
不同种群香根草叶生长分析
2013-03-14 08:07:58彭雪梅刘金祥杨允菲
草业科学 2013年7期
彭雪梅,刘金祥,杨允菲
(1.贵州师范学院实验教学管理中心,贵州 贵阳 550018; 2.湛江师范学院热带草业科学研究所,广东 湛江 524048;3.东北师范大学草地研究所 植被生态科学教育部重点实验室,吉林 长春 130024)
自从20世纪70年代初建立了植物种群构件结构理论以来,植物种群构件的研究一直成为植物种群生态学研究领域的前沿课题[1]。叶是植物重要的物质生产器官,在植物和植物种群的生活史中具有重要的作用,其动态特征对于深入了解种群变化规律及种群对环境调节的生物生态机理有重要理论和现实意义。
香根草(Vetiveriazizanioides)是原产热带的多年生禾草,具有纵深发达的根系和坚硬的茎秆,是一种优良的环境改良与生态恢复型植物。目前,在非洲大陆、东南亚、南美和中国等地都发现了香根草的自然分布[2],在我国广东和海南发现有野生香根草的存在[3]。香根草营养分蘖繁殖力强,可进行有性繁殖,但有性繁殖力较弱。一年四季都在生长,3月进入迅速生长时期,11月依然在分蘖。分蘖株抽穗开花后开始枯萎死亡,11月到翌年2月,停止拔节,营养分蘖株生长缓慢。有关香根草的研究主要集中在种子的生产和萌发[4-5]、保持水土[6-7]、稳固堤坝[8]、与其他作物混播的影响[9-10]、重金属抗性[11-12]、污染地的恢复[13]、光合生理生态[14-16]以及香根油的成分、作用和提取[17]等方面。国内研究人员做过一些植物种群统计分析的研究[18-19],但有关香根草叶的研究未见报道。
本研究以野生栽培驯化生长1年的香根草种群、引种生长1年的香根草种群、吴川多年生野生香根草和野生栽培驯化生长3年的香根草种群的分蘖株为研究对象,分析4个样地香根草种群的叶,以期揭示引进种与当地野生种及野生驯化不同年限的香根草种群叶的生长规律,为香根草的广泛应用提供科学累积和基础。
1 材料与方法
