
不同蛋白质水平饲料中添加α-酮戊二酸对松浦镜鲤生长性能、体成分和血清生化指标的影响
2013-09-20 03:05:50位莹莹徐奇友李晋南王常安赵志刚
动物营养学报 2013年12期
位莹莹 徐奇友 李晋南 王常安 罗 亮 赵志刚
(1.上海海洋大学水产与生命学院,上海 201306;2.中国水产科学研究院黑龙江水产研究所,动物营养实验室,哈尔滨 150070)
谷氨酰胺是动物体内游离氨基酸池中最为丰富的一种氨基酸,占细胞内游离氨基酸的60%以上[1]。近年来,谷氨酰胺对机体肠道营养与免疫等方面的作用受到了国内外众多学者的广泛关注,但是谷氨酰胺单体在溶液中不稳定,对温度和酸碱度敏感,溶解度低,易被环化为有毒性的焦谷氨酸和氨,上述缺点限制了其在动物生产中的应用。而α-酮戊二酸(α-ketoglutarate,AKG)作为谷氨酰胺的前体物质,在溶液中具有良好的稳定性[2]。AKG在谷氨酸脱氢酶或转氨酶的作用下生成谷氨酸,并进一步通过谷氨酰胺合成酶形成谷氨酰胺发挥谷氨酰胺的作用,这种转变在动物体内是迅速的,从而可以替代谷氨酰胺应用于生产实践[3]。目前关于 AKG 对仔猪[4-5]、断奶大鼠[6]等生长、肠道发育、免疫等方面的影响已有一些文献报道,而对水产动物的研究仍是一片空白。松浦镜鲤是黑龙江水产研究所在德国镜鲤选育系(F4)的基础上选育的新品种,具有少鳞、生长速度快、体形好、养殖成活率高等优点。本试验拟以松浦镜鲤为研究对象,研究在不同蛋白质水平饲料中添加AKG对松浦镜鲤生长性能、体成分和血清生化指标的影响,以期为AKG替代谷氨酰胺应用于水产饲料提供理论依据。
1 材料与方法
1.1 试验材料
AKG:购自Sigma-Aldrich公司,白色粉末,纯度≥98.5%。
1.2 试验饲料
根据鲤鱼营养需求配制基础饲料,以蒸汽鱼粉为动物蛋白质源,豆粕为植物蛋白质源,混合油(磷脂∶鱼油∶豆油 =1.0∶2.5∶1.5)为脂肪源,以次粉和豆粕的添加比例调节饲料蛋白质水平。试验设4个蛋白质水平(28%、31%、34%和37%),每个蛋白质水平下设3个 AKG浓度[0(对照)、0.75%和1.50%],共配制12种试验饲料,其组成及营养水平见表1。各饲料原料粉碎过80目筛,逐级混合均匀,再加入一定量的水分充分混合后用小型颗粒机挤压成直径为2 mm的颗粒饲料,常温风干后,-20℃保存备用。

表1 试验饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of experimental diets(DM basis) %
1.3 试验分组及饲养管理
试验用鱼购自黑龙江水产研究所呼兰试验站,养殖于黑龙江水产研究所养殖车间的可控温水循环系统中,水温24℃左右,溶氧大于5 mg/L,氨氮小于0.1 mg/L,自然光照。试验前将松浦镜鲤经3%盐水消毒后在暂养池驯养2周,挑选体健无伤、规格整齐、平均体重为(61.28±3.40)g的松浦镜鲤720尾,随机分成12组,每组3个重复,每个重复20尾鱼。12组试验鱼分别投喂不同的试验饲料,每天饱食投喂,日投喂3次,分别为每天的08:00、13:00、18:00,同时记录投喂量,投喂30 min后,收集残饵,烘干后称重,定期换水、消毒,确保水质良好。养殖周期为8周。
