
辽河保护区7个三裂叶豚草种群果实多样性分析
2013-04-25 09:50:20王学治凌兴泽黄晓星
草业科学 2013年11期
王学治,凌兴泽,翟 强,黄晓星,曲 波
(1.沈阳农业大学生物科学技术学院,辽宁 沈阳 110866; 2.辽河平原森林生态系统定位研究站,辽宁 沈阳 110866; 3.辽宁省辽河保护区管理局,辽宁 沈阳 100054)
辽河是全国七大江河之一,发源于河北省七老图山脉,流经河北省、内蒙古自治区、吉林省、辽宁省,至盘锦注入渤海。辽河保护区是2010年3月经中国共产党辽宁省委员会、辽宁省人民政府批准,依辽河干流设立的狭长区域。辽河保护区始于东西辽河交汇处(铁岭福德店),穿越铁岭、沈阳、鞍山、盘锦 4市,地理坐标为123°55′-121°41′ E,43°02′-40°47′ N,占地面积为1 869.2 km2,是我国第一家大河的河流“划区设局”,流经区域人口较为稠密,土地开发利用程度较高,经济活动强度较大,尤其近年来受到工业化、城市化和新农村建设快速发展的影响,河流生态系统遭到严重破坏,特别是外来入侵植物已严重破坏了辽河保护区的生物多样性。目前,辽河保护区内已发现外来入侵植物27种,其中分布最为广泛的是三裂叶豚草(Ambrosiatrifida)。三裂叶豚草为菊科豚草属植物,一年生高大草本。叶掌状三裂,有时五裂,几乎全部对生[1]。植株高50~120 cm,有的可达170~200 cm,原产北美洲,其生命力、竞争力极强,传播途径多,繁殖系数大。三裂叶豚草分布范围广,危害大,难以治理,已对农业生产和生态环境造成了巨大危害[2]。据资料记载,20世纪50年代在辽宁地区就发现了三裂叶豚草,分布在以沈阳、铁岭两处为中心的辽河流域[1],近年来已扩散到整个辽宁乃至吉林、黑龙江、河北、北京、山东,甚至上海也发现三裂叶豚草[3]。
