
茯苓多糖对中性粒细胞向肿瘤细胞趋化黏附作用的体外活性研究
2013-09-11 08:13:16李丽娟张德生庄朋伟张艳军张密霞
长春中医药大学学报 2013年4期
李丽娟,张德生,庄朋伟,张艳军,张密霞
(天津中医药大学,天津 300193)
近期研究[1]表明,不同人群粒细胞对肿瘤细胞的杀伤能力不同,而且随着年龄增加,粒细胞活力逐渐下降,肿瘤患者的粒细胞表现为极弱的肿瘤细胞杀伤能力。崔正等[2]报道,肿瘤自然消退/完全消退(SR/CR)小鼠的粒细胞能抵抗多种类型的癌细胞,使肿瘤产生自发消退和/或其中性粒细胞体外能在肿瘤细胞周围形成玫瑰花环样结构,并杀死瘤细胞/移植SR/CR 小鼠粒细胞至普通小鼠,普通小鼠可获得抗肿瘤能力。当机体发生细菌感染时白细胞可被激活,而细菌细胞壁脂多糖是激活白细胞的主要物质,真菌多糖可提高机体的细胞免疫及体液免疫水平[3],故推测某些真菌类多糖可能能通过提高粒细胞杀伤肿瘤细胞的能力从而发挥防治肿瘤作用。茯苓多糖具有调节固有免疫和抗肿瘤作用[4]。本实验从大鼠全血分离中性粒细胞,经茯苓多糖作用后加入体外培养的肿瘤细胞,通过观察中性粒细胞在肿瘤细胞周围形成玫瑰花环样结构的比例,探讨茯苓多糖体外激活中性粒细胞从而抗肿瘤的活性,并对其作用机制进行初步研究。
1 实验方法
1.1 主要试剂与仪器 DT-500(Pharmacia 公司,Lot.No.307041);Ficoll 分离液(Pharmacia 公司,Lot.No.10038229);胎牛血清(Bioind 公司);RPMI 1640(Sigma公司,Lot.No.688600)。RT-PCR 试剂盒(大连宝生物)。核酸蛋白分析仪(Beckman,Du530);基因扩增仪(Hybaid limited Equipment Class,PE-480);凝胶成像分析仪(Bio Image system Gene Company,Gene Genius);茯苓多糖提取物粉末,天津中医药大学赵骏老师提供;Bel-7402 细胞株,上海润成生物科技有限公司;SD 大鼠:天津中医药大学实验动物中心。
1.2 大鼠PMNs 的分离[5]5月龄大鼠1 只,乙醚麻醉,腹主动脉取血8 mL,加入肝素抗凝的试管中,再加入等体积的6%DT-500,混匀,37℃静置40 min,取上清,加等体积生理盐水,1 800 rpm 离心10 min,弃上清,加少量生理盐水重悬后加至装有Ficoll 的离心管上层,2 000 rpm 离心18 min,取PMNs 层,生理盐水洗涤,台盼蓝染色,活细胞数占总细胞数95%以上。
1.3 人肝癌细胞株Bel-7402 培养 将Bel-7402 细胞株以含10%胎牛血清(FBS),1%青、链霉素的RPMI1640培养液接种于75 cm2培养瓶,细胞密度为5×104/mL,隔天换液。待细胞铺满瓶底80%时,以0.25%胰蛋白酶和0.02%EDTA 等体积组成的消化液消化传代,以含10%FBS、1%青、链霉素RPMI1640培养液重悬,调整浓度为1×104/mL,种于6 孔板,每孔2 mL,每组3 复孔,培养24 h 备用。
1.4 茯苓多糖对PMNs 活性的影响 将分离出的PMNs 以8×105个/孔接种于6 孔板,加入茯苓多糖,使其终浓度分别为 62.5、31.25、15.625、7.813、3.906 μg/mL,以等量全培养基作对照。作用2 h 后离心,弃上清,用1 mL 全培养基重悬后加入预先培养有Bel-7402 的6 孔板。共培养4 h 后,在倒置相差显微镜下观察,每孔随机取10 个10×10 倍视野,照像并计数PMNs 在Bel-7402 细胞周围形成玫瑰花环形态的比例,以3 个或3 个以上PMNs 围绕在一个Bel-7402 周围计为一个玫瑰花环。
