
不同硒源及水平对蛋用种公鸡肝脏中硒含量、抗氧化性及基因表达的影响
2013-09-20 03:04:52陈秀芸杨佐君王晓霞杨开伦
动物营养学报 2013年9期
陈秀芸 滑 静 杨佐君 王晓霞 杨开伦 张 洁
(1.新疆农业大学动物科学学院,乌鲁木齐 830052;2.北京农学院动物科学技术学院,北京 102206;3.北京农学院生物技术学院,北京 102206;4.北京市昌平区区委,北京 102200;5.北京农业职业技术学院畜牧兽医系,北京 102442)
肝脏中有含量丰富的微量元素,因此有“微量元素之库”之称。通常情况下肝脏中的微量矿物元素比较稳定,所以肝脏中微量矿物元素的含量能够反映机体所在环境中的微量元素的情况[1]。现有研究表明,硒是具有重大生物学意义的微量元素之一,它与动物的生长发育、繁殖和疾病发生有着密切的关系,并且硒具有抗氧化、提高机体免疫力和抗病力、调节机体代谢、拮抗有毒元素等多种功能[2-3]。近年来在动物生产应用中,无机硒亚硒酸钠(sodium selenite,SS)正逐步被有机硒取代,与无机硒相比,有机硒在动物体内具有吸收率高、生物活性强、毒性低、环境污染小等特点;并且可以通过提高动物机体内硒沉积量,发挥更高的抗氧化性能,以提高动物机体对疾病的抵抗力[4-6]。壳聚糖为甲壳素的脱乙酰化产物,广泛存在于虾、蟹和昆虫的外壳及真菌、藻类的细胞壁中;壳聚糖具有调节免疫、抗癌、抗病毒等多种功能,而且无毒、无副作用,对环境无公害并且可生物降解,其分子结构上有多种活性基团,使其具有很好的螯合性,易于进行化合。我国有72%国土为不同程度的缺硒区[7-8],2/3以上地区的饲料和饲料牧草硒含量小于0.05 mg/kg[9],同时我国家禽饲料的大部分原料是以玉米-豆粕型为主,通常情况下难以达到其对硒的需要量,因此必然要在饲粮中补硒,以满足家禽的正常需要。人工合成的有机硒产品很多,如硒蛋氨酸、硒化卡拉胶、硒化脂多糖等,这些产品生产工艺比较复杂,价格昂贵,实验室研究报道较多,而在实际生产应用中很少。显然,寻求低毒、廉价、生物可利用性良好及生产工艺简单的硒化物对于进一步提高家禽的抗氧化性及抵抗力等性能是十分重要的。作为一种安全、廉价、低污染、高生物利用率的有机硒——硒化壳聚糖(seleno-chitosan,SC),其对蛋用种公鸡肝脏中硒含量及其抗氧化性的影响还未见相关报道。本文旨在通过在海兰褐蛋用种公鸡饲粮中分别添加不同水平的有机硒(硒化壳聚糖)和无机硒(亚硒酸钠),研究其对蛋用种公鸡肝脏中硒含量、抗氧化性、硒酶活性及谷胱甘肽过氧化物酶4(GPx4)基因mRNA表达水平的影响,以期为其在家禽生产中的应用提供科学理论依据。
1 材料与方法
1.1 试验材料
试验所用的硒化壳聚糖是由无机硒与壳聚糖通过化学的方法稳定地相结合得到的有机硒源,由青岛农业大学饲料研究所研制,硒含量为11.38%。亚硒酸钠购自黄骅市津华添加剂有限公司,硒含量为44.7%。
1.2 试验动物及试验设计
选用196只150日龄健康的配种前期海兰褐蛋用种公鸡,平均体重为(2.112±0.034)kg。试验采用单因子试验设计,随机分为7组,每组4个重复,每个重复7只,各组初始体重差异不显著(P>0.05)。对照组饲喂基础饲粮,试验组分别饲喂在基础饲粮中添加0.4、0.8和1.2 mg/kg硒化壳聚糖和亚硒酸钠2种硒源的试验饲粮。所有组均饲喂基础饲粮1周后,开始饲喂相应的试验饲粮。试验期35 d。
饲养试验在北京农业职业技术学院种鸡场进行。试验鸡单笼饲养,自由采食和饮水,人工喂料,每天按规程要求清扫地面、清粪和带鸡喷雾消毒。每天观察鸡群健康状况,并记录试验数据。每周进行1次结料,准确称取饲料重,记录各重复蛋用种公鸡的耗料量;于试验第35天进行空腹称重,计算各重复的平均日增重(ADG)、平均日采食量(ADFI)和料重比(F/G)。