表型多样性能够在形态水平上反映物种遗传多样性,它体现了群体遗传适应生态环境压力的广泛程度[4]。植物果实是植物生长周期中的重要繁殖器官,其对种群的繁衍、扩张和抵抗不良环境有重大意义,而果实的传播是植物种群拓展其生存空间的重要保证[5-7]。Fenner[8]研究表明,除了表型差异之外,同一物种个体间果实的平均质量虽差别不大,但果实的质量在物种间存在极大的差异。马绍宾等[9]在研究药用植物桃儿七(Sinopodophyllumhexandrum)不同种群种子产量时发现,对于同一物种的不同地理种群,果实的平均质量存在一定差异,且这种差异与该种群的群落特征和生境条件有关。为了揭示三裂叶豚草的传播与扩散机制,关广清[1]曾对三裂叶豚草的形态进行了初步研究,发现其在各地区表型差异较大。
本研究对辽河保护区的三裂叶豚草种群果实形态和质量进行研究,探讨三裂叶豚草果实多样性,试图揭示辽河保护区内三裂叶豚草的遗传变异与其传播途径之间的关系,为三裂叶豚草的防治提供依据。
1 材料与方法
1.1材料 2011年秋季,沿辽河保护区,选取7个有代表性的三裂叶豚草种群,各种群间距离超过20 km。在各种群内选取50株三裂叶豚草,株间距大于1 m,收集每株植物的成熟果实,自然风干,备用。7个三裂叶豚草种群的具体信息和位置如表1和图1所示。

表1 三裂叶豚草种群位置信息Table 1 Location information of populations of giant ragweeds
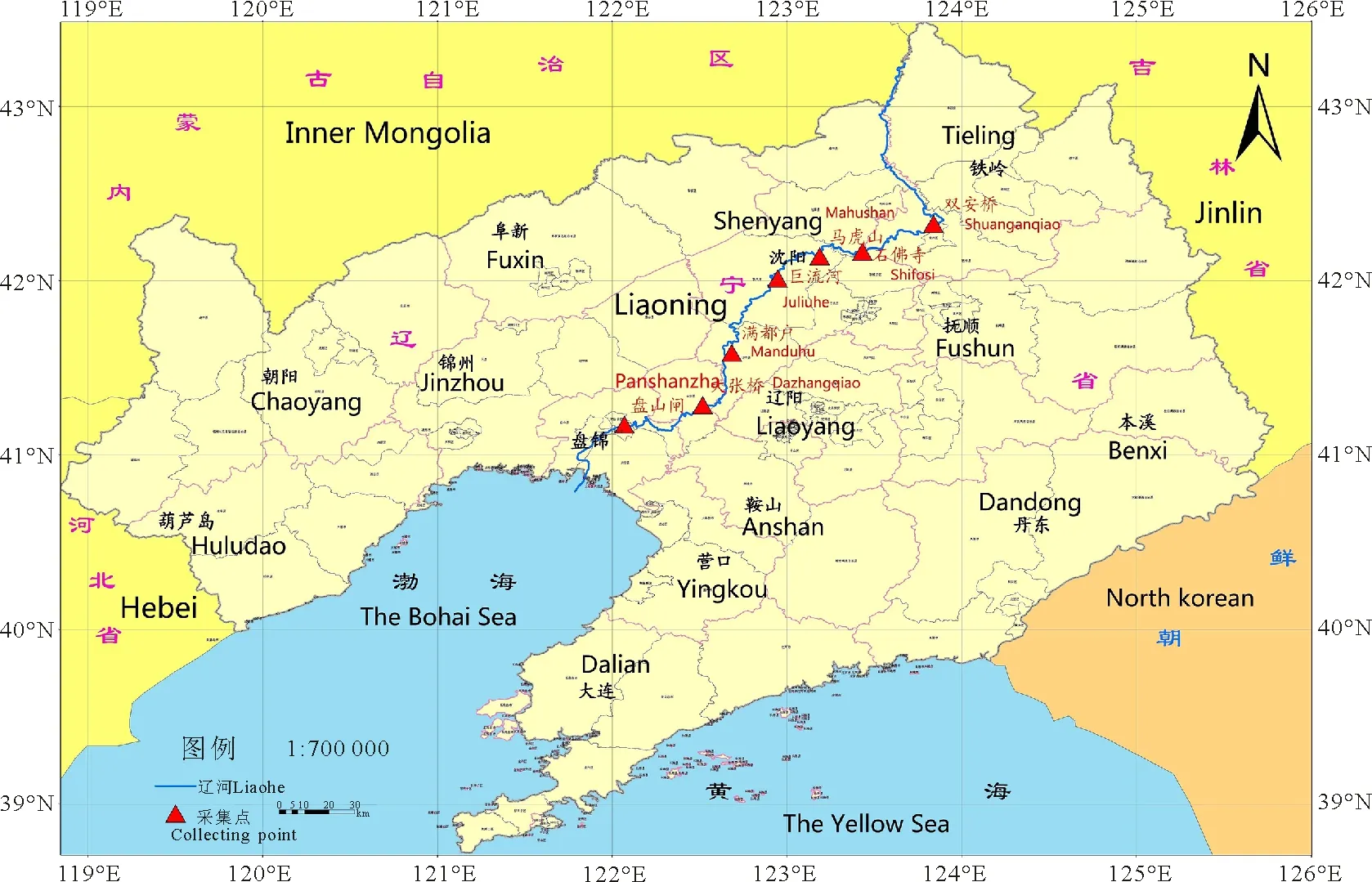