1.1样地概况 本研究在广东省吴川市野生香根草草地(21°28′~21°35′ N,110°45′~110°55′ E)和湛江市湛江师范学院院内试验田(21°11′ N,110°23′ E)进行。两地都属热带季风气候。吴川地区累年日照时数1 998 h,年均温23.2 ℃,年最高温度38.5 ℃,年最低温度2.7 ℃,年均降水量1 608.3 mm,年均蒸发量1 681.2 mm,年均相对湿度80%。野生香根草种群分布在吴川县东北部与茂名、电白两县交界的冲积平原上,面积6 670余hm2,群落组成以香根草和牛鞭草(Hemarthriaaltissima)占绝对优势[3]。湛江市累年日照时数1 935 h,年均温23.2 ℃,年最高温度38.1 ℃,年最低温度2.8 ℃,年均降水量1 617.8 mm,年均蒸发量1 610.3 mm,年均相对湿度81%[20]。
样地1(P1)为湛江师范学院院内试验田野生栽培驯化生长1年的香根草试验小区,于2004年8月末单株移栽吴川野生香根草(株距35 cm)建成。样地2(P2)为湛江师范学院院内试验田引种生长1年的香根草试验小区,于2004年10月单株栽种购买引进种香根草(株距35 cm)建成。样地3(P3)为吴川野生香根草草地,多生长在水边,有当地黄牛放牧,生长稀疏。样地4(P4)为湛江师范学院院内试验田野生栽培驯化生长3年的香根草试验小区,于2002年从吴川野生香根草地挖取成丛栽种建成。
1.2研究方法 取样时间在5月初,此时两地香根草都已开始拔节,个别分蘖株已经抽穗,调查各样地香根草构件生长状况,分析环境条件和品种对香根草构件影响的差异。分别在样地1、2、3、4随机取完整分蘖株50株(从分蘖节处剪下)和不同叶位叶片50枚带回实验室。分蘖株随机测量30株(3号样地样本在处理过程中损失2株)的绝对高度和叶片数,随机测量30枚新鲜叶的叶长和叶宽,作标记烘干后对逐个分蘖株、叶片称重。用SPSS对数据进行统计分析,用Excel绘图。取相关程度最高的函数作为刻画模型。叶片生物量为单个分蘖株叶片总生物量,叶片生物量比例为分蘖株叶片的生物量与该分蘖株生物量的比。
2 结果与分析
2.1香根草种群叶特征 在随机测量的分蘖株中,4个样地香根草绝对高度存在差异(表1)。各样地香根草平均高度的顺序为P4>P3>P1>P2。4个种群高度的变异系数分别是34.46%、34.82%、30.49%和19.87%,种群间变异系数为43.30%,种群间变异系数大于种群内。方差分析(F=54.756)表明,种群间差异显著(P<0.05)。多重比较表明,P4显著高于其他种群,P2显著低于其他种群。