1.3 试验饲粮
依据NRC(1994)标准配制玉米-豆粕型基础饲粮(粉料),基础饲粮组成及营养水平见表1。

表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet(air-dry basis) %
1.4 组织取样及组织匀浆的制备
试验结束后,每个重复挑选1只鸡,取肝脏称重,放入液氮罐,随后移至-80℃冰箱保存,以备用。
组织匀浆上清液的制备:肝脏样品用冷生理盐水清洗,剔除结缔组织、脂肪,滤纸吸干,分别取0.5 g左右于玻璃匀浆器中,加5 mL冷的生理盐水匀浆,匀浆后于4℃条件下10 000 r/min离心25 min,取上清液。
1.5 指标测定
硒含量测定:按照GB/T 12399—1996的方法测定血浆及肝脏中的硒含量,由北京华英生物技术研究所检测。
肝脏抗氧化性测定:谷胱甘肽过氧化物酶(GSH-Px)活性、总超氧化物歧化酶(T-SOD)活性、过氧化氢酶(CAT)活性、总抗氧化能力(T-AOC)、丙二醛(MDA)含量均采用南京建成生物工程研究所提供的鸡专用试剂盒,具体步骤按照试剂盒说明进行,用紫外分光光度计测定。
肝脏硒酶活性测定:Ⅰ型脱碘酶(IDⅠ)和硫氧还蛋白还原酶(TrxR)活性检测采用放免法,由北京华英生物技术研究所检测。
1.6 GPx4基因mRNA表达水平的测定
1.6.1 引物设计与合成
根据GenBank公布的鸡基因序列设计引物如表2,引物由上海生物工程技术服务有限公司合成。

表2 RT-PCR引物序列Table 2 Primer sequences for RT-PCR
1.6.2 组织样本总RNA的提取和反转录
用超纯RNA提取试剂盒(Omega)提取肝脏组织样本中总RNA。试验操作按产品说明书进行,并对所提取的RNA进行质量检测,判断有无污染。cDNA合成用MMLV cDNA第1链合成试剂盒(Omega)进行反转录,试验操作按产品说明书进行。
1.6.3 实时荧光定量PCR扩增条件
反应体系用Real Super Mixture进行扩增,试验操作按产品说明书进行。扩增程序为:95℃5 min,(95 ℃ 15 s,60 ℃ 20 s,68 ℃ 20 s)×39个循环。反应体系为:REALSYBR Mixture(2×)10μL,10μmol/L上、下游引物各 0.4 μL,cDNA 2μL,加入灭菌蒸馏水至20μL。
1.6.4 标准曲线绘制
本试验以逆转录的cDNA作为待测基因的标准品制作标准曲线。具体方法如下:取逆转录的cDNA样本,进行10倍梯度稀释,每个稀释度设3组重复,进行实时荧光定量检测,采用2-△△Ct法分析待测肝脏中GPx4基因mRNA相对表达水平。
1.7 数据处理及统计分析
所有试验原始数据均用Excel 2003进行初步处理,应用SPSS 17.0软件进行统计分析,用oneway ANOVA进行单因素方差分析,采用Duncan氏法对差异显著的数据进行多重比较。试验数据均以平均值±标准误表示,P<0.05表示差异显著。
2 结果与分析
2.1 不同硒源及水平对蛋用种公鸡生长性能的影响
由表3可知,与对照组相比,各试验组的ADFI和F/G略有降低,其中以0.8 mg/kg硒化壳聚糖组最低,而ADG略有增加,但3个指标组间均差异不显著(P>0.05),但可以看出相同硒添加水平下,有机硒优于无机硒。
2.2 不同硒源及水平对蛋用种公鸡血浆及肝脏中硒含量的影响
