
甲氧虫酰肼对舞毒蛾幼虫多功能氧化酶和羧基酰胺酶活性的影响1)
2013-03-05 09:58曹传旺严善春
东北林业大学学报 2013年5期
钟 华 曹传旺 薛 煜 严善春
(东北林业大学,哈尔滨,150040)
舞毒蛾(Lymantria dispar)属鳞翅目(Lepidoptera)毒蛾科(Lymantridae),是一种食性广谱的食叶害虫,能取食500 多种植物,主要为害杨、柳、栎、榆、桑、桦、油杉、沙棘、落叶松、苹果、梨、杏、桃、柑桔、山楂等树木[1-3],给农林业经济造成严重的损失。甲氧虫酰肼属于酰肼类拟蜕皮激素类杀虫剂,能模拟20 -羟基蜕皮激素与目标害虫蜕皮激素受体复合物相互作用,以蜕皮激素的作用方式导致昆虫致死性蜕皮[4]。该药剂对鳞翅目昆虫具有高度的选择毒性,对非靶标生物以及环境安全,已经成为防治蔬菜、棉花、玉米及其他农作物上多种鳞翅目害虫的理想药剂[5]。害虫对杀虫剂的氧化代谢,在其抗药性形成中起重要作用。其中,多功能氧化酶(MFOs)能够催化各种结构不同的内源或外源化合物氧化,如:脂肪酸、甾体激素、药物、杀虫剂及各种环境有害化合物等[6]。王建军[7]等研究发现,亚致死剂量甲氧虫酰肼对斜纹夜蛾MFOs 具有一定的诱导作用。廖月枝等[8-9]研究表明甲氧虫酰肼能有效干扰舞毒蛾幼虫体内羧酸酯酶、MFOs、谷胱甘肽S -转移酶活性及不同组织中蛋白质的表达。甲氧虫酰肼分子结构中含有芳基酰胺结构,而广泛存在于昆虫体内的酰胺酶则被认为可能参与带有这类结构的某些农药的水解作用,在昆虫对具有此类结构的杀虫剂抗性中可能起着重要作用[10]。刘玉娣等[11]报道指出小菜蛾对抑太保的敏感性差异较大,并且其体内羧基酰胺酶的活力会随着温度及虫龄的变化而改变。但迄今为止,国内外有关甲氧虫酰肼对羧基酰胺酶活性的影响研究报道较少。笔者研究甲氧虫酰肼对舞毒蛾幼虫MFOs 及其酰胺酶活性的影响,为明确甲氧虫酰肼对舞毒蛾的毒理分子机制及开发新型蜕皮激素类似物提供理论依据,以便在林间合理防治舞毒蛾的危害。
1 材料与方法
供试昆虫:2010年3月在东北林业大学校园及其示范林场内采集舞毒蛾卵块,将卵块置于人工智能培养箱内孵化,用人工饲料饲养幼虫,每天观察幼虫生长发育和蜕皮情况,3 ~4 d 更换一次饲料,及时清理幼虫的排泄物和食物残渣。恒温饲养条件为温度(25 ±1)℃,相对湿度60% ~80%,光周期16 h 光照、8 h 黑暗。
供试药剂:98%甲氧虫酰肼原药,美国陶氏益农公司产品,配置为100 mol·L-1的母液。乙二胺四乙酸、牛血清白蛋白、考马斯亮蓝G -250、氢氧化钡、硫酸锌,国药集团化学试剂有限公司;对硝基苯甲醚、苯甲基磺酰氟、二硫苏糖醇、硫脲、对硝基乙酰苯胺,美国Sigma 公司;还原型辅酶Ⅱ四钠,瑞士Rocha 公司;磷酸氢二钠和磷酸二氢钠及其他试剂均为分析纯。