4个样地分蘖株的叶片生物量比例(表1)中,P2叶片生物量比例最大,P4叶片生物量比例最小。4个种群叶片生物量比例变异系数分别是12.43%、7.20%、9.35%和12.98%,种群间变异系数为18.81%。方差分析(F=86.708)表明种群间差异显著(P<0.05)。多重比较表明,除P1和P2之间无显著差异外,其他各种群两两之间都存在显著差异。
2.2叶生物量生长格局 各样地香根草分蘖株叶片生物量排序为P4>P3>P1>P2(表1)。各种群叶片生物量的变异系数分别是85.12%、121.30%、88.39%和29.03%,种群间变异系数为87.04%。方差分析(F=41.221)表明,种群间差异显著(P<0.05)。多重比较表明,P4与其他种群差异显著。4个样地分蘖株叶片生物量均随分蘖株高度增加呈幂函数形式增加(图1)。虽然种群间存在一定差异,但叶片生物量随分蘖株高度增加的趋势是相同的。
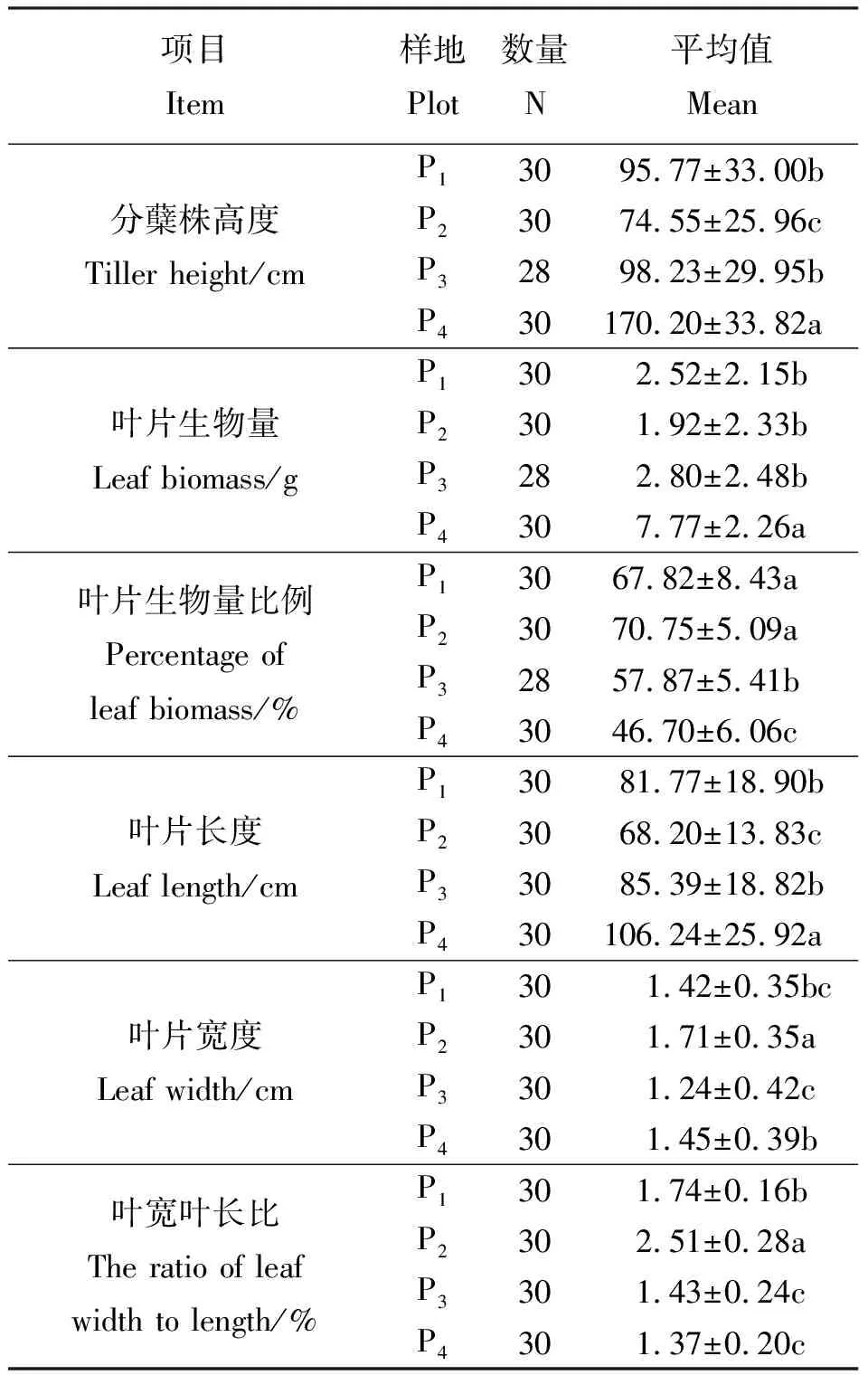
表1 4个样地样本观测量统计分析Table 1 Description of four plots measure data
2.3叶片的异速生长规律 各样地测量的30枚的叶片长度(表1)中,P2叶片长度最小,P4叶片长度最大。4个种群的叶长变异系数分别为23.12%、20.28%、22.04%和24.40%,种群间叶长变异系数为27.99%,种群间变异系数大于种群内。方差分析(F=18.885)表明种群间差异显著(P<0.05)。多重比较(表1)表明,在测量的叶片中,P1与P3之间差异不显著(P>0.05),其他各种群之间均差异显著(P<0.05)。
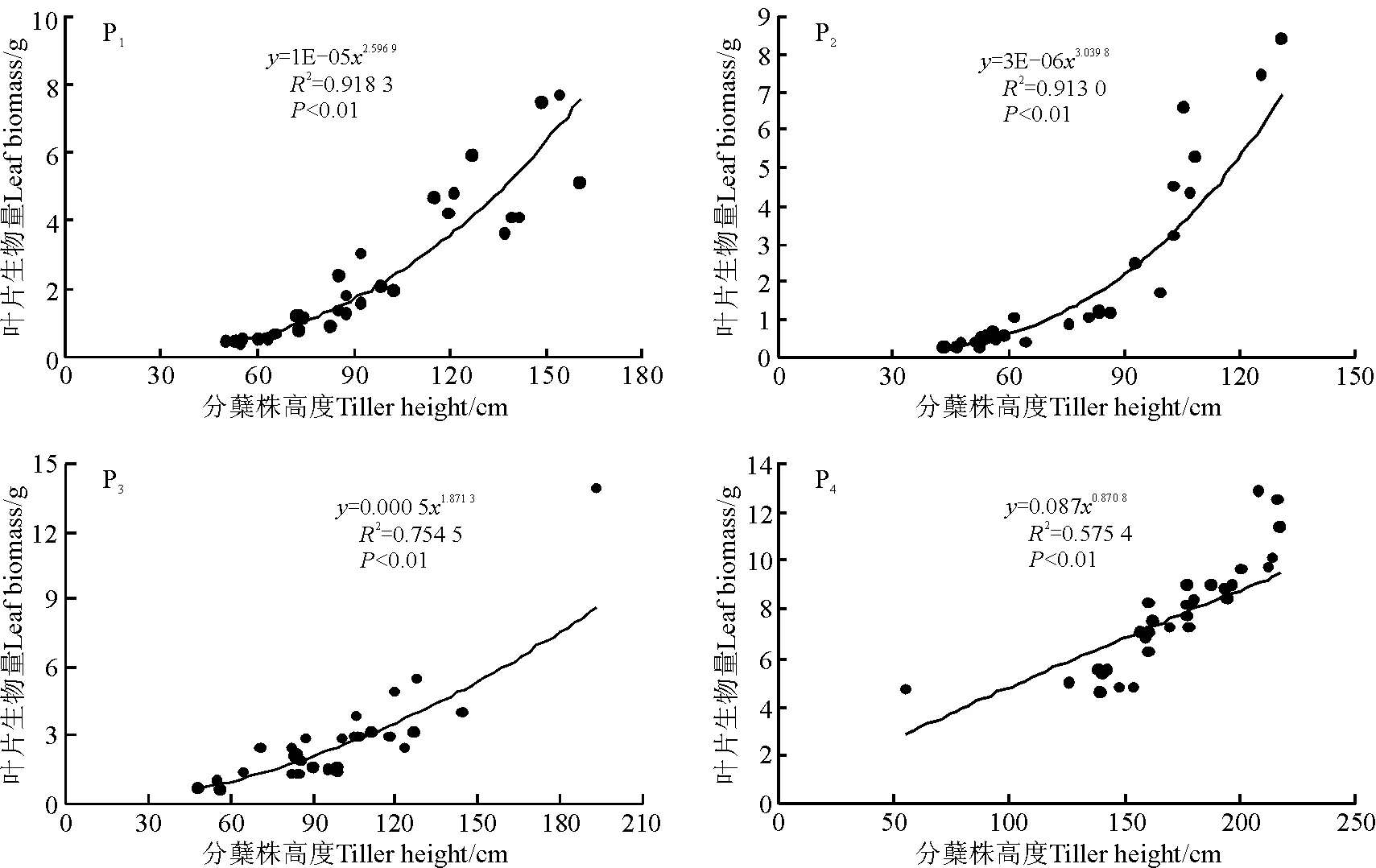
图1 4个样地香根草种群营养分蘖株叶片生物量与分蘖株高度观测值及拟合曲线Fig.1 Observed data and simulated curves on the relationships between leaf biomass and tiller height of four Vetiveria zizanioides populations