1.4 样品采集及指标测定
1.4.1 样品采集与处理
养殖试验结束后各组试验鱼饥饿24 h后采样。统计每缸鱼的总数、称总重后,随机取3尾鱼-40℃冷冻保存,用于体成分分析;另随机取3尾鱼用浓度为100 mg/L的MS-222(化学名称:间氨基苯甲酸乙酯甲磺酸盐)麻醉,称重、量体长后尾静脉取血,全血置于一次性试管中于4℃冰箱中静置1~2 h,3 500 r/min离心30 min,取上层血清分装于1.5 mL离心管中,暂存于-20℃冰箱用于血清生化指标的测定;取内脏团和肝胰脏称重。计算出各组试验鱼的增重率、蛋白质效率、肝体指数、脏体指数、肥满度、饲料系数,各指标计算公式如下:

1.4.2 指标测定
水分含量的测定采用105℃烘箱干燥恒重法(GB/T 5009.3—2003);粗蛋白质含量的测定采用凯氏定氮法(GB/T 5009.5—2003);粗脂肪含量的测定采用索氏抽提法(GB/T 5009.6—2003);粗灰分含量的测定采用550℃高温灼烧法(GB/T 5009.4—2003)。
将制备好的血清样品送至黑龙江省电力医院,采用贝克曼ProCX4全自动生化分析仪进行测定。各指标测定方法如下:总蛋白(TP)含量的测定采用双缩脲法;白蛋白(ALB)含量的测定采用溴甲酚绿法;球蛋白(GLB)含量经计算得出,为TP与ALB含量的差值;谷丙转氨酶(ALT)和谷草转氨酶(AST)活性的测定采用速率法;甘油三酯(TG)和总胆固醇(TCHO)含量的测定采用酶法;碱性磷酸酶(ALP)活性的测定采用比色法;肌酐(CREA)含量的测定采用酶法;尿酸(UA)含量的测定采用磷钨酸还原法。
1.5 数据处理与分析
试验数据采用SPSS 17.0软件分析,以蛋白质水平和AKG浓度为影响因素,采用双因素方差分析和Duncan氏法多重比较进行统计,显著水平设定为P<0.05。试验结果以平均值±标准差表示。
2 结果与分析
2.1 不同蛋白质水平饲料中添加AKG对松浦镜鲤生长性能的影响
由表2可以看出,各个蛋白质水平下,饲料中添加AKG均有提高松浦镜鲤增重率、蛋白质效率和降低饲料系数的趋势,除34%蛋白质水平下松浦镜鲤增重率显著提高(P<0.05)外,其余差异不显著(P>0.05)。28%、31%、34%和37%蛋白质水平下,0.75%和1.50%AKG组增重率分别比对照组(未添加AKG组)提高了10.71%和29.95%、5.49%和17.37%、42.04%和23.71%、2.41%和-9.27%,饲料系数降低了-9.58%和5.39%、6.67%和 9.63%、6.29%和 5.59%、1.60%和4.80%。各个蛋白质水平下,饲料中添加AKG对松浦镜鲤肝体指数、脏体指数和肥满度均无显著影响(P>0.05)。就松浦镜鲤的生长性能而言,蛋白质水平和AKG浓度之间不存在显著的交互作用(P>0.05)。

表2 不同蛋白质水平饲料中添加AKG对松浦镜鲤生长性能的影响Table 2 Effects of AKG supplementation in different protein level diets on growth performance of Songpu mirror carp
2.2 不同蛋白质水平饲料中添加AKG对松浦镜鲤体成分的影响
