
园林绿化华木莲的生长动态及环境影响研究
2013-02-22 07:39施建敏裘利洪俞志雄
江西农业大学学报 2013年3期
施建敏,裘利洪,俞志雄
(江西农业大学 林学院,江西 南昌 330045)
华木莲(Sinomanglietia glauca Z.X.Yu et Q.Y.Zheng),又名:落叶木莲(Manglietia decidua Q.Y.Zheng)是1994 年发表的木兰科新物种[1],为国家I 级重点保护植物、中国古特有植物,它对于研究木兰科分类系统及演化具有重要价值[2]。目前,华木莲的野生种群仅孤岛状分布于江西省宜春市和湖南省永顺县两地[3,4]。
华木莲除具有重要科学研究价值外,其树干挺直、树形端庄、枝叶扶疏、花朵芳香馥郁,是园林绿化的优良树种,具有较高的推广应用价值[3-5]。然而,研究表明华木莲对环境条件的要求严格[3,6],笔者研究发现华木莲作绿化树种应用时,只在公园、小区等较向阳、凉爽、湿润的生境成活率较高,而在广场、街道等光照强、温度高的场所不能成活(另文发表),这也是濒危植物生态适应能力较弱的共同特点[7]。
光照、水分、温度和土壤养分等是制约树木生长的主要环境因子,研究并掌握这些因子与树木生长之间的内在联系是林木培育和管理的关键[8-11]。俞志雄等[12]通过树干解析研究了野生华木莲的生长过程,龚德海等[5]对赣北引种的华木莲进行了生长观测,但是环境对华木莲生长的影响尚无报道。华木莲是濒危树种,其生态适应能力稍弱,当用作园林绿化时,城市环境比自然环境要严酷。虽然华木莲在公园、小区等绿化生境栽植能成活,但是其长势直接影响景观美化效果。因此,掌握华木莲生长与环境的关系是绿化应用的关键,它可为华木莲的绿化栽植和养护提供理论指导。
1 材料与方法
1.1 材料
研究对象为植于南昌市的华木莲绿化苗,小区生境13 株,公园生境28 株。小区生境:居住区路缘草坪;公园生境:乔木群落林缘,乔木层郁闭度为0.75,灌木层和草本层稀疏。两种生境的华木莲均为2006 年栽植,移栽时大小均匀整齐,平均胸径3.8 cm、株高4.2 m;土壤类型均为红壤。
1.2 方法
1.2.1 生长观测 胸径大小与树木的生长具有密切关系,胸径常作为树木生长的表征指标[13]。本试验对华木莲进行每株编号,从2011 年3 月至2012 年3 月每季度测量1 次胸径,具体为2011 年3 月、6月、9 月、12 月和 2012 年 3 月的月初。
1.2.2 生境因子测定 从2011 年3 月至2012 年3 月,采用英国 Delta-T 公司的 WET -2 测定仪每月测量华木莲的土壤含水率(Water Content,WC)、土壤温度(Soil Temperature,ST)和土壤电导率(Electrical Conductivity,EC),每株进行3 点测量,取平均值。每月选典型天气用TES—1332A 数字照度计测量光照强度,以各植株的实际光照与全光照比值表征植株全年的光照状况。每植株进行3 重复随机采集土壤样品,取样深度为0 ~30 cm。土壤样品风干后分别过2 mm 和100 目筛待分析,养分的测定为3 重复取平均值。土壤理化特性分析按照《土壤农业化学分析方法》的方法进行测定[14],测定指标为有机质(OM)、速效氮(AN)。
1.2.3 数据分析 本研究采用2011 年至2012 年观测值的平均数和变异系数(CV)来表征华木莲的年度土壤水分、温度和电导率的资源水平和异质性[15]。运用SPSS 15.0 进行试验数据的显著性检验、方差分析、相关分析和逐步回归分析。如果自变量与因变量呈明显二次曲线关系,则用二次多项式回归进行拟合,逐步回归分析时把二次多项式的变量转换成线性再纳入回归方程分析[16]。使用AMOS 7.0 构建通径分析模型,模型适合度的衡量指标为x2、RMSEA 和GFI。所有变量在分析前均检查正态性,如有不符则进行对数转换。