1.5 茯苓多糖对PMNs 黏附相关基因表达的影响将原代分离的PMNs 以1×106/mL 种于6 孔板,分为空白PMNs 对照组(全培养基孵育2 h)和茯苓多糖激活PMNs组(7.813 μg/mL 茯苓多糖孵育2 h),每组3复孔。2 h 后1 500 rpm 离心5 min,弃上清,每孔加1 mL Trizol 提取总RNA,RT-PCR 法检测黏附相关基因TLR-4、CD11b、CD18、MyD88、MAPK-3、NF-κb 等mRNA 的表达。
引物序列根据Primer3 软件设计,经NCBI 数据库检测,由上海生工生物技术有限公司合成。序列如下:β-actin:上游AGCCATGTACGTAGCCATCC,下游TC TCAGCTGTGGTGGTGAAG(227bp);TLR-4 上游CAGGG AATTAGGCTCCATGA,下游TCCATGACAGAACGGTCAA A(164bp);MyD88 上游GAGATCCGCGAGTTTGAGAC,下游CTGTTTCTGCTGGTTGCGTA(192bp);MAPK-3 上游ATGAAGGCCCGAAACTACCT,下游ATCCAGCTCCATGTCAAAGG(237bp);NF-κb 上 游GCTTTGCAAACCTGGGAATA,下游TCAGGTCCATCTCCTTGGTC(:225 bp);CD11b 上游CATCACCGTGAGTTCCACAC,下游GAGAACTGGTTCTGGCTTGC (174bp);CD18:上游AGTCCCAGTGGAACAACGAC,下 游 GCACTGGGGCTAGCTGTAAG(161bp)。反应条件:94℃,2 min,1cycle;94℃,30sec,58℃,30sec,72℃,30sec,(其中βactin 扩增28 cycles,其余38 cycles)。琼脂糖凝胶电泳,凝胶成像分析仪上应用BIO IMAGING SYSTEM(SYNGENE)照像,以目的基因β-actin 灰度比值进行半定量比较分析。计算方法:目的基因灰度比值=目的基因灰度值/β-actin 灰度值。
1.6 数据统计 采用SPSS 11.5 统计软件进行统计学处理,用单因素方差分析(one-way ANOVA)进行组间比较,所有数据用均数±标准差()表示。
2 实验结果
2.1 不同浓度茯苓多糖对PMNs 活性的影响 见表1。
表1 不同浓度茯苓多糖对PMNs 活性的影响()
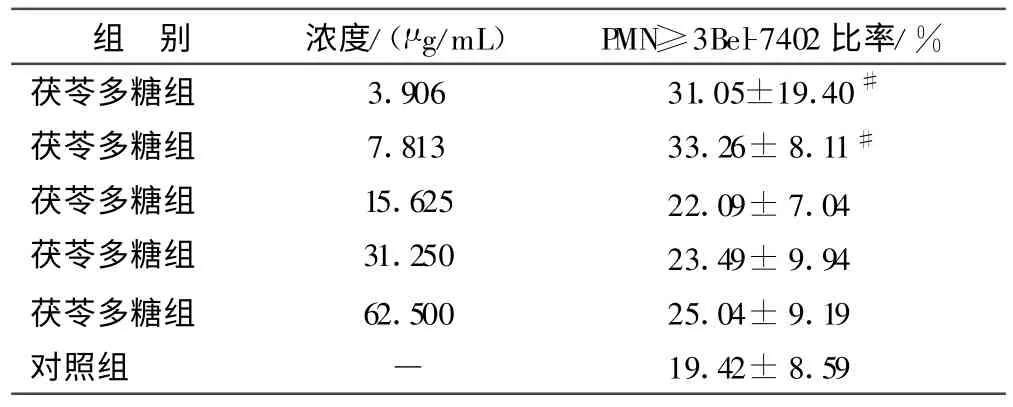
表1 不同浓度茯苓多糖对PMNs 活性的影响()
注:与对照组比较,#P <0.05。
2.2 茯苓多糖对离体PMNs 黏附相关基因表达的影响 见表2。
表2 茯苓多糖对离体PMNs 黏附及活化相关基因表达的影响()
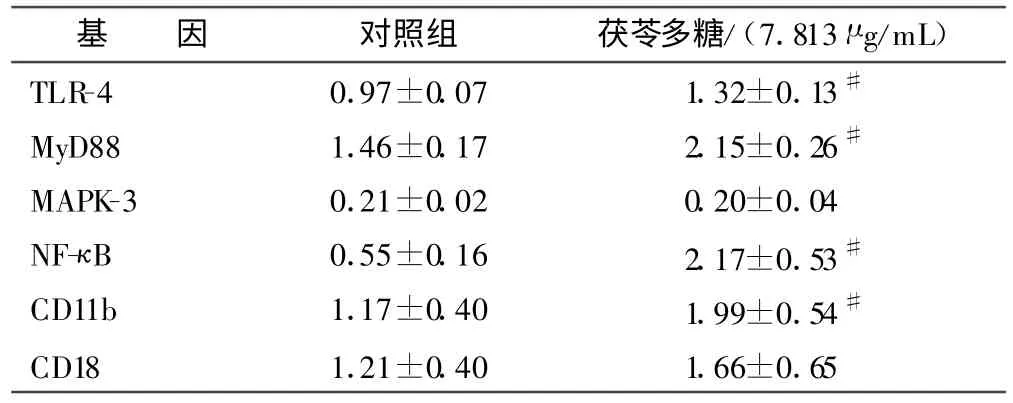
表2 茯苓多糖对离体PMNs 黏附及活化相关基因表达的影响()
注:与对照组比较,#P <0.05。
从表2 可以看出,与对照组比较,茯苓多糖可以增加PMNs 黏 附相 关 基因TLR-4、MyD88、NF-κB、CD11bmRNA 的表达。
3 讨论
慢性炎症与癌症的发生发展密切相关,慢性炎症可导致肿瘤形成,炎细胞的募集也可能抗肿瘤产生和肿瘤进展。研究[6]认为,细胞因子的平衡调节影响炎症浸润的类型和数量及肿瘤进程:肿瘤产生少量或不产生细胞因子或产生过量抗炎因子,限制炎症反应和血管形成,限制肿瘤生长;产生大量的促炎症因子导致炎症反应达到增加血管新生的地步,则促进肿瘤生长;抗炎和促炎因子失衡,导致大量单核细胞或中性粒细胞浸润肿瘤时,可有细胞毒和血管新生停滞,结果肿瘤萎缩