由表3可以看出,各个蛋白质水平下,饲料中添加AKG对松浦镜鲤全鱼体成分均无显著影响(P>0.05)。蛋白质水平和AKG浓度的交互作用除对全鱼粗灰分含量有显著影响(P<0.05)外,对其他体成分指标均无显著影响(P>0.05)。
2.3 不同蛋白质水平饲料中添加AKG对松浦镜鲤血清生化指标的影响
由表4可以看出,AKG显著影响血清ALT、AST活性及UA、TG含量(P<0.05)。蛋白质水平和AKG浓度的交互作用除对血清UA和TG含量有显著影响(P<0.05)外,对其他各指标均无显著影响(P>0.05)。28%蛋白质水平下,1.50%AKG组血清TG含量显著低于对照组(P<0.05),与0.75%AKG组差异不显著(P>0.05)。31%蛋白质水平下,1.50%AKG组血清ALB含量和ALT活性显著高于其他组(P<0.05);0.75%和1.50%AKG组血清UA和TG含量显著低于对照组(P<0.05)。34%蛋白质水平下,0.75%AKG组血清TP和TG含量显著高于对照组(P<0.05)。37%蛋白质水平下,1.50%AKG组血清ALT活性和UA含量显著高于其他组(P<0.05);0.75%和1.50%AKG组血清AST活性显著高于对照组(P<0.05)。各个蛋白质水平下,饲料中添加AKG对血清GLB、TCHO、CREA含量及ALP活性均无显著影响(P>0.05)。
3 讨论
3.1 不同蛋白质水平饲料中添加AKG对松浦镜鲤生长性能的影响
目前,AKG对畜禽生产性能影响的报道较多。胡泉州等[7]研究表明,饲料添加1%AKG可以促进仔猪生长并减弱免疫应激对仔猪生长性能的影响。余亲平等[8]研究得出,在肉仔鸡饲粮中添加0.7%AKG可显著提高2周龄的体重及1~2周龄的平均日增重,其促生长效果优于0.7%谷氨酰胺。王金泉等[9]研究发现,中草药和AKG联合使用比单独使用对断奶仔猪的促生长效果更明显,饲料回报率更高。此外,研究还表明,饲粮中添加能显著缓解脂多糖应激对断奶仔猪生长的抑制作用[10]。

表3 不同蛋白质水平饲料中添加AKG对松浦镜鲤体成分的影响Table 3 Effects of AKG supplementation in different protein level diets on body composition of Songpu mirror carp%
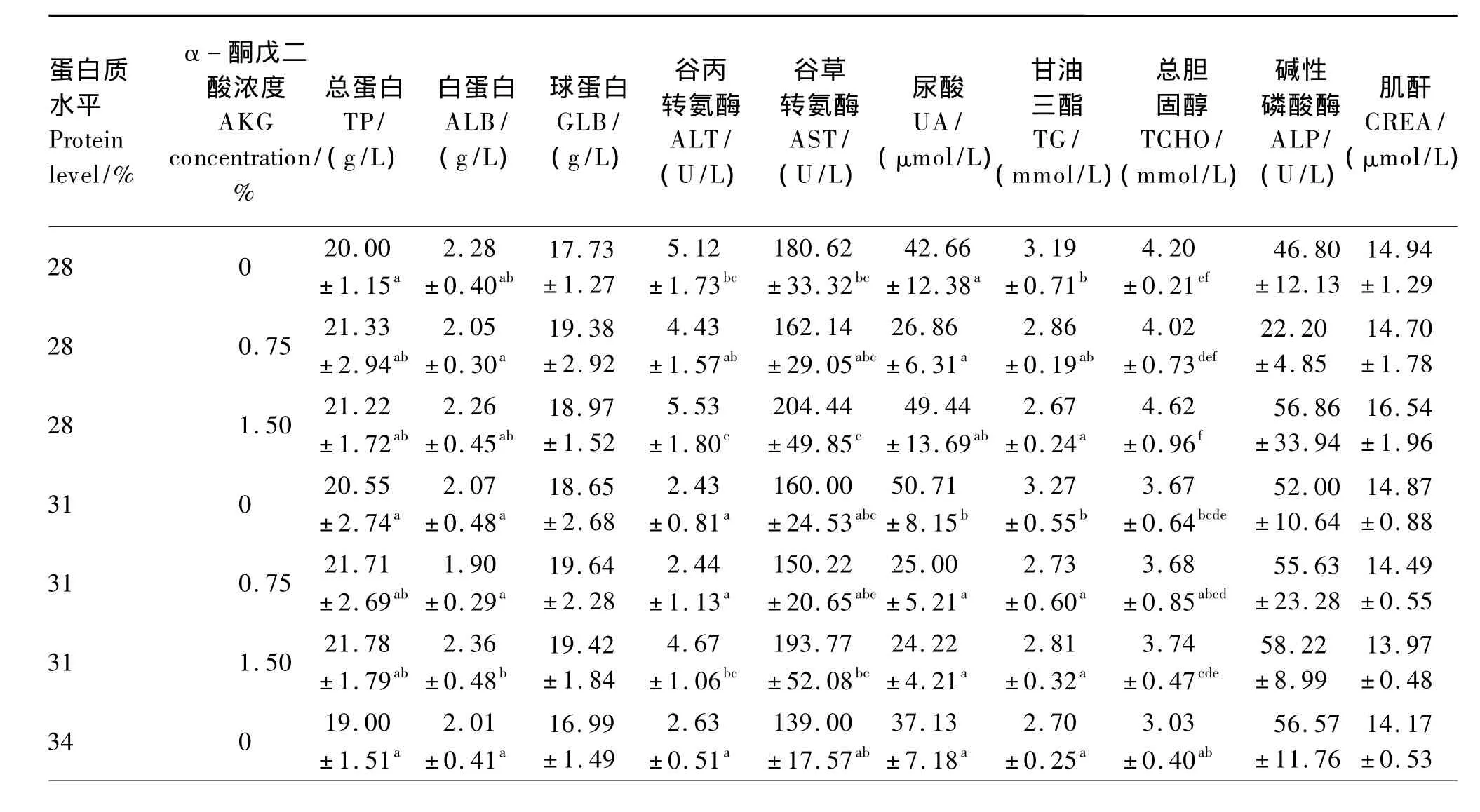
表4 不同蛋白质水平饲料中添加AKG对松浦镜鲤血清生化指标的影响Table 4 Effects of AKG supplementation in different protein level diets on serum biochemical indices of Songpu mirror carp

续表4
目前未见AKG在水产动物生产中应用的报道。本试验条件下,不同蛋白质水平下,饲料中添加AKG对松浦镜鲤生长性能的影响不尽相同,可能与谷氨酰胺为条件性必需氨基酸有关。28%、31%和34%蛋白质水平下,添加AKG的各组松浦镜鲤的增重率与对照组相比均有所提高,其中,34%蛋白质水平下0.75%AKG组松浦镜鲤增重率显著高于对照组;37%蛋白质水平下,1.50%AKG组松浦镜鲤增重率反而低于对照组。这可能是因为AKG直接用于氧化供能或直接排出体外而没有用于体增长。目前,未见关于AKG对水生动物营养利用率的报道,但在陆生动物上有相关报道。王金泉等[9]研究表明,AKG可以促进饲粮中营养物质的吸收,对促进断奶仔猪的生长具有协同作用。本试验条件下,在饲料能量和蛋白质水平基本相同的情况下,随AKG浓度的增加饲料蛋白质效率有增加趋势,饲料系数有降低趋势,说明饲料中添加AKG对饲料利用有积极作用。