图1 三裂叶豚草种群分布图Fig.1 The populations distribution of giant ragweeds
1.2方法
1.2.1三裂叶豚草果实千粒重测定 取每株植物成熟果实各60粒,每个种群计3 000粒,混合后均分为3组,每组1 000粒,称量,重复3次,计算其平均值。
1.2.2三裂叶豚草果实形态特征测定 每个种群随机取出30粒果实进行测定,主要指标为正面观面积、长(果实纵轴长度)、宽(果实横轴长度)、周长以及顶刺、主刺数目、刺长等(图2),参考Sako等[10]的方法,略作改动:根据同一张图片中像素密度和大小成正比,制作1 cm×1 cm的对照矩形图片,将三裂叶豚草果实与对照矩形置于同一平面,利用数码相机(松下GF3,像素为1 200万)对其拍照,利用Photoshop CS4软件测定各指标像素数量,根据与标准对照的像素比例,计算出果实的正面观面积、周长、刺长等。从果实正面正上方观察,把果实分为四边形、五边形、六边形和椭圆形4种形状。
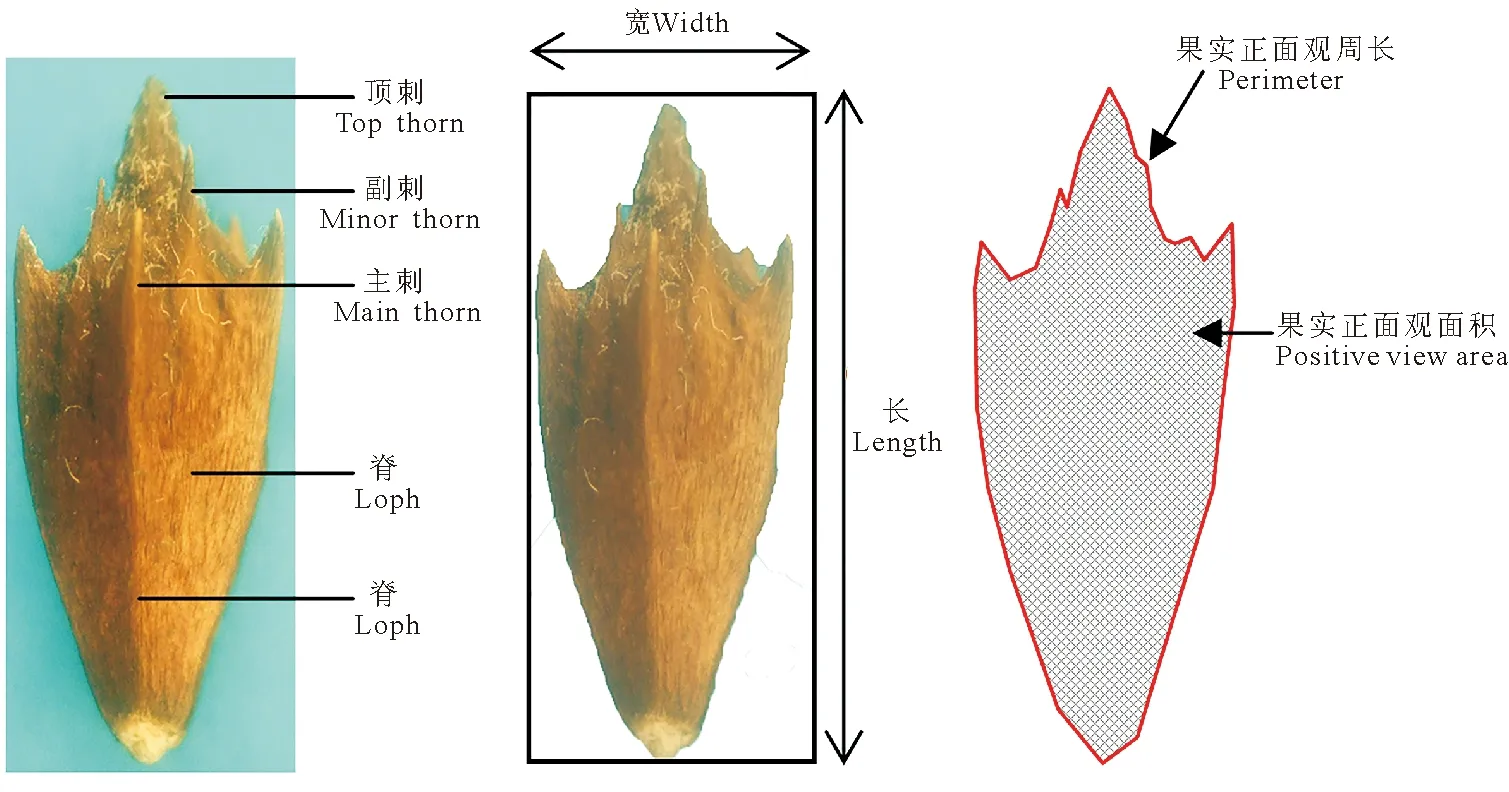
图2 三裂叶豚草果实形态指标图Fig.2 The fruit shape of giant ragweeds
1.2.3数据处理 采用SPSS软件进行聚类分析[11-12],聚类方法采用平方Euclidean法。采用DPS (Data Processing System 9.50)软件对数据进行差异显著性分析。
2 结果与分析