毒力测定:采用叶片药膜法[12-13]。甲氧虫酰肼用丙酮液(V(丙酮)∶ V(蒸馏水)=2∶ 1)作为溶剂配成100 mol·L-1母液,之后用蒸馏水稀释成0.1、1.0、1.5、2.0、2.5、5.0、10.0、50.0 mol·L-1,每个梯度中丙酮溶剂含量均小于1%,以蒸馏水处理作为对照。将未接触过任何药剂的白桦叶片浸入稀释好的药液中,10 s 后取出,置于阴凉处晾干。用蘸有充足水分的脱脂棉裹住叶柄,放入透气性良好的塑料培养皿(直径9 cm)中。将健康、大小一致,且饥饿12 h 的2、3 龄舞毒蛾幼虫接到白桦叶片上,每龄每皿放入10 头,每龄每个质量浓度重复3 次。置于人工智能培养箱内饲养,48 h 后检查死亡数,以毛笔轻触幼虫不能活动视为死亡。
亚致死浓度处理:根据毒力测定结果,选用甲氧虫酰肼48 h 的LC5和LC20剂量处理白桦叶片,饲喂健康、大小一致,且饥饿12 h 的2、3 龄舞毒蛾幼虫,每龄每皿放入10 头,处理方法和饲养条件同毒力测定,以蒸馏水处理作对照。处理6、12、24、48、72 h后,分别挑选活泼的2、3龄舞毒蛾幼虫,于冰箱中-80 ℃储存,用于酶活性测定。
酶活性测定:在用甲氧虫酰肼处理后不同时间,每一处理每一重复取5 ~10 头幼虫,整体匀浆,测定不同处理时间MFOs 和羧基酰胺酶的活性。MFOs活性测定参考Hansen 等的方法[14]略作修改。取4×10-3mol·L-1对硝基苯甲醚1.0 mL,0.1 mol·L-1pH 值7.8 的磷酸缓冲液0.8 mL,5 ×10-4mol·L-1NADPH-Na40.2 mL 和酶液1.0 mL,以等体积的0.1 mol·L-1pH 值7.8 的磷酸缓冲液作对照,摇匀置于37 ℃水浴中反应30 min,然后在反应体系中加入1.0 mol·L-1盐酸1.0 mL 终止反应,再加入5.0 mL 氯仿萃取,静止10 min 后,在氯仿层移取3.0 mL 到另一组试管内,再加入0.5 mol·L-1氢氧化钠3.0 mL溶液萃取,静止10 min,取水相2.0 mL 于比色皿中,于400 nm 处测定OD 值。酶活力单位以mol·g-1·min-1表示。羧基酰胺酶活性测定参考Erlenger等的方法[15]略作修改。5 mL 反应体系中含有2 mL酶液(1 mg 蛋白),0.1 mol·L-1pH 值7.5 的磷酸缓冲液2.9 mL,0.1 mL 5 g·L-1对硝基乙酰苯胺。用煮死的酶液作为对照。反应混合物于30 ℃恒温水浴锅中震荡温育30 min,将反应管于沸水中放置10 min 以终止反应,然后向所有反应管中加入1 mL 0.15 mol·L-1氢氧化钡和0.15 mol·L-1硫酸锌去除蛋白;于0 ℃、10 000 r/min 下离心15 min,吸取上清液在405 nm 处测定OD 值。酶活力单位以mol·g-1·min-1表示。酶的比活力(U/mg)=酶活力/蛋白质浓度。