3.2 不同蛋白质水平饲料中添加AKG对松浦镜鲤体成分的影响
Yao等[11]研究表明AKG可以促进蛋白质的合成,而本试验条件下,鱼体的粗蛋白质含量相对稳定,受饲料AKG浓度影响较小,其原因可能为Yao等[11]的研究是以体外培养的猪肠道上皮细胞为研究对象,结果显示AKG可以促进细胞蛋白质合成,而本试验是以鱼体为研究对象,AKG促进细胞合成的蛋白质并没有用于鱼体蛋白质的沉积。其具体代谢机制有待于进一步研究。
3.3 不同蛋白质水平饲料中添加AKG对松浦镜鲤血清生化指标的影响
血清TP包括ALB和GLB,具有运输多种代谢产物,维持血管内正常胶体渗透压和酸碱度等多种生理功能。本试验条件下,31%蛋白质水平下,饲料添加1.50%AKG可以显著提高血清中ALB含量;34%蛋白质水平下,饲料添加0.75%AKG可以显著提高血清中TP和GLB含量。这表明,一定蛋白质水平下,饲料添加AKG可以促进松浦镜鲤蛋白质的合成。这与黄冠庆等[12]在鸡上的研究结果一致。血清中非蛋白质类含氮物主要包括尿素、UA等,其含量可以反映体内蛋白质分解代谢和肾功能情况,当饲料氨基酸平衡时,血清尿素氮及UA含量下降[13]。本试验条件下,饲料添加AKG后松浦镜鲤血清中UA含量均有不同程度的降低,表明饲料添加AKG有利于氨基酸平衡,有利于蛋白质代谢。ALT和AST是动物体内最主要的2个转氨酶,在氨基酸代谢方面发挥重要作用,是反映蛋白质代谢利用率的重要指标[14]。当饲料中蛋白质水平升高时,氨基酸代谢强度增加,从而造成转氨酶活性升高[15]。本试验中,饲料中添加AKG可以显著提高血清中ALT和AST活性,表明饲料中添加AKG有利于促进松浦镜鲤肝胰脏氨基酸代谢,提高蛋白质利用率,促进蛋白质合成。以上结果表明,饲料中添加一定量的AKG可以促进松浦镜鲤蛋白质代谢。
TG和TCHO的含量反映了动物体内脂肪沉积的情况[16]。低蛋白质水平(28%、31%)下饲料添加AKG可以显著降低血清中TG含量,而高蛋白质水平(34%、37%)下饲料添加AKG虽有升高血清中TG含量的趋势,但差异不显著,以上结果表明在低蛋白质水平饲料中添加AKG有利于降低松浦镜鲤血脂,利于鱼体健康。CREA是肌肉组织中储能物质肌酸代谢的终产物,血清CREA的含量能反映肾脏的排泄功能。血清中的ALP主要来自骨骼,由成骨细胞产生。ALP经肝胆系统进行排泄。所以当ALP产生过多或排泄受阻时,均可使血中ALP活性发生变化。本试验条件下,饲料中添加AKG未对血清中CREA含量、ALP活性产生显著影响,表明饲料中添加AKG未对松浦镜鲤产生负面影响。
4 结论
饲料中添加适量的AKG可以在一定程度上提高松浦镜鲤的增重率和蛋白质效率,降低饲料系数,促进鱼体蛋白质和脂肪代谢。
[1]谭文军,吴金生.谷氨酰胺的代谢及在肠外营养液中应用的进展[J].肠外与肠内营养,2001,8(1):41-43.
