
中国东北部野生杏资源ISSR遗传多样性1)
2013-09-18 11:11马丽娟郭太君何丹丹
东北林业大学学报 2013年12期
马丽娟 郭太君 何丹丹
(吉林农业大学,长春,130118)
李 锋 张艳波
(吉林省农业科学院果树研究所)
杏是世界上重要的果树和观赏树种之一,在各大洲都有栽培。中国是杏的遗传多样性分布中心,具有丰富的种质资源。我国东北部寒冷地区野生杏属植物资源位于世界杏属植物生态群的东北亚生态群[1],典型的特征是果实小、核仁小、味苦,果肉可食性差;树体抗寒力强,可耐-30℃以下的低温,通常作为园林绿化树种和栽培杏的砧木加以利用,也有少数用作杏抗寒育种的亲本。东北野生杏主要是西伯利亚杏(Armeniaca sibirica(L.)Lam.)、辽 杏 (A.mandshurica(Maxim.)Skv.)或是带有西伯利亚杏、辽杏、普通杏(A.vulgaris Lam.)血缘的杂合体。由于21世纪八十、九十年代人们对于土地的不当利用、林地的开发和生态环境的恶化,使野生杏资源的数量急剧减少,物种的多样性受到严重的威胁。本文利用ISSR分子标记技术对东北野生杏遗传多样性分析,以期获得东北野生杏居群遗传变异情况,为居群划分、优良品种培育及野生资源保护提供理论基础。
1 材料与方法
材料来自黑龙江的林口(LK)、东宁(DN)、兴凯湖(XKH)和吉林通榆(TY)4个野生杏居群共176份样本。其中通榆居群位于亚洲最大的野生西伯利亚杏林,林口居群、东宁居群可能是野生西伯利亚杏,兴凯湖居群可能是野生辽杏。每个居群根据杏树分布范围选取样株,于野外取幼嫩叶片低温带回实验室,液氮冷冻后-80℃保存备用。为保证重复取样,对每株样树的形态(树高、胸径/株型、生境)进行记录和拍照并做好标记。采样点地图见图1,每个居群的产地、生境及采样数见表1。
1.1 ISSR-PCR 体系及扩增
DNA提取和检测按文献[2]进行。试验所用ISSR引物序列,由上海生工生物工程技术服务有限公司合成,从40条ISSR引物中筛选出12条扩增条带清晰、多态性高、稳定性好的引物用于全部样本的PCR扩增。ISSR-PCR反应体系经比较和优化确定为20 μL,其中模板 DNA 2 μL(20 ng/μL),10×buffer 2 μL,超纯 dNTP 1.6 μL(2.5 mmol/L),引物 1 μL(10 μm/L),Taq DNA 聚合酶 0.4 μL(2.5 U/μL),双蒸水 13 μL。扩增程序参考[1]:94 ℃ 预变性 5 min;94℃变性45 s,51~59℃退火 1 min,72℃延伸1.5 min,共40个循环;72℃后延伸7 min;扩增后4℃保存。扩增产物(6 μL PCR产物)用含EB的1.5%琼脂糖凝胶电泳检测,在紫外凝胶成像仪下观察拍照。

表1 4个野生杏居群基本生境条件

图1 野生杏居群采样点示意图
1.2 数据统计与分析
由于ISSR是显性标记,在相同电泳迁移位置的条带具有同源性。用DL2000Maker作为分子量标准,对获得的电泳条带进行0、1编码,有带(包括弱带)记为“1”,无带记为“0”,建立0、1原始矩阵。采用 POPGENE1.32[3]软件对各居群和全部个体进行遗传参数分析。分别计算了多态性条带百分率(BPP)、有效等位基因数(Ne)、Nei’s基因多样度(H)、Shannon’s信息指数(Ho),总的遗传多样性(Ht)、居群内遗传多样性(Hs)、基因分化系数(Gst)(Gst=1-Hs/Ht)和基因流(Nm,gene flow),Nei’s遗传距离(D)与遗传一致度(I)。利用 NTSYS-pc2.10[4]软件,依据样品的Nei’s遗传距离和Nei&Li相似系数,进行UPGMA(非加权配对算数平均法)聚类分析。
2 结果与分析
2.1 引物扩增结果
从40条ISSR引物中筛选出的12条效果好的ISSR引物,对4居群176个个体进行PCR扩增。部分引物扩增结果见图2。12条引物产生的清晰条带在9~18条,共扩增出180条带,多态性条带149条,占82.8%,平均每个引物的扩增条带为15条;不同引物产生的多态性百分比在73.3% ~100%(表2),表明12条ISSR引物均具有较高的多态性。其中引物UBC855扩增多态性最高为100%,引物UBC881扩增多态性最低为73.3%。