由表4可知,在血浆中,1.2 mg/kg硒化壳聚糖组的硒含量显著高于对照组及其他试验组(0.8 mg/kg亚硒酸钠组除外)(P<0.05),分别依次高出了44.46%、44.15%、35.92%、37.75%和31.64%。在肝脏中,1.2 mg/kg亚硒酸钠组、0.8 mg/kg硒化壳聚糖组和1.2 mg/kg硒化壳聚糖组的硒含量与均对照组差异显著(P<0.05),分别依次提高了37.95%、36.83%和39.48%。亚硒酸钠和硒化壳聚糖均能增加血浆和肝脏的硒含量,并随着硒添加水平的升高而增加;并且总体上分析,有机硒的沉积率优于无机硒。

表3 不同硒源及水平对蛋用种公鸡生长性能的影响Table 3 Effects of different selenium sources and levels on growth performance of breeder roosters
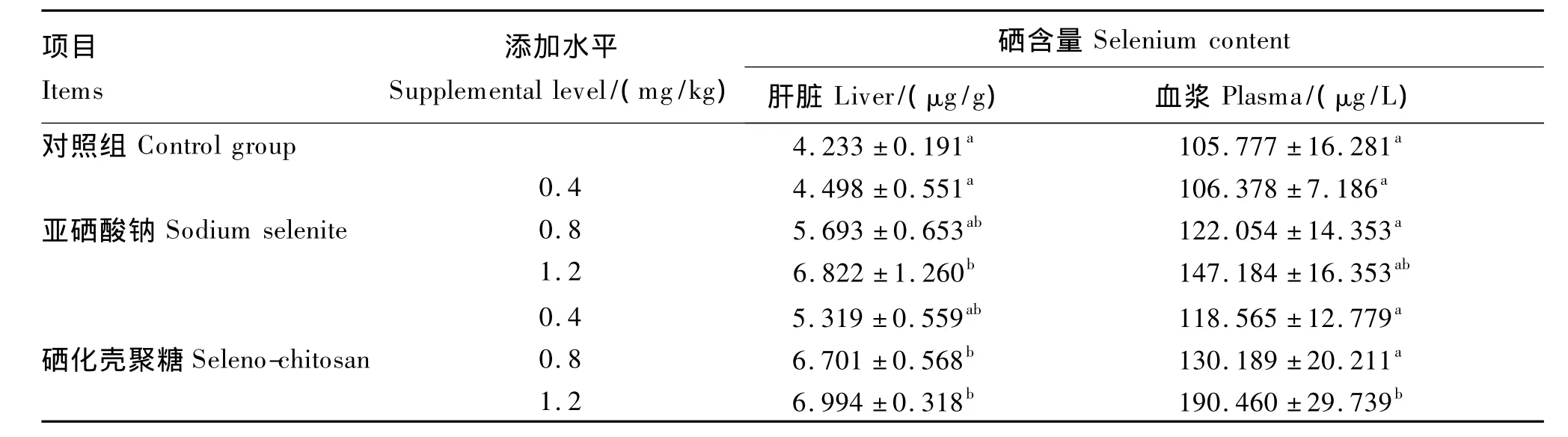
表4 不同硒源及水平对蛋用种公鸡血浆及肝脏中硒含量的影响Table 4 Effects of different selenium sources and levels on selenium content in plasma and liver of breeder roosters
2.3 不同硒源及水平对蛋用种公鸡肝脏抗氧化性的影响
由表5可知,0.4 mg/kg硒化壳聚糖组的GSH-Px活性显著高于对照组(P<0.05),比对照组提高了40.24%。0.8 mg/kg硒化壳聚糖组的T-SOD活性与对照组差异显著(P<0.05),比对照组提高了24.44%。其他各试验组的GSH-Px和T-SOD活性比对照组略有增加,但差异不显著(P>0.05)。
0.8 mg/kg亚硒酸钠组、1.2 mg/kg亚硒酸钠组、0.8 mg/kg硒化壳聚糖组和1.2 mg/kg硒化壳聚糖组的CAT活性均显著高于对照组(P<0.05),依次比对照组提高了62.37%、55.20%、68.43%和56.84%。各试验组的T-AOC较对照组略有增加,但差异不显著(P>0.05)。各试验组的MDA含量均显著低于对照组(P<0.05),按照硒添加水平由低到高,亚硒酸钠组依次比对照组降低了26.25%、60.49%和51.35%;硒化壳聚糖依次比对照组降低了45.95%、81.22%和55.54%。无论是亚硒酸钠组或硒化壳聚糖组,随着硒添加水平的增加,肝脏抗氧化性先升高后降低。
2.4 不同硒源及水平对蛋用种公鸡肝脏中硒酶活性的影响
由表6可知,TrxR和IDⅠ活性随硒添加水平增加,呈现出先升高后降低趋势。各试验组的TrxR活性均显著高于对照组(P<0.05),依次比对照组提高了 118.26%、38.58%、48.77%、40.65%、166.44%和118.26%。0.8 mg/kg硒化壳聚糖组的TrxR活性显著高于其他试验组(P<0.05),依次比其他试验组高出了 22.07%、36.49%、63.22%、58.13%和22.07%。

表5 不同硒源及水平对蛋用种公鸡肝脏抗氧化性的影响Table 5 Effects of different selenium sources and levels on antioxidant in liver of breeder roosters U/mg prot
0.8 mg/kg亚硒酸钠组IDⅠ活性与对照组及0.4 mg/kg亚硒酸钠组、0.4 mg/kg硒化壳聚糖组差异显著(P<0.05),依次比这3组提高了52.91%、37.88%和32.69%。0.8 mg/kg硒化壳聚糖组IDⅠ活性显著高于对照组和0.4 mg/kg亚硒酸钠组、0.4 mg/kg硒化壳聚糖组和1.2 mg/kg硒化壳聚糖组(P<0.05),依次提高了60.74%、48.21%、43.89%和35.23%。0.8 mg/kg硒化壳聚糖组与0.8 mg/kg亚硒酸钠组虽差异不显著(P>0.05),但比其提高了16.64%。