蛋白质质量浓度测定:采用Bradford 考马斯亮蓝G-250 法[16]。
数据统计与分析:利用POLO 软件处理分析毒力测定结果,计算致死中浓度(LC50)和亚致死浓度(LC5和LC20)及其95%置信区间;采用SPSS17.0软件进行方差分析,差异显著性检验采用LSD 法,显著水平p=0.05 或p=0.01。
2 结果与分析
2.1 甲氧虫酰肼对舞毒蛾幼虫的毒力
甲氧虫酰肼对舞毒蛾幼虫致死中浓度及其95%置信区间见表1。甲氧虫酰肼对舞毒蛾2、3 龄幼虫均表现出较高的活性,2 龄幼虫对药剂较为敏感,48 h LC50为14.004 mol·L-1,而3 龄幼虫48 h LC50则为24.105 mol·L-1。随着龄期的增长,所需药量增加,但是随着处理时间的延长,LC50逐渐降低,96 h 时,2、3 龄幼虫LC50分别降至2.150、2.835 mol·L-1。
2.2 甲氧虫酰肼对舞毒蛾幼虫MFOs 活性的影响
甲氧虫酰肼对舞毒蛾2 龄幼虫MFOs 活性的影响见表2。72 h 内,LC5与LC20剂量下的甲氧虫酰肼对舞毒蛾2 龄幼虫体内MFOs 活性都具有先激活再抑制再激活的作用。处理6 h 时,处理组MFOs 活性均显著高于对照组,分别为对照组的1.867、2.000倍(p <0.01);在处理12、24 h 时,MFOs 活性被抑制,其中LC5剂量下24 h 抑制作用最强,仅为对照的0.452 倍,与对照差异显著(p <0.05);LC20剂量下12 h 抑制作用达到最大,为对照的0.500 倍,与对照差异显著(p <0.05)。从处理48 h 开始,甲氧虫酰肼对MFOs 活性具有明显的激活作用,且均在处理72 h 时,LC5与LC20剂量对MFOs 的激活作用最强,分别为对照的6.143、4.571 倍,随着时间的延长,药剂对该酶激活作用呈上升趋势。

表1 甲氧虫酰肼对舞毒蛾幼虫的毒力 mol·L -1

表2 亚致死剂量甲氧虫酰肼对舞毒蛾幼虫MFOs 活性的影响
由表2可以看出,甲氧虫酰肼LC5和LC20对舞毒蛾3 龄幼虫体内MFOs 活性的影响与对2 龄幼虫该酶活性的影响具有相反的趋势,即对MFOs 活性具有先抑制再激活再抑制的作用。处理6 h 时,MFOs 活性被抑制;处理12 h 时,MFOs 活性有所上升;处理后24 ~72 h,MFOs 活性再次被抑制,其中24 h 抑制作用最强,LC5和LC20处理下MFOs 活性分别为对照的0.160、0.115 倍,均与对照差异极显著(p <0.01)。
2.3 甲氧虫酰肼对舞毒蛾幼虫羧基酰胺酶活性的影响