通过对辽河保护区内三裂叶豚草7个种群的果实特征进行观察与测定发现,在果实的正面观面积和周长方面,盘山闸种群的正面观面积最大,平均为0.227 cm2,周长平均为2.117 cm;石佛寺种群正面观面积和周长最小,分别为0.133 cm2和1.507 cm;其他种群果实的面积在0.149~0.173 cm2,周长在1.623~1.892 cm。巨流河种群与其余三裂叶豚草的果实形状不同,其果实上端呈放射状。盘山闸种群的果实千粒重最大,平均为24.200 g;石佛寺种群最小,平均为14.100 g;其他种群果实千粒重在15.500~19.600 g,不同位置种群果实千粒重差异显著。统计结果显示,石佛寺种群果实的正面观面积和周长显著低于其他种群(P<0.05)。
果实的长宽比反映果实的形状[10],本试验中三裂叶豚草种群果实的长宽比不同地区间均差异显著(P<0.05)。马虎山种群和巨流河种群果实长宽比分别为1.399和1.277,较接近;而大张桥种群、石佛寺种群和盘山闸种群果实长宽比较接近,介于1.483~1.595(表2)。
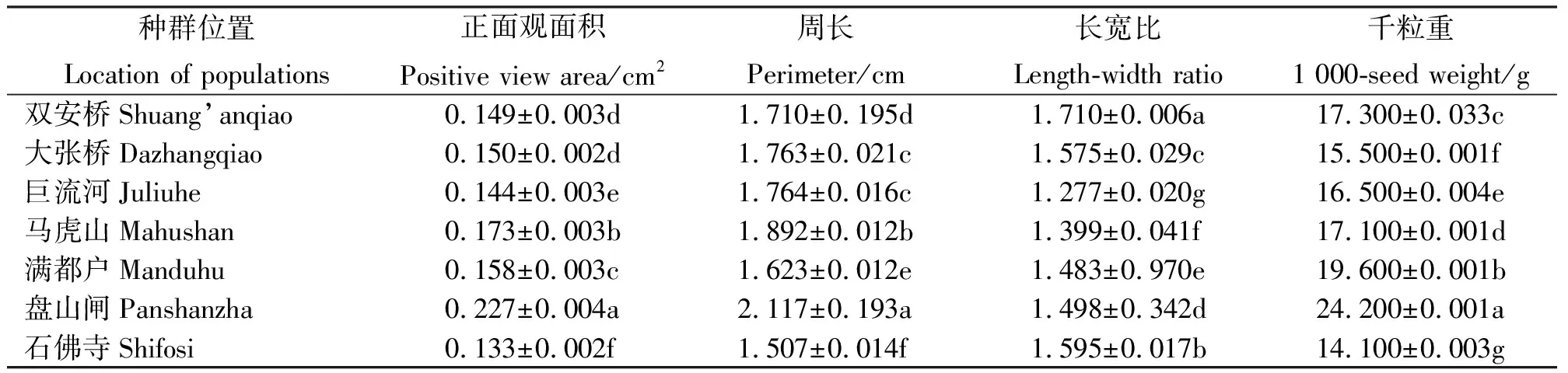
表2 辽河流域各地区三裂叶豚草果实特征Table 2 The fruit characteristics of giant ragweeds in Liaohe River
在三裂叶豚草7个种群中,马虎山种群果实顶刺最长,平均为0.101 cm;其次为巨流河种群,其果实顶刺长平均为0.098 cm。主刺最长的为盘山闸种群,平均为0.171 cm;其次为双安桥,平均为0.155 cm(表3),其中,石佛寺种群的果实没有明显的主刺。三裂叶豚草7个种群果实均没有明显的副刺。巨流河种群和马虎山种群果实都有6个主刺,而且正面观形状接近六边形(图3)。满都户种群果实形状近似椭圆形,既没有主刺也没有顶刺,而石佛寺种群果实没有主刺,大张桥种群和盘山闸种群都有5个主刺,双安桥有4个主刺。统计结果显示,满都户种群果实的顶刺长、顶刺数、主刺长、主刺数均为0,且顶刺长和顶刺数显著低于其他种群(P<0.05)。