表2 对野生杏4个居群176个样本进行ISSR扩增的引物序列扩增结果
2.2 遗传多样性
评价群体遗传多样性水平或等位基因丰富程度的指标主要有多态性百分率、Nei’s基因多样度、Shannon信息指数、有效等位基因数。本研究结果如表3所示:4个居群中多态性百分率、Nei’s基因多样度、Shannon信息指数变化均以兴凯湖居群(XKH)最高,林口居群(LK)最低,通榆居群(TY)、东宁(DN)居中;表明XKH居群野生辽杏的遗传多样性高于野生西伯利亚杏。野生杏个体水平的多态性百分率、有效等位基因数、Nei’s基因多样度和Shannon 信息指数分别为 82.8%、1.4096、0.2471、0.3799,均高于居群范围52.8% ~73.9%、1.2947 ~1.3836、0.1728 ~0.2269、0.2608 ~0.3456。

图2 引物UBC843对东宁居群(DN)25-41号样本扩增结果
野生杏个体总的遗传多样性为0.2468,而居群内的遗传多样性为0.2043,4个居群间的基因分化系数为0.1720。居群间遗传变异和居群内的遗传变异分别占总遗传变异的17.2%和82.8%,表明遗传变异主要存在于居群内部。居群间基因流为 2.4063。

表3 4个野生杏居群的遗传多样性指数
2.3 遗传关系聚类
为了分析不同居群间亲缘关系远近,计算了不同居群间的 Nei’s遗传一致度(I)和遗传距离(D)[5]列于表4。可以看出I值变化范围是0.9041~0.9711,均大于0.9,说明4个居群的亲缘关系很接近。其中TY和DN的遗传一致度最大,说明这两个居群的亲缘关系最近,TY和LK的遗传一致度最小,说明TY和LK亲缘关系最远。

表4 4个野生杏居群的Nei’s遗传一致度和遗传距离的关系
根据Nei’s遗传距离利用UPGMA法构建的居群遗传关系聚类图见图3。
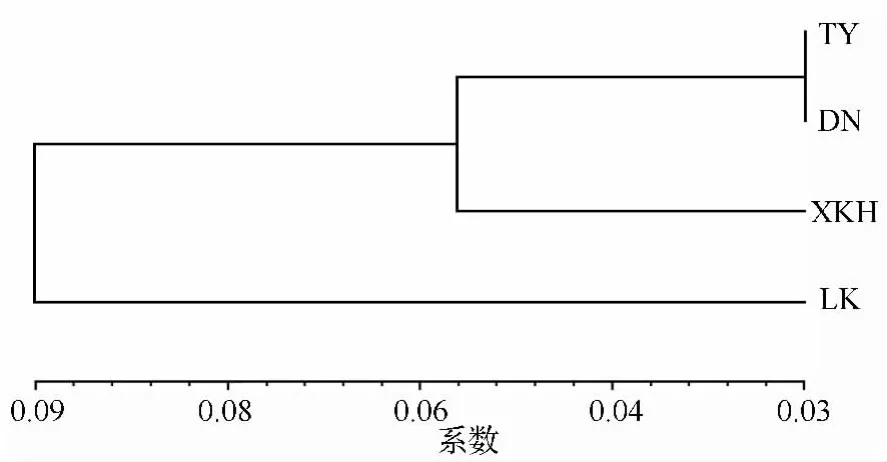
图3 4个野生杏居群间Nei’s遗传距离的UPGMA聚类图
为了阐明供试杏样品之间是否存在基因交流,进行了176个样本基于Nei&Li遗传相似性系数的聚类分析(图4)。从图中可以看出在相似系数0.745处截取聚类线,可将176份样本分成3大组:第1组包含TY居群和DN居群81个西伯利亚杏样本,两居群间存在基因交流,或存在同源性基因。第2组包含XKH居群的XKH1-XKH65的65个辽杏样品,其在0.785处截取聚类线分成5个亚组(亚组Ⅰ、亚组Ⅱ亚组Ⅲ、亚组Ⅳ、亚组Ⅴ),表明XKH居群辽杏具有丰富的遗传变异。第3组包含XKH居群的XKH66-XKH79和LK居群全部样本,包含两个种(辽杏和西伯利亚杏),表明LK居群和XKH居群的亲缘关系较近;其中XKH居群在相似系数0.785处又分为两个亚组(亚组Ⅵ、亚组Ⅶ)。
2.4 遗传多样性指数与气候和地理相关性
将各项遗传参数与相关影响因子进行分析(表5)。可以看出无霜期与各种遗传参数相关性程度很高,相关系数均大于60%;纬度和年均气温与遗传一致度I和遗传距离D有明显相关性,相关系数大于60%。

