据,很多学者[45-46]通过野外采样和实验室模拟分析初级生产力的日变化和垂直分布.依据公式(5)中的第6项浮游植物光合作用制氧来计算毛生产力.光合作用在白天进行,刚开始随着光照作用增强,初级生产力增大;中午前后光抑制现象发生,表层初级生产力急剧下降,次表层(水下20~40 cm)初级生产力达到最大值;随着光照减弱,表层初级生产力逐渐升高(图8).从垂直方向看,除了表层会受到光抑制作用的影响外,初级生产力随着深度越深而下降,上层初级生产力显著高于下层,在底层达到最小值.此外,在8月17日风浪较大的时候,初级生产力高于风浪较小的时候.从数量级上看,初级生产力的范围大概在0.05~0.85 mg/(L·h),上层主要集中在0.3~0.4 mg/(L·h)之间;若以日为单位,水柱毛初级生产力大约为 1.4 mg/(L·d),这与蔡后建等[45]以及张运林等[46]关于太湖初级生产力的研究结果是一致的.可以发现,光合作用是导致溶解氧出现日变化的主要因素.
若水体中的氧气低于饱和值,可以通过大气复氧来补充[40].复氧系数是关于风速、气温、水温和水深的函数[47].当水温较高(8月19日、8月20日)时,这种复氧作用表现的尤为明显.由公式(5)可知,复氧过程对溶解氧的贡献的正负主要取决于湖泊中溶解氧的饱和度,即水中溶解氧C1与该水温下饱和溶解氧Cs的差值.若C1>Cs,则复氧过程的作用为负(图9,以监测点为例),即此时水体中的溶解氧处于向大气逸散的状态;若C1<Cs,则复氧过程的贡献为正,大气补充水体中的氧气.

图8 各层初级生产力随时间的变化Fig.8 The primary productivity at each depth varying with hours

图9 监测点处复氧作用(a)和主要生物化学过程(b)的贡献Fig.9 The contribution of reparation(a)and main bio-chemical processes(b)at the monitoring point
耗氧过程包括5个:浮游植物耗氧、碳化需氧、硝化作用耗氧、底泥耗氧、细菌呼吸耗氧.以监测点整个水柱层平均为例,浮游植物是梅梁湾处溶解氧最大的消耗者(包括呼吸和死亡过程),其贡献占到了除去复氧过程的总耗氧量的76.44%(8 月 17 日)、74.74%(8 月18日)、69.22%(8 月 19 日)和 68.38%(8 月20日),在数量级上和以前的研究[45]是一致的;其次分别为底泥耗氧(多日平均为10%)、碳化需氧(8%)、细菌呼吸耗氧(5%)以及硝化作用耗氧(3%)(图9b).另外可以发现,光合作用制氧与各耗氧过程(除去由湖面向大气逸散的溶解氧部分)大致可以平衡,也就是说,生化过程是影响太湖梅梁湾溶解氧浓度的主要动力因子;这一点与学者对Mississippi河口[15]以及 Chesapeake湾[16]的研究结果一致,但与张恒等[17]在珠江河口的结论存在明显差异,其原因可以从水体滞留时间上进行考虑.太湖水体流速小,水体置换周期长(约为250 d),有利于浮游植物繁殖以及C、N、P循环;相较于Mississippi河口(密度层以下水体置换周期为95 d)、Chesapeake湾(水体置换周期为180 d)以及太湖,珠江河口的水体置换周期很小(在汛期只有3~5 d),不适宜浮游植物大量繁殖.
在风浪较大的时候(8月17日),各项耗氧过程在数值上都大大增加(图9b).Kristensen等的研究[48]也表明,风浪扰动时浅水湖泊沉积物悬浮和营养盐释放的主要驱动力之一,半天的强风浪扰动可导致水体营养盐浓度增加近一倍.以往的研究[49]表明,营养盐是浮游植物生长的主要控制因子.可以推测,在大风浪初期,如果有足够的营养盐,藻类植物大量繁殖,水中溶解氧浓度会增加;随后,大风若持续进行,底泥营养盐不断释放,藻类植物大量繁殖,呼吸作用增加了对水中溶解氧的消耗,并且大量藻类植物死亡又需要消耗大量的氧气;大风浪后风平浪静期间,藻类植物易聚集在水面上,阻碍了阳光进入湖水中,不利于光合作用的进行,造成水体缺氧.
4 结论与展望
本文利用耦合了水动力和水质模块的FVCOM模式,较好地模拟了太湖水温以及溶解氧的日变化和垂直分布.通过分析和讨论认为,太阳辐射、风速是影响水温分层以及大小的主要因子;受水温和光照的影响,夏季梅梁湾的溶解氧存在垂直差异,呈现出“双峰双谷”的日变化特征;水下光衰减影响着水体浮游植物光合作用,使之呈现出垂向变化.在模拟时段内,梅梁湾地区监测点的溶解氧处于过饱和状态,复氧过程对溶解氧浓度的贡献为负;在影响太湖梅梁湾处的溶解氧生物-化学过程中,浮游植物光合作用制氧以及耗氧(包括呼吸和死亡作用)起着决定性的作用;在其他耗氧过程中,对溶解氧的贡献依次为底泥耗氧、碳化需氧、细菌呼吸耗氧、硝化作用耗氧.