表6 不同硒源及水平对蛋用种公鸡肝脏中硒酶活性的影响Table 6 Effects of different selenium sources and levels on selenoenzyme activities in liver of breeder roosters
2.5 不同硒源及水平对蛋用种公鸡肝脏中基因表达水平的影响
2.5.1 RNA凝胶电泳
取5μL RNA用1%琼脂糖凝胶进行电泳,结果如图1,从图片中可清晰见目的条带,样本无异常降解,可用于后续试验。
2.5.2 不同硒源及水平对肝脏中GPx4基因mRNA相对表达水平的影响
由表7可知,0.8 mg/kg硒化壳聚糖组的GPx4基因mRNA相对表达水平与对照组差异显著(P<0.05),比对照组提高了56.79%。同时还可看出,随着硒添加水平的增加,GPx4基因mRNA相对表达水平先升高后降低,亚硒酸钠组在硒添加水平为0.8 mg/kg时就开始降低,而硒化壳聚糖组到1.2 mg/kg时才降低。2组硒源的GPx4基因mRNA相对表达水平虽然差异不显著(P>0.05),但硒化壳聚糖组比亚硒酸钠组提高了15.84%。
3 讨论
3.1 不同硒源及水平对蛋用种公鸡生长性能的影响
Wang等[10]研究发现,添加硒水平为0.2 mg/kg的有机硒和无机硒均显著降低了肉鸡的F/G。郭峰等[11]研究表明,在饲粮中添加酵母硒或无机硒可提高生长前期肉鸡的ADG,降低F/G,其中同水平的酵母硒增重量大于无机硒。田金可等[12]研究发现,添加有机硒显著降低了全期肉鸡ADFI,而各试验组对ADG及F/G均未产生显著影响。Wang等[13]研究表明,饲粮添加不同硒源对肉鸡的生长性能影响不显著。本试验研究表明,饲粮添加亚硒酸钠和硒化壳聚糖对蛋用种公鸡生长性能虽无显著影响,但与对照组相比,各试验组降低了ADFI和F/G,并且有机硒的效果优于无机硒,与Wang等[13]报道相一致,说明饲粮添加硒化壳聚糖可以更好的节约饲料,降低成本;而与 Wang 等[10]和郭峰等[11]的报道不一致,可能是由于所用试验动物的种属、试验条件以及生长阶段等不同导致的。添加较高剂量硒时,ADFI、F/G略有升高,ADG略有降低,可能是机体对硒的吸收达到了最大值,多余的硒随机体排出体外;同时并未出现厌食、生长停滞及其他急慢性中毒症状,所以初步推出硒添加水平为1.2 mg/kg时并未超过机体排出过多的硒的能力,但长期饲喂高剂量硒化壳聚糖是否会导致硒中毒,还有待于进一步研究其机理。
图1 RNA凝胶电泳结果Fig.1 Gel electrophoresis result of RNA

表7 不同硒源及水平对蛋用种公鸡肝脏中GPx4基因mRNA相对表达水平的影响Table 7 Effects of different selenium sources and levels on the GPx4 gene mRNA relative expression level in liver of breeder roosters
3.2 不同硒源及水平对蛋用种公鸡血浆及肝脏中硒含量的影响
硒的吸收主要在小肠中进行,无机硒主要以简单扩散方式通过肠壁的吸收进入肝脏,从而转化成生物硒被机体吸收,在高摄食情况下具有一定的毒性,并且促进氧化反应的发生;而有机硒具有独特的吸收特点,会通过主动运输穿过肠壁,或运输至肝脏与硒蛋白结合,或直接运输至机体组织与组织中的蛋白质结合[14]。Payne 等[15]比较有机硒与无机硒对鸡生物利用率的影响,结果显示,有机硒可显著增加组织与血浆中的硒含量。Yoon等[16]研究表明,与无机硒相比,有机硒可以显著提高动物组织中硒的存储量。