对于叶片宽度,P2叶宽最大,P3叶宽最小。4个种群变异系数分别是24.63%、20.47%、34.34%和27.03%,种群间变异系数是28.31%。野生种群与引种种群叶片宽度存在显著差异。4个种群的叶宽叶长比的变异系数分别为9.27%、11.31%、16.84%、14.90%,种群间变异系数为28.80%,种群间变异系数远大于种群内。P2叶宽叶长比最大,相同的叶片长度,P2叶片最宽。P1叶宽叶长比显著大于P3,P4和P3叶宽叶长比无显著差异(P>0.05)。各种群叶宽均随着叶长的增加呈幂函数形式增加(图2)。
3 讨论与结论
由于自然环境和品种的差异,香根草叶种群特征也存在一定的差异。野生种群和引进种群的香根草高度差异显著。栽培驯化3年香根草与多年生野生香根草种群分蘖株高度差异显著。原因可能是栽培驯化3年的种群受外界干扰小,且地理位置更接近热带,春天温度回升稍早,温度相对较高,因此生长较为迅速,分蘖株较高。吴川当地多年生野生香根草种群前一年秋季被放牧黄牛采食,分蘖株相对矮小,对第2年分蘖株高度产生了影响。栽培驯化1年香根草种群移栽时间较短,虽然生长空间充分,与野生草地相比受外界干扰小,但前1年停止分蘖时,分蘖株都比较矮小,因此,翌年5月份,分蘖株高度与栽培驯化3年香根草种群存在显著差异。
从叶片生物量比例上看,生长了3年的种群和生长1年的种群叶片生物量与分孽株生物量的比例有显著差异(P<0.05)。栽培驯化1年和引种1年香根草种群之间无显著差异。可以认为这两个种群之间叶生物量比例不存在显著差异。由于栽培驯化1年和多年生野生种群与栽培驯化3年香根草种群已拔节分蘖株数量存在差异,因此叶生物量比例也有显著差异,生长环境和生长年限对叶生物量比例有影响。叶片生物量的比例的排列顺序与分蘖株高度排序相反,随分蘖株高度的增加和节数的增多,叶生物量比例在样本间逐渐下降,茎生物量所占比例逐渐升高。
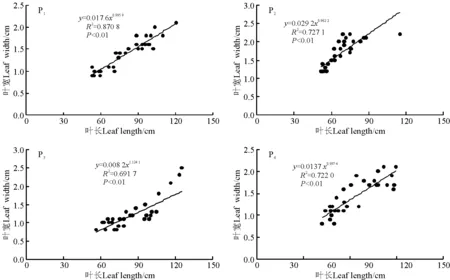