7个种群中,大张桥和盘山闸种群果实形状一致,均为五边形;巨流河和马虎山种群果实形状一致,均为六边形;满都户和石佛寺种群果实形状一致,均为椭圆形;双安桥果实形状为四边形。
利用果实长宽比和刺数对三裂叶豚草7个种群进行综合聚类,发现大张桥种群和盘山闸种群聚类最近,巨流河种群和马虎山种群聚类最近,满都户种群和石佛寺种群在另一个分支上聚类最近(图4)。
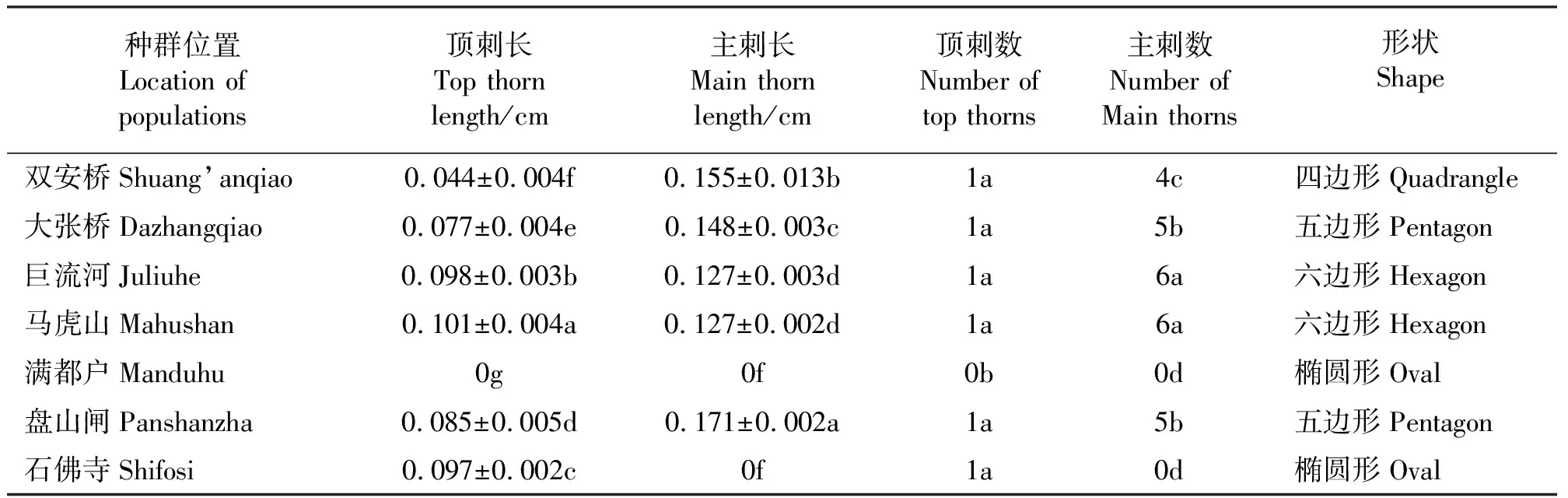
表3 辽河流域各地区三裂叶豚草果实刺的特征Table 3 The prick characteristics of giant ragweeds fruit in Liaohe River

图3 三裂叶豚草果实形态Fig.3 The fruit shape of giant ragweeds
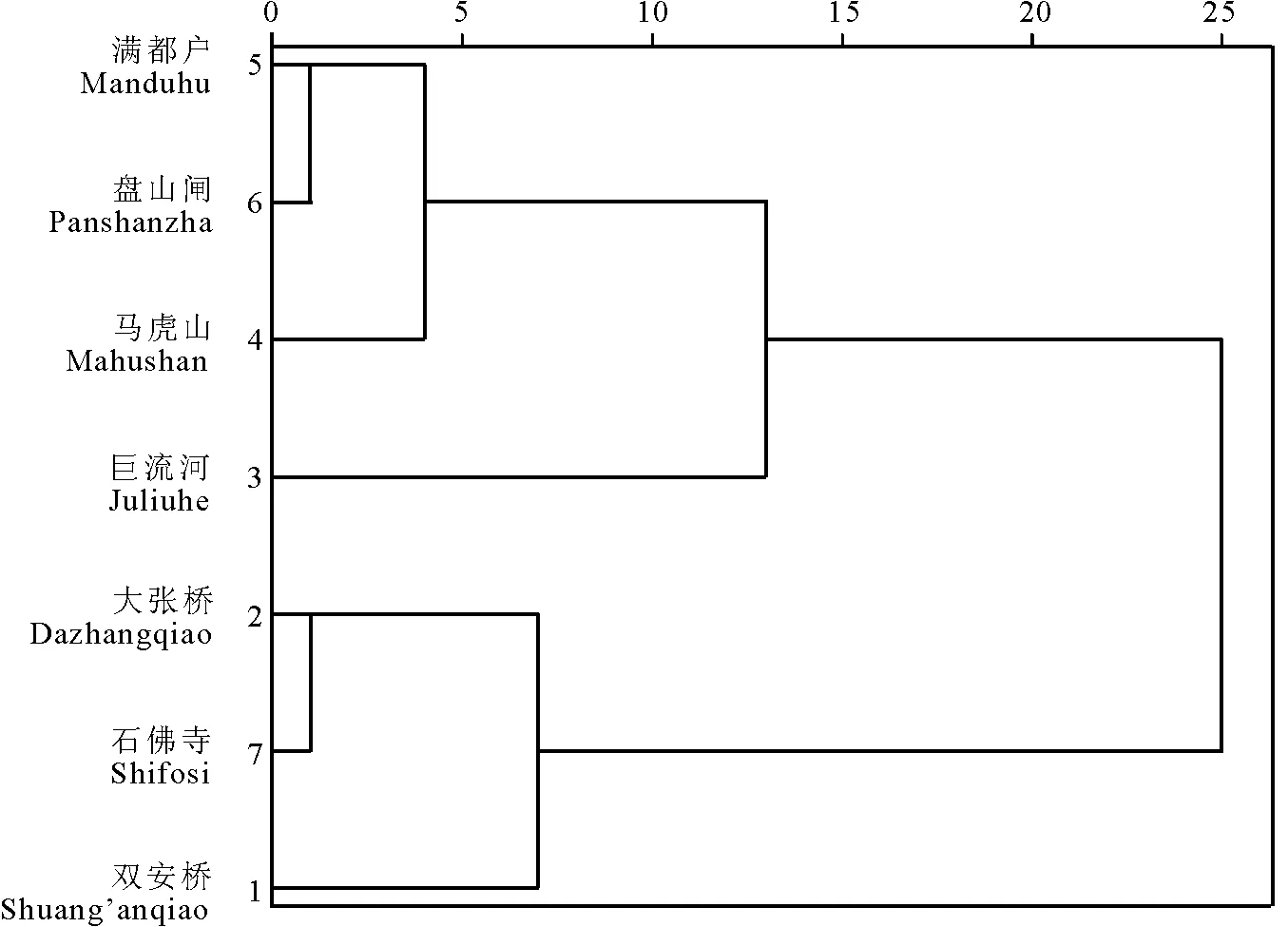
图4 三裂叶豚草果实形态聚类图Fig.4 The clustering diagram of the fruit shape of giant ragweeds
3 讨论与结论
本研究表明,三裂叶豚草果实质量、面积和刺长指标,在同一种群内差异较大,具体表现在同一种群内果实有大有小,刺有长有短,没有规律性,并且果实的大小受当地降水、光照等影响很大,不能反映种群间的遗传性;不同三裂叶豚草种群间的果实长宽比和果实刺的数量,即果实的形状,在同一种群内较一致,有一定规律,可作为反映三裂叶豚草种群间果实多样性的主要依据,用来分析三裂叶豚草果实遗传多样性。