[2]WU G,MEIER S A,KNABE D A.Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs[J].The Journal of Nutrition,1996,126(10):2578-2584.
[3]KRISTENSEN N B,JUNGVID H,FERNANDEZ J A,et al.Absorption and metabolism ofα-ketoglutarate in growing pigs[J].Journal of Animal Physiology and Animal Nutrition,2002,86(7/8):239-245.
[4]HOU Y Q,WANG L,DING B Y,et al.Dietaryα-ketoglutarate supplementation ameliorates intestinal injury in lipopolysaccharide-challenged piglets[J].Amino Acids,2010,39(2):555-564.
[5]BUDDINGTON R K,PAJOR A,BUDDINGTON K K,et al.Absorption ofα-ketoglutarate by the gastrointestinal tract of pigs[J].Comparative Biochemistry and Physiology,Part A:Molecular&Integrative Physiology,2004,138(2):215-220.
[6]SEGAUD F,COMBARET L,NEVEUX N,et al.Effects of ornithineα-ketoglutarate on protein metabolism in Yoshida sarcoma-bearing rats[J].Clinical Nutrition,2007,26(5):624-630.
[7]胡泉州,侯永清,丁斌鹰,等.α-酮戊二酸对仔猪小肠组织学形态与功能的影响[J].动物营养学报,2008,20(6):662-667.
[8]余亲平,陈雁群,谢金蝉,等.日粮添加α-酮戊二酸对肉仔鸡生长性能及组织器官发育的影响[J].中国畜牧兽医,2010,37(10):10-14.
[9]王金泉,项方献,姚刚.中草药结合AKG对断奶仔猪生长及消化吸收功能的影响[J].西北农林科技大学学报:自然科学版,2011,39(5):27-31.
[10]刘坚,侯永清,丁斌鹰,等.α-酮戊二酸对脂多糖应激断奶仔猪生长抑制的缓解作用[J].动物营养学报,2009,21(4):519-524.
[11]YAO K,YIN Y L,LI X L,et al.Alpha-ketoglutarate inhibits glutamine degradation and enhances protein synthesis in intestinal porcine epithelial cells[J].Amino Acids,2012,42(6):2491-2500.
[12]黄冠庆,余亲平,陈雁群,等.α-酮戊二酸对黄羽肉鸡饲料代谢能和蛋白质代谢的影响[J].中国饲料,2012(18):22-24.
[13]李忠荣,陈婉如,叶鼎承,等.低蛋白质补充氨基酸饲粮对北京鸭生长性能、血清生化指标及粪氮含量的影响[J].动物营养学报,2013,25(2):319-325.
[14]唐玲,徐奇友,王常安,等.不同水温和饲料蛋白质水平对镜鲤血清生化指标的影响[J].大连海洋大学学报,2011,26(1):41-46.
[15]ABDEL-TAWWAB M,AHMAD M H,KHATTAB Y A,et al.Effect of dietary protein level,initial body weight,and their interaction on the growth,feed utilization,and physiological alterations of Nile tilapia,Oreochromis niloticus(L.)[J].Aquaculture,2010,298(3/4):267-274.
[16]姜柯君,王际英,张利民,等.饲料中添加小肽对星斑川鲽幼鱼生长性能、体组成及血清生化指标的影响[J].动物营养学报,2013,25(1):222-230.
猜你喜欢
World Journal of Pediatrics(2023年11期)2023-12-11 01:35:46
肝博士(2022年3期)2022-06-30 02:48:48
思维与智慧·下半月(2022年5期)2022-05-17 00:54:54
海外星云(2021年9期)2021-10-14 07:26:10
青苹果·教育研究版(2016年9期)2016-12-23 11:52:36
中外医疗(2015年11期)2016-01-04 03:58:50
医学研究杂志(2015年12期)2015-06-10 06:57:46
食品工业科技(2014年9期)2014-03-11 18:15:27
城市道桥与防洪(2014年4期)2014-02-27 07:25:53
城市道桥与防洪(2013年8期)2013-03-11 15:18:06