Edens等[17]研究发现,随着饲粮中硒添加水平的提高,鸡体内组织硒含量随之增加,并且有机硒的硒沉积效果优于无机硒。Tiwary等[18]指出,羊饲粮中添加硒蛋氨酸或亚硒酸钠,硒蛋氨酸可显著提高其肝脏、肾皮质、心脏、全血及血清硒含量,并且以组织或器官内硒含量为依据,得出有机硒的利用优于无机硒。本试验结果表明,2种硒源均能显著增加血浆和肝脏的硒含量,均随硒添加水平增加而硒含量增加。这与上述的研究结果基本一致。饲粮中的硒添加水平直接影响机体中硒含量,饲粮硒含量在一定范围内时,动物机体的硒含量随饲粮硒添加水平的升高而呈线性提高;饲粮硒含量超出此范围时,组织、器官硒含量则会趋于平缓或降低,本试验中饲粮硒添加水平为1.2 mg/kg时,血浆及肝脏中硒含量并未趋于平缓或降低,则进一步说明硒添加水平未超出种公鸡机体对硒的耐受力。并且有机硒的沉积率高于无机硒,究其原因可能是因为家禽胃肠道对有机硒的吸收是主动的,所以吸收效率要优于无机硒。
3.3 不同硒源及水平对蛋用种公鸡肝脏抗氧化性的影响
动物机体的自由基以氧自由基为主,具有极强的氧化能力,可导致DNA的复制异常,同时引起生物膜中蛋白质、酶及磷酸交联失活以及细胞膜通透性的改变[19]。一般机体在正常情况下,本身的抗氧化系统基本上可以清除体内产生的自由基以及由自由基引起的过氧化物。但是在很多时候,很多外因,如病毒或者细菌的入侵、周围环境的改变等会造成体内自由基的大量聚集和增加,这会严重危及到机体的健康,长时间受其危害后会严重影响机体对自由基的清除能力。所以,随时清除体内的自由基,保持机体内环境的相对平衡是非常重要的。GSH-Px能特异地催化还原型谷胱甘肽(GSH)对过氧化氢的还原反应,可以起到保护细胞膜结构和功能完整的作用。SOD能特异性地清除机体代谢中产生的超氧自由基,以保护生物大分子不被氧化破坏,避免细胞损伤。CAT常常被细胞用做催化过氧化氢分解的工具,它的主要作用就是将过氧化氢分解为H2O与O2。T-AOC的值反映机体抗氧化酶系统以及机体自由基代谢状况。MDA含量可反映机体内脂质过氧化程度,并间接地反映出细胞损伤程度。这些酶促防御体系可以有效地清除氧自由基、过氧化氢等活性氧并且能够终止自由基的链式反应[20]。Wang等[10]研究表明,肉仔鸡饲粮中添加0.2~0.3 mg/kg的酵母硒可以显著增加血清和肝脏GSH-Px 活性。Dlouhci等[21]、Pappas 等[22]研究发现,在种鸡饲粮中适量添加不同硒源都可提高机体内抗氧化酶活性。本试验结果表明,饲粮添加硒化壳聚糖能显著提高蛋用种公鸡肝脏中GSH-Px、CAT、T-AOC和 T-SOD 活性,降低 MDA含量,本试验结果与以上研究报道及Tapiero等[23]、Spears等[24]的研究结果相一致,即补硒可改善家禽机体的抗氧化能力,并且有机硒的抗氧化能力强于无机硒。但在本试验中,当硒处于高水平范围内时,肝脏的抗氧化能力呈下降趋势,这与朱航等[25]、许宗运等[26]研究结果一致,分析其原因可能是硒虽然能清除氧自由基,但当组织内硒过量时,GSH过度氧化,损伤了膜结构,使细胞中超氧阴离子产量增加,降低了体内自由基水平,同时使某些机体抗氧化酶活性降低,从而导致家禽体内许多疾病和功能失调,此时硒的生物效应表现为毒性作用。本试验中,较高硒添加水平时,硒化壳聚糖组下降程度低于亚硒酸钠组,所以进一步说明硒化壳聚糖能更好地提高机体的抗氧化能力,增强机体对疾病的抵抗力。
3.4 不同硒源及水平对蛋用种公鸡肝脏中硒酶活性的影响
肝脏是甲状腺激素代谢的重要器官之一,其中的脱碘酶是IDⅠ,研究证实IDⅠ是一种含有硒半胱氨酸的蛋白酶,它在甲状腺激素的代谢中起非常重要的作用,并维持甲状腺激素的动态平衡[27]。IDⅠ主要是使甲状腺激素生理活性低的形式脱去其5'-碘转化为生理活性高的形式,其活性受硒添加水平的影响[28]。研究表明,缺硒可导致肝、肾等组织中IDⅠ的活性下降[29-30],而补硒可提高动物肝脏中IDⅠ的活性[31-32]。