同时在模式模拟的过程中存在以下缺陷:(1)模式中对逐时气象数据采用的是线性插值方法,不能完整刻画气象场的变化;(2)水质模型中只考虑了浮游植物的光合作用,而忽略了水生植物的呼吸与光合作用;(3)由于条件限制,水质模型中的参数值是在参考经验值的基础上通过程序调试完成,误差难以避免.这些缺陷将在今后的研究中予以重视.
致谢:文中所用水质采样数据以及逐10 min的气象及水温监测数据均来源于中国科学院太湖生态网络站,逐时云量及蒸发量数据来源于宜兴市气象局,在此一并表示感谢.
[1]Vassilis ZA,Soultana KG.Simulation of water temperature and dissolved oxygen distribution in Lake Vegoritis,Greece.Ecological Modelling,2003,160:39-53.
[2]Stefan HG,Hondzon M,Fang X.Lake water quality modeling for projected future climate scenarios.Journal of Environmental Quality,1993,22(3):417-431.
[3]吴军林,王建萍.太湖底泥中蓝藻复苏的温度阈值研究.绿色中国,2006,(13):68-69.
[4]赵林林,朱广伟,陈元芳等.太湖水体垂向分层特征及其影响因素.水科学进展,2011,22(6):844-850.
[5]孙顺才,黄漪平.太湖.北京:海洋出版社,1993.
[6]张玉超,钱 新,钱 瑜等.太湖水温分层现象的监测与分析.环境科学与管理,2008,33(6):117-121.
[7]张玉超,钱 新,石川忠晴等.浅水湖泊水温日成层现象的初步探讨——以太湖为例.四川环境,2008,27(3):45-48.
[8]秦伯强,胡维平,高 光等.太湖沉积物悬浮的动力机制及内源释放的概念性模式.科学通报,2003,48(17):1822-1831.
[9]逄 勇,韩 涛,李一平等.太湖底泥营养要素动态释放模拟和模型计算.环境科学,2007,28(9):1960-1964.
[10]Lovestedt CB,Bengtsson L.The role of non-prevailing wind direction on suspension and redistribution of sediments in a shallow lake.Aquatic Science,2008,70:304-313.
[11]佘丰宁,蔡启铭,徐勇积.太湖水温模型和气象参数对水温的影响.海洋与湖沼,1993,24(4):393-399.
[12]陈黎明,钱 新,杨 珏等.基于DYRESM模型的太湖全年水温模拟及其在水华预警中的应用.环境保护科学,2009,35(2):18-21.
[13]张竹琦.渤海、黄海(34°N以北)溶解氧年变化特征及与水温的关系.海洋通报,1992,11(5):41-45.
[14]张 恒.珠江河口夏季溶解氧收支模拟研究[学位论文].广州:中山大学,2009.
[15]Bierman VJ,Hinz SC,Zhu DW et al.A preliminary mass balance model of primary productivity and dissolved oxygen in the Mississippi River Plume:Inner Gulf Shelf Region.Estuaries,1994,7(4):886-899.
[16]Kemp WM,Sampou PA,Garber J et al.Seasonal depletion of oxygen from bottom waters of Chesapeake Bay:roles of benthic and planktonic respiration and physical exchange processes.Marine Ecology Progress Series,1992,85:137-152.
[17]Zhang H,Li SY.Effects of physical and biochemical processes on the dissolved oxygen budget for the Pearl River duriner.Journal of Marine System,2010,79(1/2):65-88.
[18]Zhu GW,Wang F,Zhang Y et al.Hypoxia and its environmental influences in large,shallow,and eutrophic Lake Taihu,China.Inland Waters:Journal of the International Society of Limology,2008,30(3):361-365.
[19]逄 勇,姚 琪,濮培民.太湖地区大气 水环境的综合数值研究.北京:气象出版社,1998.
[20]刘元波,陈伟民,范成新.太湖梅梁湾藻类生态模拟与蓝藻水华治理对策分析.湖泊科学,1998,10(4):53-59.
[21]吴 琼.大型浅水湖泊生态动力学特性的数值研究[学位论文].南京:河海大学,2007.
[22]秦伯强.太湖水环境面临的主要问题、研究动态与初步进展.湖泊科学,1998,10(4):1-9.
[23]许旭峰,刘青泉.太湖风生流特征的数值模拟研究.水动力研究与进展:A辑,2009,24(4):512-518.
[24]夏 健,钱培东,朱 玮.2007年太湖蓝藻水华提前暴发气象成因探讨.气象科学,2009,29(4):531-535.
[25]陈宇炜,秦伯强,高锡云.太湖梅梁湾藻类及其相关因子逐步回归统计和蓝藻水华的初步预测.湖泊科学,2001,13(1):63-71.