图4 基于Nei&Li遗传相似性系数的野生杏176个个体的UPGMA聚类图

表5 居群遗传参数与影响因子的相关性
3 讨论
遗传多样性是保护生物学研究的核心之一,是生物各分支学科重要的背景资料,本试验利用ISSR分子标记技术,对我国东北部野生杏进行了遗传多样性分析,表明居群遗传多样性指数(多态性百分率、Nei’s基因多样度、Shannon信息指数)均以兴凯湖居群(XKH)野生辽杏最高,林口居群(LK)野生西伯利亚杏最低,其中XKH居群样品在聚类图中显示0.785处分为7个亚组,表明野生辽杏的遗传多样性程度较高。根据Nei’s基因多样度、Shannon信息指数对野生西伯利亚杏3个居群遗传多样性进行排列得出:TY>DN>LK。居群遗传多样性和影响因子相关分析表明:地理纬度、无霜期、年均气温与遗传多样性相关程度较高。将本研究结果(H=0.2471,Ho=0.3799)与前人对普通杏群体[6]和伊犁野生杏种群[7]研究结果比较,表明东北野生杏遗传多样性程度高于普通杏群体,但低于伊犁野生杏种群。与伊犁野生杏种群[7]居群内遗传分化比较,东北地区野生辽杏和西伯利亚杏种内(居群内)遗传变异较高,具有丰富的种内遗传多样性。居群间的基因流为2.4063,介于伊犁野杏种群和新疆杏品种群之间[7-8]。产生这些变化的原因,一方面是4个居群地理位置不同,导致年均气温、无霜期等气候环境差异,影响了居群间的基因交流;另外是东北地区冬季漫长、气候干寒,春秋两季温度变化剧烈,野生杏为适应恶劣的自然环境而产生的遗传变异的结果。
居群间聚类分析表明:TY居群与DN居群、XKH居群亲缘关系较近,与LK居群亲缘关系最远。样本间的聚类同样表明:TY和DN亲缘关系较近,聚在同一组,两个居群内部存在基因交流。XKH居群部分样品和LK居群聚在一组,表明两居群亲缘关系较近,其中LK居群的LK7号西伯利亚野生杏与XKH居群的XKH72-XKH79号辽杏聚为同一个亚组,表明两地的野生杏亲缘关系较近,而与同居群的其他个体亲缘关系较远。通过地理研究发现林口处于老爷岭和完达山的交接处,兴凯湖西北岸耸峙着完达山,兴凯湖湖岗在历史形成中发生过多次变迁,周围地区的完达山等山脉在历次构造中也发生变化。其亲缘关系较近的原因可能是由于LK与XKH居群在进化历史中存在繁殖体的迁移而产生种间杂交所致,有待继续考证。
物种的稳定性和进化的潜力依赖其遗传多样性,本研究从分子水平证明了东北地区野生辽杏和西伯利亚杏存在丰富的种内遗传多样性变化,应对其进行复杂性状的定位和相关基因研究,并结合形态观察,挖掘更多特异性种质,为新品种选育提供物质基础。同时,调查时发现野生杏所处环境较差,缺乏人为养护管理,建议加强对东北野生杏资源的原生境保护和迁地保护,防止珍惜资源的流失。
[1]张加延,张钊.中国果树志:杏卷[M].北京:中国林业出版社,2003.
[2]谢佳,郭太君,李锋,等.杏92份种质资源的ISSR分析[J].中国果树,2011(2):14-18.
[3]Yeh F C,Yang R C,Boyle T B,et al.Pop gene version 1.31 the user friendly shareware for population genetic analysis[M].Edmonton:Molecular Biology and Biotechnology Centre University of Alberta,1997.
[4]Rohlf F J.NTSYS-PC version 2.10[M].New York:Applied Biostatistics Inc,1994.
[5]Nei M.Estimation of average heterozygosity and genetic distance from a small number of individuals[J].Genetics,1978,89(3):583-590.
[6]苑兆和,陈学森,何天明,等.中国南疆栽培杏群体遗传结构的荧光 AFLP 分析[J].遗传学报,2007,34(11):1037-1047.
[7]何天明,陈学森.利用SSR标记对伊犁河谷野杏种群遗传结构的分析[C]//中国园艺学会第十届会员代表大会暨学术讨论会论文集.北京:中国园艺学会,2005.
[8]何天明,陈学森,高疆生,等.新疆栽培杏群体遗传结构的SSR分析[J].园艺学报,2006,33(4):809-812.
猜你喜欢
作物学报(2022年2期)2022-11-06
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
小读者(2019年20期)2020-01-04
武汉冶金管理干部学院学报(2018年3期)2018-11-15
天然产物研究与开发(2018年10期)2018-11-06
广东农业科学(2017年5期)2017-08-29
植物资源与环境学报(2012年2期)2012-09-06
中学生英语高中综合天地(2008年10期)2008-12-22