TrxR是比较重要的硒蛋白之一,可以催化还原天然底物硫氧还蛋白。TrxR具有调节机体的氧化还原、调节细胞生长增殖等多种生物学活性。滕宗艳等[33]采用低硒饲料喂养大鼠,大鼠心肌中TrxR活性显著降低,说明心肌TrxR活性对硒营养较敏感;并进一步检测了心肌中TrxR的蛋白表达水平,低硒组蛋白水平较低,导致TrxR的还原功能减弱,使心肌抗氧化能力降低。
本试验结果表明,饲料添加亚硒酸钠和硒化壳聚糖均可显著提高种公鸡肝脏中IDⅠ和TrxR的活性,与上述报道及 Ganther等[34]研究结果相一致。其中硒添加水平为0.8 mg/kg时,IDⅠ和TrxR的活性达到最高值,随后硒添加水平增加到1.2 mg/kg时,IDⅠ和TrxR的活性开始下降,初步推断可能是饲料添加较高剂量的硒时,达到了种公鸡机体硒中毒水平,影响了组织器官的正常代谢功能,也影响到受到损害的组织器官的自我修复功能。从本试验结果还可看出,亚硒酸钠和硒化壳聚糖在硒添加水平相同时,硒化壳聚糖组的IDⅠ和TrxR的活性显著高于亚硒酸钠组,这也说明硒化壳聚糖的饲喂效果优于亚硒酸钠,可以更好地维持甲状腺激素代谢的动态平衡,调节机体的氧化还原等多种生物学活性。
3.5 不同硒源及水平对蛋用种公鸡肝脏中GPx4基因mRNA相对表达水平的影响
GPx4是机体内酶类抗氧化系统中的重要成员,不仅参与细胞内抗氧化反应,保护生物膜系统免受氧化损伤,而且还在细胞凋亡中起作用[35],是目前已知的唯一能直接降低细胞膜和脂蛋白内部磷脂氢过氧化物的抗氧化酶,它和维生素E一起发挥作用,抑制脂质过氧化反应[36]。赵庆等[37]研究指出,2种硒源(亚硒酸钠和酵母硒)不同添加水平均显著提高种公鸡睾丸中GPx4基因mRNA相对表达水平,且有机硒优于无机硒。Scimeca等[38]发现硒缺乏导致大鼠多种组织中GPx4活性下降。本试验结果显示,饲粮添加硒水平为0.8 mg/kg的硒化壳聚糖能显著提高种公鸡肝脏中GPx4基因mRNA相对表达水平,而亚硒酸钠则不显著,这与以上研究结果相一致。本试验中随着硒添加水平的增加,肝脏中GPx4基因mRNA相对表达水平先增高后降低,亚硒酸钠组在硒添加水平为0.8 mg/kg时就开始降低,而硒化壳聚糖组到1.2 mg/kg时才降低,由此说明种公鸡机体对有机硒的耐受力强于无机硒,饲粮添加硒化壳聚糖能更有效的保护细胞膜免受氧化损伤,保护线粒体的稳定状态,阻止细胞凋亡的发生。
综上所述,本试验条件下,饲粮添加硒化壳聚糖对22~28周龄的蛋用种公鸡生长性能影响不大,但硒化壳聚糖在低中硒添加水平(0.4和0.8 mg/kg)时,对肝脏中硒含量、抗氧化性、硒酶活性及GPx4基因mRNA相对表达水平有显著影响,亚硒酸钠则不显著;在较高硒添加水平(1.2 mg/kg)时肝脏中硒含量、抗氧化性、硒酶活性及GPx4基因mRNA相对表达水平有降低的趋势,其中硒化壳聚糖组降低的程度低于亚硒酸钠组,这表明蛋用种公鸡对硒化壳聚糖的耐受范围是宽于亚硒酸钠的,进一步说明有机硒优于无机硒;但饲喂期间未呈现出厌食、运动失调、生长状态不良等中毒症状,长期饲喂较高剂量的硒化壳聚糖是否会导致中毒,其有关原因还有待于进一步研究。
4 结论
饲粮添加硒化壳聚糖能显著增加蛋用种公鸡肝脏的硒含量,提高其抗氧化能力,增强其硒酶活性,以此来加强对疾病的抵抗力。确定硒的添加水平时,应该以硒的生物学功能得到最佳发挥为基础,因此在本试验条件下,饲粮添加低中硒水平(0.4和0.8 mg/kg)的硒化壳聚糖饲喂效果较好。
[1]魏长友.有机硒有助于母猪和断奶前仔猪的生长[J].国外畜牧学:猪与禽,2006,26(5):15-17.