7个种群之中,大张桥种群和盘山闸种群果实形状接近,表现为主刺的数量有5个,并且它们的地理位置距离也较近,相距39 km。马虎山种群和巨流河种群果实形状接近,表现为主刺的数量有6个,且地理位置相距24 km。满都户种群和石佛寺种群果实形状十分接近,在聚类图上同在另一分支,但距离较远,相距101 km,由此推断,可能是因为这些种群之中,石佛寺和满都户在辽河南岸,与辽河北岸的其他种群产生了地理隔离,加上两岸的生境不同,导致其果实形状圆润,没有明显的主刺。石佛寺种群虽然与马虎山种群距离很近,但是这两个种群分布在辽河两岸,产生了地理隔离,基因交流机会少,所以并没有聚在一组。
本研究结果表明,辽河保护区内三裂叶豚草不同种群的果实表观形态差异程度与其地理位置有关。
Fennell等[13]和Chun等[14]研究表明,果实形态相近的种群可能最初由一个种群分化而来。由此推断,果实最为相似的巨流河种群和马虎山种群,它们地理位置靠近,可能种群间有基因交流,并且它们间有国道101和国道304等道路,交通十分畅通发达,可以由交通工具作为传播媒介,使最初同一种群的果实在不同地域形成新的种群,从种群发展的历史来看,可能属于种群分割(Population subdivision)[15]。 有研究表明,不同种群之间通过道路运输会有基因交流[16-17]。由此可见,辽河保护区内三裂叶豚草可能通过交通运输进行传播。建议辽河保护区防治三裂叶豚草以道路为主线,以线带面,深入保护区,采取相关措施控制三裂叶豚草,并在重要交通区域建立监测区,采取预警措施,防止三裂叶豚草再度发生。
[1] 关广清.豚草和三裂叶豚草的形态特征和变异类型[J].沈阳农学院学报,1985,16(4):9-17.