[26]Cao WH,Mehrab M.Slaughterhouse wastewater treatment by combined anaerobic baffled reactor and UV/H2O2processes.Chemical Engineering Research and Design,2011,89(7):1136-1143.
[27]Prakash RK,Lee S,Lee YS et al.Application of automated QUAL2Kw for water quality modeling and management in the Bagmati River,Nepal.Ecological Modelling,2007,202(3/4):503-517.
[28]Prakash RK,Seockheon L,Sushil RK et al.Spatial-temporal variation,and comparative assessment of water qualities of urban river system:a case study of the river Bagmati(Nepal).Environmental Monitoring and Assessment,2007,129(1/2/3):433-459.
[29]Chen CS,Robert CB,Geoffrey C.An unstructured grid,finite-volume coastal ocean model,FVCOM User Manual.2006.
[30]Chen C,Huang H,Robert CB et al.Tidal dynamics in the Gulf of Maine and New England Shelf:An application of FVCOM.Journal of Geophysical Research,2011,116:C12010.
[31]Qi JH,Chen CS,Robert CB et al.An unstructured-grid finite-volume surface wave model(FVCOM-SWAVE):Implementation,validations and applications.Ocean Modelling,2009,28(1/2/3):153-166.
[32]Jennifer AS.Modelling the circulation and exchange of Kingston Basin and Lake Ontario with FVCOM.Ocean Modelling,2009,30(2/3):106-114.
[33]逄 勇,庄 巍,韩 涛等.风浪扰动下的太湖悬浮物实验与模拟.环境科学,2008,29(10):2743-2748.
[34]Zhao QH,Sun JH,Zhu GW.Simulation and exploration of the mechanisms underlying the spatiotemporal distribution od surface mixed layer depth in a large shallow lake.Advanced in Atmospheric Sciences,2012,29(6):1360-1373.
[35]Churchill JH,Kerfoot WC.The impact of surface heat flux and wind on thermal stratification in Portage lake,Michigan.Journal of Great Lake Research,2007,33:143-155.
[36]Kim TA,Cho YK.Calculation of heat flux in a macrotidal flat using FVCOM.Journal of Geophysical Research,2011,116:C03010.
[37]Maggiore A,Zavatarelli M,Angelucci MG et al.Surface heat and water fluxes in the Adriatic sea:seasonal and interannual variability.Physics and Chemistry of the Earth,1998,23(5):561-567.
[38]Gill AE.Atmosphere-ocean dynamics.UK:Academic Press,1982:662.
[39]Zheng LY,Chen CS,Zhang FY.Development of water quality in the Satilla River Estuary,Georgia.Ecological Modelling,2004,178:457-482.
[40]Ambrose RB,Wool TA,Martin JL et al.The water quality analysis simulation program,WASP5,Part A:Model documentation.US Environmental Protection Agency Athens Georgia,1993.
[41]雷泽湘,徐德兰,黄沛生等.太湖沉水和浮叶植被及其水环境效应研究.生态环境,2006,15(2):239-243.
[42]Lin J,Xie L,Pietrafesa LJ et al.Dissolved oxygen stratification in two micro-tidal partially-mixed estuaries.Estuaries Coastal and Shelf Science,2006,70(3):423-437.
[43]David AB,Asce AM.Dissolved oxygen variations in stratified lakes.Journal of the Sanitary Engineering Division,1970,95(5):1129-1146.
[44]秦伯强,范成新.大型浅水湖泊内源营养盐释放的概念性模式探讨.中国环境科学,2002,22(2):150-153.
[45]蔡后建,陈宇伟,蔡启铭等.太湖梅梁湾口浮游植物初级生产力及其相关因素关系的研究.湖泊科学,1994,6(4):340-348.
[46]张运林,秦伯强,陈伟民等.太湖梅梁湾浮游植物叶绿素a和初级生产力.应用生态学报,2004,15(11):2127-2131.
[47]O'Connor DJ.Wind effects on gas-liquid transfer coefficients.Journal of Environmental Engineering,1983,109(9):731-752.
[48]Kristensen P,Søndergaard M,Jeppensen E.Resuspension in a shallow eutrophic lake.Hydrobiologia,1992,228:101-109.
[49]杨东方,陈生涛,胡 均等.光照、水温和营养盐对浮游植物生长重要影响大小的顺序.海洋环境科学,2007,26(3):201-206.
猜你喜欢
家庭科学·新健康(2020年7期)2020-08-06 08:15:35
科学与信息化(2020年11期)2020-06-19 08:50:42
南方农业·中旬(2020年2期)2020-05-28 13:47:28
玩具世界(2017年4期)2017-07-21 13:27:24
计算机测量与控制(2017年6期)2017-07-01 16:24:28
中国海洋大学学报(自然科学版)(2017年7期)2017-06-05 15:08:52
水利科技与经济(2017年6期)2017-04-28 08:30:14
宝藏(2017年2期)2017-03-20 13:16:42
中亚信息(2016年3期)2016-12-01 06:08:24
金融经济(2015年16期)2015-08-21 11:23:58