[2]冯婧,王安,霍思远.饲粮添加硒对生长期蛋鸭生长性能、免疫机能及内分泌的影响[J].动物营养学报,2011,23(10):1697-1702.
[3]关丽萍,周良生,李术,等.硒对氟所致鸡睾丸组织p53 mRNA 表达的影响[J].中国家禽,2008,30(18):16-19.
[4]TASHJIAN D H,HUNG S S O.Selenium absorption,distribution,and excretion in white sturgeon orally dosed with L-selenomethionine[J].Environmental Toxicology and Chemistry,2006,25(10):2618-2622.
[5]VENDELAND S C,DEAGEN J T,BUTLER J A,et al.Uptake of selenite,selenomethionine and selenate by brush border membrane vesicles isolated from rat small intestine[J].Biometals,1994,7:305-312.
[6]WANG C L,LOVELL R T.Organic selenium sources,selenomethionine and selenoyeast,have higher bioavailability than an inorganic selenium source,sodium selenite,in diets for channel catfish(Ictalurus punctatus)[J].Aquaculture,1997,152:223-234.
[7]侯少范,李德珠,王丽珍,等.我国土壤中结合态硒的含量和分布规律[J].地理研究,1990,9:17-25.
[8]易秀.生态环境中的硒及其地方病[J].西安工程学院学报,2000(4):69-72.
[9]OLDFIELD J E.Selenium world atlas:selenium-tellurium development association[M].Grimbergen:[s.n.],1999.
[10]WANG Y B,XU B H.Effect of different selenium source(sodium selenite and selenium yeast)on broiler chickens[J].Animal Feed Science and Technology,2008,144:306-314.
[11]郭峰,李同树,苗朝华,等.日粮硒源及添加水平对肉鸡生产性能、肉质和甲状腺激素的影响[J].动物营养学报,2004,16(3):63-64.
[12]田金可,HUSSAIN A,王恬,等.不同硒源及水平对肉鸡组织硒含量及抗氧化功能的影响[J].动物营养学报,2012,24(6):1030-1037.
[13]WANG Y X,ZHAN X A,AHANG X W,et al.Comparison of different forms of dietary selenium supplementation on growth performance,meat quality,selenium deposition,and antioxidant property in broilers[J].Biological Trace Element Research,2011,143(1):261-273.
[14]COMBS G F,COMBS S B.Absorption and transfer[M]//The role of selenium in nutrition.New York:Academic Press,1986.
[15]PAYNE R L,SOUTHERN L L.Comparison of inorganic and organic selenium sources for broilers[J].Poultry Science,2005,84:898-902.
[16]YOON I,WERNER T M,BUTLER J M.Effect of source and concentration of selenium on growth performance and selenium retention in broiler chickens[J].Poultry Science,2007,86:727-730.
[17]EDENS F W,PARKHURST C R,HAVENSTEIN G B,et al.Housing and selenium influences on feathering in broilers[J].Journal Applied Poultry Research,2001,10(2):128-134.
[18]TIWARY A K,STEGELMEIER B L,PANTER K E,et al.Comparative toxicosis of sodium selenite and selenomethionine in lambs[J].Journal of Veterinary Diagnostic Investigation,2006,18(1):61-70.
[19]郭小权,胡国良,曹华斌,等.不同形式和水平的锌对断奶仔猪抗氧化功能的影响[J].黑龙江畜牧兽医,2010(5):64-65.
[20]凌波.微量元素硒与自由基[J].微量元素与健康研究,2007,24(3):67-68.
[21]DLOUHA G,SEVEIKOVD S,DOKOUPILOVCI A,et a1.Effect of dietary selenium sources on growth performance,breast muscle selenium,glutathione peroxidase activity and oxidative stability in broilers[J].Czech Journal of Animal Science,2008,53:265-269.