为探讨茯苓多糖提高PMNs 对肿瘤细胞趋化黏附作用的机制,检测了PMNs 炎症相关因子TLR-4、MyD88、NF-κB、CD11b、CD18mRNA 的表达。TLR4 是LPS 的跨膜受体,被LPS 激活后可激活MyD88 依赖性信号转导途径,诱导炎症基因如NF-κB、TNF-α、MAPK等的表达,诱导免疫反应发生。Mac-1 是位于白细胞表面的黏附分子,由CD11b 和CD18 2 个亚基组成。吞噬细胞上的CD11b/CD18 对iC3b 调理的细胞毒性反应需依赖病原体多糖与CD11b 结合,使CD11/CD18 致敏才能触发,这些多糖常含甘露糖、葡萄糖等糖基。而iC3b 调理的肿瘤细胞缺乏适宜的多糖刺激,必须依赖外源性多糖的参与,才能使CD11b/CD18致敏[8]。茯苓多糖主要含岩藻糖、甘露糖、葡萄糖和半乳糖[9],它有可能致敏CD11b/CD18,触发吞噬细胞对肿瘤细胞的毒性反应。本研究发现茯苓多糖能增加PMNs TLR-4、MyD88、NF-κB 及CD11bmRNA 表达,这可能是其促进白细胞黏附于肿瘤细胞的机制之一。总之,本实验研究发现茯苓多糖可以增加PMNs活性,提高PMNs 对肿瘤的趋化黏附作用,其机制与增加PMNs 黏附 相关 基因TLR-4、MyD88、NF-κB、CD11bmRNA 的表达相关。
[1]高艳红,董矜,田亚平.具有抗肿瘤特性的中性粒细胞的筛选技术[J].标记免疫分析与临床,2009,16(5):298-300.
[2]Cui Z,Willingham MC,Hicks AM,et al.Spontaneous regression of advanced Cancer:identification of a unique genetically determined,age-dependent trait in mice[J].Proc Natl Acad Sci U S A,2003,100(11):6682-6687.
[3]张诚,赵慧群,刘江成,等.食药用真菌的生物活性概述[J].吉林中医药,2006,26(10):56-58.
[4]李庆云,张艳军.茯苓多糖抗肿瘤机理研究[J].吉林中医药,2010,30(4):345-347.
[5]张艳军.活血化瘀注射液对大鼠肺微血管内皮细胞与白细胞黏附率的影响[J].天津中医学院学报,2001,20(2):20-22.
[6]Coussens L M,Werb Z.Inflammation and Cancer[J].Nature,2002,420(6917):860-867.
[7]况荣华,周林,傅颖珺.中药以LPS-TLR4 为作用靶点的抗炎机制的研究进展[J].南昌大学学报:医学版,2011,51(10):89-92.
[8]刁飞,朱晓燕,刘宇健.Mac-1 的结构与功能及其在肿瘤免疫治疗中的作用[J].中国癌症杂志,2006,16(3):232-234.
[9]黄灿,王玉明,赵骏.抗肿瘤活性茯苓多糖的提取、纯化与结构分析[J].中草药,2012,43(11):2146-2149.
猜你喜欢
Journal of Traditional Chinese Medicine(2022年5期)2022-10-14 11:38:16
中国民间疗法(2021年14期)2021-08-30 08:24:52
基层中医药(2020年7期)2020-09-11 06:38:04
基层中医药(2018年10期)2018-12-06 09:27:20
天然产物研究与开发(2018年6期)2018-07-09 06:01:28
中成药(2018年3期)2018-05-07 13:34:45
中成药(2018年3期)2018-05-07 13:34:17
医学研究杂志(2015年3期)2015-06-10 06:41:52
中国当代医药(2015年21期)2015-03-01 02:05:08
中成药(2014年11期)2014-02-28 22:29:50