从表3可以看出,用甲氧虫酰肼48 h LC5和LC202 种剂量处理后,舞毒蛾2、3 龄幼虫体内羧基酰胺酶呈激活—抑制—激活—抑制的曲线波动:处理6 h 时,羧基酰胺酶的活性明显被激活;至12 h 时,羧基酰胺酶活性随之被抑制;处理后24 ~48 h,该酶活性被再次激活,其中24 h 2 种剂量激活作用均达到最强。24 h 时,LC5剂量对舞毒蛾2、3 龄幼虫激活作用分别为对照的1.409、1.148 倍,LC20剂量分别为对照的1.264、1.148 倍,均与对照差异极显著(p <0.01)。随着处理时间的延长,处理72 h 时,2种剂量对羧基酰胺酶的活性均表现出很强的抑制作用。72 h 时,LC5剂量下2、3 龄幼虫的羧基酰胺酶活性分别为对照的0.668、0.729 倍,LC20剂量分别为对照的0.724、0.729 倍,均与对照差异极显著(p <0.01)。
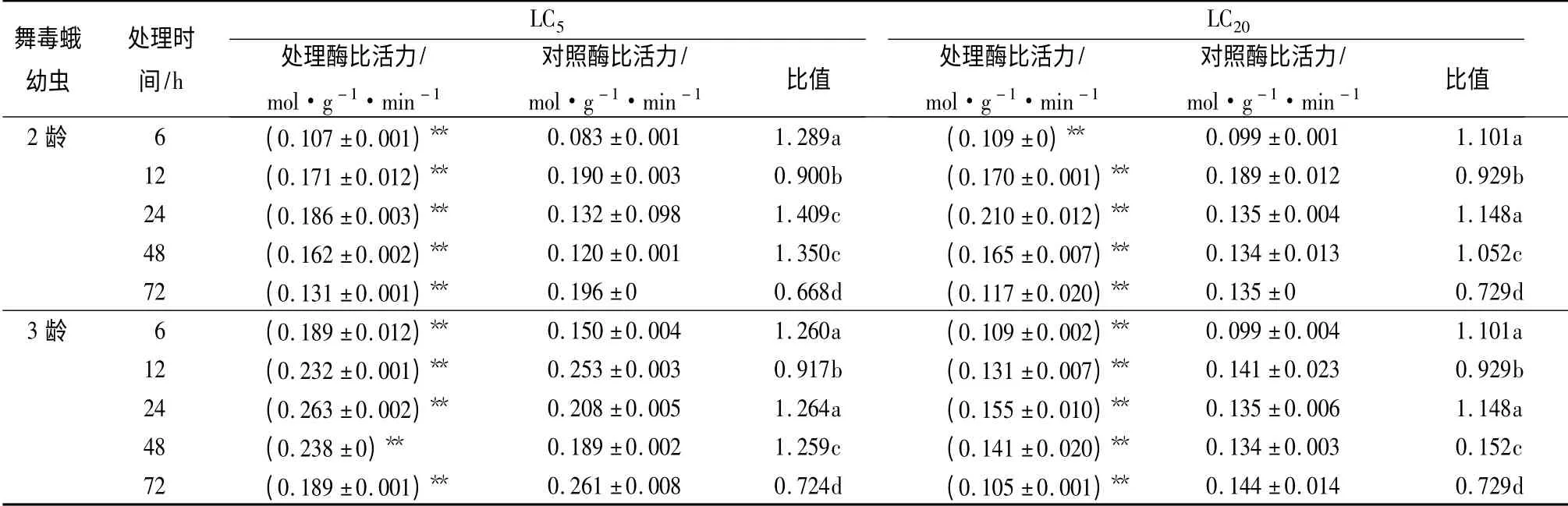
表3 亚致死剂量甲氧虫酰肼对舞毒蛾幼虫羧基酰胺酶活性的影响
3 结论与讨论
多功能氧化酶是昆虫体内重要的解毒酶,在杀虫剂代谢、昆虫抗药性等方面都具有重要作用[17-18]。王建军等[7]在室内研究了甲氧虫酰肼对斜纹夜蛾亚致死效应,结果表明甲氧虫酰肼对MFOs 具有一定的诱导作用。廖月枝等[8]报道甲氧虫酰肼对舞毒蛾4、6 龄幼虫MFOs 表现为先抑制后诱导作用。本研究发现,甲氧虫酰肼LC5和LC20对舞毒蛾2 龄幼虫MFOs 活性均表现出先激活再抑制再激活效应,处理初期MFOs 活性被诱导激活,增强水解代谢达到解毒的目的;随着处理时间的延长,当药剂在幼虫体内含量增加后,该酶活性逐渐被抑制,降低了幼虫对该药剂的解毒作用。2 种剂量对舞毒蛾3 龄幼虫MFOs 的影响与2 龄幼虫具有相反的趋势,这与鄢杰明等[9]研究发现LC50剂量甲氧虫酰肼对舞毒蛾3 龄幼虫体内的MFOs 表现为先抑制后激活的作用具有一致性。这些变化都表明甲氧虫酰肼对舞毒蛾2 龄和3 龄幼虫体内MFOs 活性具有一定的干扰作用。昆虫对具有酰胺键的某些有机磷化合物(如乐果)的抗性是由于对酰胺酶的诱导形成。Metcalf 等[19]报道盐泽灯蛾幼虫及蚊幼虫对灭幼脲的降解,发生于灭幼脲分子中的酰胺键位点。吴刚等[20]研究证实小菜蛾体内酰胺酶对抑太保的降解作用,可能是导致其对抑太保产生抗性的重要机制。根据笔者的试验结果,甲氧虫酰肼对舞毒蛾2 龄和3 龄幼虫体内羧基酰胺酶影响存在明显的时间效应,具有类似的变化趋势,即表现出激活—抑制—激活—抑制的趋势,2 种剂量在24 h 对羧基酰胺酶的激活作用均达到最强,在72 h 对羧基酰胺酶的抑制作用达到最强。说明甲氧虫酰肼能有效干扰舞毒蛾2 龄和3 龄幼虫体内羧基酰胺酶的活性,从而干扰羧基酰胺酶对甲氧虫酰肼的降解作用。综上所述,甲氧虫酰肼能有效干扰舞毒蛾低龄幼虫体内MFOs和羧基酰胺酶的活性,干扰对甲氧虫酰肼的降解作用,有效防治舞毒蛾。
[1] 李晓芬.舞毒蛾基本知识及其防治[J]. 现代园艺,2012(14):155.