[22]PAPPAS A C,KARADAS F,SURAL P F,et al.The selenium intake of the female chicken influences the selenium status of her progeny[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2005,142:465-474.
[23]TAPIERO H,TOWNSEND D M,TEW K D.The antioxidant role of selenium and seleno-compounds[J].Biomedicine & Pharmacotherapy,2003,57(3/4):133-134.
[24]SPEARS J W,GRIMES J,LLOYD K,etal.Efficacy of a novel organic selenium compound(zinc-1-selenomethionine,Availa Se)in broiler chicks[R]//Proceedings of the 1stlatin American congress of animal nutrition.Cancun:[s.n.],2003:197-198.
[25]朱航,何秋实,吕阳,等.富硒酵母对铁过量导致小鼠肝损伤保护作用的实验研究[J].热带医学研究,2007,7(8):732-734.
[26]许宗运,张丽娟,韩俊文.不同水平酵母硒对奶牛血液抗氧化能力的影响[J].动物营养学报,2007,19(6):753-757.
[27]BERRY M J,BANU L,LARSEN P R.Type Ⅰ iodothyronine deiodinase is a selenocysteine-containing enzyme[J].Nature,1991,349:438-440.
[28]ARTHUR J R,BECKETT G J.Roles of selenium in type Ⅰ iodothyronine 5'-deiodinase and in thyroid hormone and iodine metabolism[M]//Selenium in biology and human health.New York:Springer-Verlag,1994:93-115.
[29]BECKETT G J,RUSSELL A,NICOL F,et al.Effect of selenium deficiency on hepatic typeⅠ5-iodothyronine deiodinase activity and hepatic thyroid hormone levels in the rat[J].The Biochemical Journal,1992,282:483-486.
[30]BATES J M,SPATE V L,MORRIS J S,et al.Effects of selenium deficiency on tissue selenium content,deiodinase activity,and thyroid hormone economy in the rat during development[J].Endocrinology,2000,141(7):2490-2500.
[31]HE JH,OHTSUKA A,HAYASHI K.Selenium influences growth via thyroid hormone status in broiler chickens[J].British Journal of Nutrition,2000,84(5):727-732.
[32]WICHTEL J J,CRAIGIE A L,FREEMAN D A,et al.Effect of selenium and iodine supplementation on growth rate and on thyroid and somatotropic function in dairy calves at pasture[J].Journal of Dairy Science,1996,79(10):1865-1872.
[33]滕宗艳,马兰,周萍,等.低硒心肌损伤与硫氧还蛋白还原酶的关系研究[J].哈尔滨医科大学学报,2003,37:305-307.
[34]GANTHER H E,IP C.Thioredoxin reductase activity in rat liver is not affected by supranutritional levels of monomethylated selenium in vivo and is inhibited only by high levels of selenium in vitro[J].The Journal of Nutrition,2001,13l(2):301-304.
[35]LIANG H Y,VAN REMMEN H,FROHLICH V,et al.Gpx4 protects mitochondrial ATP generation against oxidative damage[J].Biochemical and Biophysical Research Communications,2007,356(4):893-898.
[36]YANT L J,RAN Q T,RAO L,et al.The selenoprotein GPx4 is essential for mouse development and protects from radiation and oxidative damage insults[J].Free Radical Biology and Medicine,2003,34(4):496-502.
[37]赵庆,刘爱巧,王晓霞,等.饲粮硒对种公鸡睾丸中硒含量和硒蛋白酶基因mRNA表达的影响[J].动物营养学报,2011,23(6):1011-1018.
[38]SCIMECA M S,LISK D J,PROLLA T,et al.Effects of GPX4 haploid insufficiency on GPx4 activity,selenium concentration,and paraquat-induced protein oxidation in murine tissues[J].Experimental Biology Medicine,2005,230(10):709-714.
猜你喜欢
动物营养学报(2022年8期)2022-09-01 03:20:52
快乐语文(2021年36期)2022-01-18 05:48:40
动物营养学报(2017年8期)2017-08-16 09:40:00
小天使·一年级语数英综合(2016年5期)2016-05-14 12:21:05
食品界(2016年4期)2016-02-27 07:36:46
动物营养学报(2015年9期)2016-01-07 11:29:44
小天使·一年级语数英综合(2015年10期)2015-10-14 06:23:24
小青蛙报(2014年13期)2014-09-22 21:07:30
应用化工(2014年7期)2014-08-09 09:20:21
应用技术学报(2014年4期)2014-02-28 14:52:40