图2 4个样地香根草营养分蘖株叶宽与叶长的观测值及拟合曲线Fig.2 Observed data and simulated curves on the relationship between leaf width and length of four Vetiveria zizanioides populations
香根草种群中,栽培驯化3年香根草种群水热条件好于多年生野生香根草种群,且多年生野生香根草受黄牛放牧影响,因此,环境对不同种群叶的生物量产生影响,栽培驯化3年香根草种群与其他种群叶片生物量均有显著差异,其他3个种群之间无显著差异。4个样地香根草叶片生物量与分蘖株高度呈幂函数增长。4个样地有着趋同的叶生物量生长格局。有研究用R2来估计异速生长规律由遗传因子控制,用1-R2来估计随机环境的影响,则4个样地香根草分蘖株叶片生物量与分蘖株高度的异速生长中,有57.54%~91.83%是由遗传因子控制的,有8.17%~42.46%是由随机环境来影响的[18]。
香根草种群叶片长度的分析比较表明,栽培驯化1年和多年生野生香根草种群的叶片长度无显著差异,其他种群之间差异显著。栽培驯化3年香根草种群由于生长环境条件好,其叶片长度要大于多年生野生香根草,因此,环境对叶片长度有影响,生长条件好,水分充足土壤肥沃,叶片会生长得更长,从而接受更多的光照,生产更多的物质。野生品种和引种品种叶片宽度存在显著差异,引种品种叶片较宽。栽培驯化1年与引种香根草两种群叶宽叶长比有显著差异。栽培驯化3年香根草与多年生野生香根草叶宽叶长比差异不显著。说明虽然栽培驯化多年野生种香根草生长环境有所改变,但并未使其种的叶宽叶长比发生变化。4个种群叶片宽度均随叶片长度呈幂函数增加,说明4个样地香根草有着趋同的叶生长过程。不同叶位叶4个样地均为随机取样,因此误差相同,可忽略不计。
本研究表明,春季分蘖株开始生长的越早,叶片生长越多,当年物质生产量则越大。香根草在前一年秋天的生长对翌年春天的生长及拔节有着重要作用,生长多年分蘖丛比生长1年分蘖丛生长旺盛。野生香根草比引种香根草生活力强。
[1] 杨允菲,李建东.松嫩平原人工草地羊草和野大麦叶种群的趋同生长格局[J].草业学报,2003,12(5):38-43.