[2] 胡春祥.舞毒蛾生物防治研究进展[J]. 东北林业大学学报,2002,30(4):40 -43.
[3] 王敏,李相华,韩云. 舞毒蛾的防治技术[J]. 绿色科技,2010(11):59 -60.
[4] Carlson G R,Dhadialla T S,Hunter R,et al. The chemical and biological properties of methoxyfenozide,a new insecticidal ecdysteroid agonist[J]. Pest Management Science,2001,57:115 -119.
[5] 朱丽梅.一个新的蜕皮激素拮抗剂:甲氧酰肼[J]. 世界农药,2001,23(6):50 -52.
[6] 邱立红,张文吉.多功能氧化酶系(MFO)与棉铃虫抗药性关系初步研究[J].农药学学报,1999,1(2):54 -60.
[7] 王建军,田大军.甲氧虫酰肼对斜纹夜蛾亚致死效应研究[J].棉花学报,2009,21(3):212 -217.
[8] 廖月枝,严善春,曹传旺,等.甲氧虫酰肼对舞毒蛾幼虫解毒酶及其体内蛋白质表达的影响[J].林业科学,2012,48(8):99 -105.
[9] 鄢杰明,廖月枝,严善春,等.甲氧虫酰肼对舞毒蛾解毒酶和保护酶活性的影响[J].东北林业大学学报,2010,38(11):112 -114.
[10] 张宗炳.昆虫毒理学的新进展[M]. 北京:北京大学出版社,1982.
[11] 刘玉娣,赵士熙,吴刚.小菜蛾对抑太保的敏感性及其酰胺酶的活力[J]. 福建农林大学学报:自然科学版,2002,31(2):181 -184.
[12] 马惠,王开运,夏晓明,等.5 种新型杀虫剂对甜菜夜蛾幼虫的毒力测定[J].现代农药,2006,5(3):44 -46.
[13] Zhao J Z,Li Y X,Collins H L,et al. Monitoring and characterization of diamondback moth (Lepidoptera:Plutellidae)resistance to spinosad[J]. Journal of Economic Entomology,2002,95(2):430 -436.
[14] Hansen L G,Hodgson E. Biochemical characteristics of insect microsomes:N-and O-demethylation[J]. Biochemical Pharmacology,1971,20(7):1569 -1578.
[15] Erlanger B F,Kokowsky N,Cohen W. The preparation and properties of two new chromogenic substrates of trypsin[J]. Archives Biochemistry Biophysics,1961,95(2):271 -278.
[16] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein unitizing the principle of protein-dye binding[J]. Analytical Biochemistry,1976,72(1/2):248 -254.
[17] 吴青君,张文吉,张友军,等. 解毒酶系在小菜蛾对阿维菌素抗性中的作用[J].农药学学报,2001,3(3):23 -28.
[18] Mohan M,Gujar G T. Local variation in susceptibility of the diamondback moth,Plutella xylostella (Linnaeus)to insecticides and role of detoxification enzymes[J]. Crop Protection,2003,22(3):495 -504.
[19] Metcalf R L,Lu Po yung,Bowlus S. Degradation and environmental fate of 1-(2,6-difluoro benzoyl)-3-(4-chlorophenol)urea[J]. Agric Food Chem,1975,23(3):359 -364.
[20] 吴刚,宫田正.酰胺酶在小菜蛾对抑太保抗性中的作用[J].福建农业大学学报,1998,27(1):92 -95.
猜你喜欢
四川蚕业(2021年4期)2021-03-08
四川蚕业(2021年3期)2021-02-12
纺织科学与工程学报(2020年1期)2020-06-12
世界农药(2019年3期)2019-09-10
世界农药(2019年3期)2019-09-10
林业与生态(2016年8期)2016-12-14
现代农业(2016年6期)2016-02-28
应用化工(2014年10期)2014-08-16
应用化工(2014年7期)2014-08-09