[2] 曲波,吕国忠,杨红,等.辽宁省外来有害生物——杂草调查[J].沈阳农业大学学报,2006,37(4):578-592.
[3] 达良俊,王晨曦,田志慧,等.上海佘山地区外来入侵物种三裂叶豚草群落的新分布[J].华东师范大学学报,2008(3):36-40.
[4] Gland A,Lichter R,Schweiger H G.Genetic and exogenous factors affecting embryogenesis in isolated microspore cultures ofBrassicanapus[J].Plant Physiology,1988,132:613-617.
[5] 杨逢建,张衷华,王文杰,等.八种菊科外来植物果实形态与生理生化特征的差异[J].生态学报,2007,27(2):442-449.
[6] 徐汝梅,叶万辉.生物入侵理论与实践[M].北京:科学出版社,2003.
[7] 沈有信,刘文耀.长久性紫茎泽兰土壤种子库[J].植物生态学报,2004,28(6):768-772.
[8] Fenner M.Seed Ecology[M].New York:Chapman and Hall Ltd.,1985:1-56.
[9] 马绍宾,姜汉侨,黄衡宇.药用植物桃儿七不同种群种子产量初步研究[J].应用生态学报,2001,12(3):363-368.
[10] Sako Y,Regnier E,Daoust T.Computer image analysis and classification of giant ragweed seeds[J].Weed Science,2001,49:738-745.
[11] 张文彤,邝春伟.SPSS统计分析基础教程[M].第2版.北京:高等教育出版社,2011:251-261.
[12] 李春喜.生物统计学[M].北京:科学出版社,2005.
[13] Fennell M,Gallagher T,Osborne B.Patterns of genetic variation in invasive populations[J].Biological Invasions,2010,12:3973-3987.
[14] Chun Y J,Nason J D,Moloney K A.Comparison of quantitative and molecular genetic variation of native vs.invasive populations of purple loosestrife (LythrumsalicariaL.,Lythraceae)[J].Molecular Ecology,2009,18(14):3020-3035.
[15] 樊龙江.生物信息学札记(第3版)(网络教材)[EB/OL].[2013-01-04].http://ibi.zju.edu.cn/bioinplant/
[16] 黄文坤,郭建英,万方浩,等.紫茎泽兰群体遗传多样性及遗传结构的AFLP分析[J].农业生物技术学报,2007,15(6):992-1000.
[17] Genton B J,Shykoff J A,Giraud T.High genetic diversity in French invasive populations of common ragweed,as a result of multiple sources of introduction[J].Molecular Ecology,2005,14:4275-4285.
猜你喜欢
新少年(2023年5期)2023-05-06 20:04:57
草食家畜(2022年5期)2023-01-02 02:53:26
选煤技术(2021年3期)2021-10-13 07:33:24
源流(2021年11期)2021-03-25 10:32:07
理财·收藏版(2019年2期)2019-09-10 07:22:44
Journal of Oceanology and Limnology(2018年6期)2018-12-22 07:00:28
水利规划与设计(2017年9期)2017-12-20 08:24:57
魅力中国(2016年38期)2017-05-27 13:25:31
新农业(2016年19期)2016-08-16 12:00:18
水利建设与管理(2014年4期)2014-07-28 07:44:20