[2] 夏汉平,敖惠修.海南野生香根草资源调查报告[J].热带农业科学,2001(2):40-43.
[3] 夏汉平,敖惠修.中国野生的香根草种及其保护与分类问题[J].生物多样性,1998,6(4):292-297.
[4] Verma S,Trivedi K C,Shrivastava D K.A note on seed germination in vetiver[J].Indian Perfumer Bharatiya Gandhika,1995,39(3):131.
[5] Parihar S S,Mal B,Shankar V.Seed production and germination in vetiver grass (Vetiveriazizanoides) [J].Tropical Grasslands,1998,32(3):173-177.
[6] 宋州俊,陈富良,郑勇.香根草等高绿篱在大别山南麓黄棕壤坡耕地水土保持应用的研究[J].亚热带水土保持,2012,24(4):4-7.
[7] 夏汉平,敖惠修,刘世忠.香根草生态工程应用于公路护坡的效益研究[J].草业科学,2002,19(1):52-56.
[8] Hegde B R.Vetiver grass for soil and water conservation, land rehabilitation and embankment stabilization[J].Journal of Plantation Crops,1997,25(1):118.
[9] Maheshwari S K,Sharma R K,Gangrade S K.Intercropping of vetiver (VetiveriazizanioidesL.) with traditional crops in shallow black soils[J].Indian Perfumer Bharatiya Gandhika,1997,41(1):5.
[10] 陈凯,胡国谦,饶辉茂,等.红壤坡地柑桔园栽植香根草的生态效应[J].生态学报,1994,14(3):249-254.
[11] 夏汉平,束文圣.香根草和百喜草对铅锌尾矿重金属的抗性与吸收差异研究[J].生态学报,2001,21(7):1121-1129.
[12] 杨兵,蓝崇钰,束文圣.香根草在铅锌尾矿上生长及其对重金属的吸收[J].生态学报,2005,25(1):45-50.
[13] Prasad A,Singh D V.Effect of sodium-adsorption ratio of irrigation water on soil sodication and yield and on cation accumulation of vetiver (Vetiveriazizanioides) [J].Indian Perfumer Bharatiya Gandhika,2000,44(4):291-298.
[14] 努扎艾提·艾比布,刘云国,宋华晓,等.重金属Zn、Cu对香根草生理生化指标的影响及其积累特性研究[J].农业环境科学学报,2010,29(1):54-59.
[15] 刘金祥,王铭铭.淹水胁迫对香根草生长及光合生理的影响[J].草业科学,2005,22(7):71-73.
[16] 刘金祥,李文送,刘家琼.模拟光条件下有性繁殖香根草光合生理的研究[J].生态学杂志,2005,24(4):390-394.
[17] Weyerstahl P,Marschall H,Splittgerber U.New sesquiterpene ethers from vetiver oil[J].Liebigs Annalen:Organic and Bioorganic Chemistry,1996(7):1195-1200.
[18] 杨允菲,李建东.松嫩平原不同生境芦苇种群分株的生物量分配与生长分析[J].应用生态学报,2003,14(1):30-34.
[19] 周婵,杨允菲.松嫩平原两个生态型羊草叶构件异速生长规律[J].草业学报,2006,15(5):76-81.
[20] 胡异.湛江自然环境与生态[M].湛江:湛江市政协人口资源环境委员会,2003:1-164.
猜你喜欢
贵州林业科技(2021年4期)2021-12-27 02:39:54
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
安徽农学通报(2019年18期)2019-10-16 06:21:32
山东林业科技(2018年6期)2019-01-08 09:48:04
农家科技中旬版(2018年2期)2018-05-05 10:09:54
现代农业(2018年7期)2018-02-18 10:58:09
新农村(2015年10期)2015-09-28 00:37:29
河池学院学报(2014年5期)2014-02-27 13:37:18