2 结果与分析
2.1 胸径年生长动态
从2011 年3 月至2012 年3 月,华木莲胸径年平均生长量为(0.52 ±0.35)cm;其中小区生境的华木莲平均胸径从(7.11 ±1.28)cm 生长到(7.93 ±1.62)cm,年平均生长量(0.82 ±0.39)cm;公园生境的华木莲平均胸径从(4.50 ±0.93)cm 生长到(4.87 ±1.12)cm,年平均生长量(0.37 ±0.23)cm(图1A);小区生境的华木莲胸径年生长量是公园生境的2.2 倍,两者差异达极显著水平(P =0.002)。总体来看,华木莲胸径年生长量(Annual Growth of DBH,AGDBH)与植株观测时初始胸径大小呈极显著正相关,相关系数为0.863(P=0.000),小区生境和公园生境的相关系数分别为0.798 和0.791(图1B)。
从季节生长来看,华木莲在春季、夏季生长旺盛,而秋季、冬季基本停止生长;但不同生境类型的生长量有差异,小区生境华木莲每个季节的胸径生长量均大于公园生境(图1C)。而且,虽然两种生境华木莲的夏季胸径生长量与春季相当,但是夏季胸径生长分化大,夏季小径植株基本停止生长,大径植株的生长仍旺盛(图1C、1D)。

图1 华木莲胸径年生长动态及其与胸径关系Fig.1 Gowth dynamic of DBH and its correlation with DBH of S.glauca planted in residential district and park
2.2 胸径年生长量与环境因子关系
对实验观测年度内华木莲的胸径年生长量(AGDBH)和光照状况(LC)、土壤含水率(WC)、土壤温度(ST)、电导率(EC)的资源水平与异质性进行分析可知(表1),土壤含水率、土壤温度、电导率、光照状况和有机质含量对胸径年生长量均具有显著影响,其中土壤温度、光照状况与胸径年生长量呈明显二次曲线关系(图2c,2g),而水分、土壤温度、电导率的异质性和速效氮含量则没有显著影响。将土壤温度和光照状况转化为线性与其他因子进行逐步回归分析,结果为:AGDBH= -1.490LC +0.006EC-0.366(R2=0.538,F =22.101,P =0.000),光照状况和电导率是华木莲胸径年生长量的决定因子。
然而,影响华木莲胸径年生长量的环境因子因生境的不同而有差异(图2a-g):小区生境的土壤含水率对胸径年生长量具有显著正影响,土壤温度和光照状况具有显著负影响;而公园生境的土壤含水率和土壤温度异质性对华木莲胸径年生长量具有显著负影响,水分异质性(CV of WC)、土壤温度、光照状况则具有显著正影响;甚至同一因子在两种生境的作用相反,如土壤含水率、土壤温度和光照状况(图2a,2c,2g)。土壤电导率、电导率异质性(CV of EC)和有机质、速效氮对两种生境胸径年生长量均无显著影响(图2e,2f,2h,2i)。逐步回归结果表明,小区生境只有光照状况是胸径年生长的决定因子:AGDBH= -2.445LC+2.992(R2=0.723,F =28.700,P =0.000),含水率和土壤温度虽然对胸径年生长有显著影响,但因其与光照状况密切相关(rWC·LC= -0.766,P=0.002;rST·L=0.933,P =0.000)而被排除出方程;公园生境胸径年生长主要受光照状况和土壤温度共同影响,AGDBH =0.590LC +0.129ST -1.978(R2=0.488,F =11.921,P =0.000),水分异质性、土壤温度异质性和光照状况、土壤温度具有显著的多重共线性而未进入方程(rWC·LC= - 0.436,P = 0.020;rCVofWC·LC= 0.445,P = 0.018;rCVofST·LC= -0.585,P =0.001;rCVofWC·S=0.398,P =0.036;rCVofST·ST= -0.978,P =0.000)。
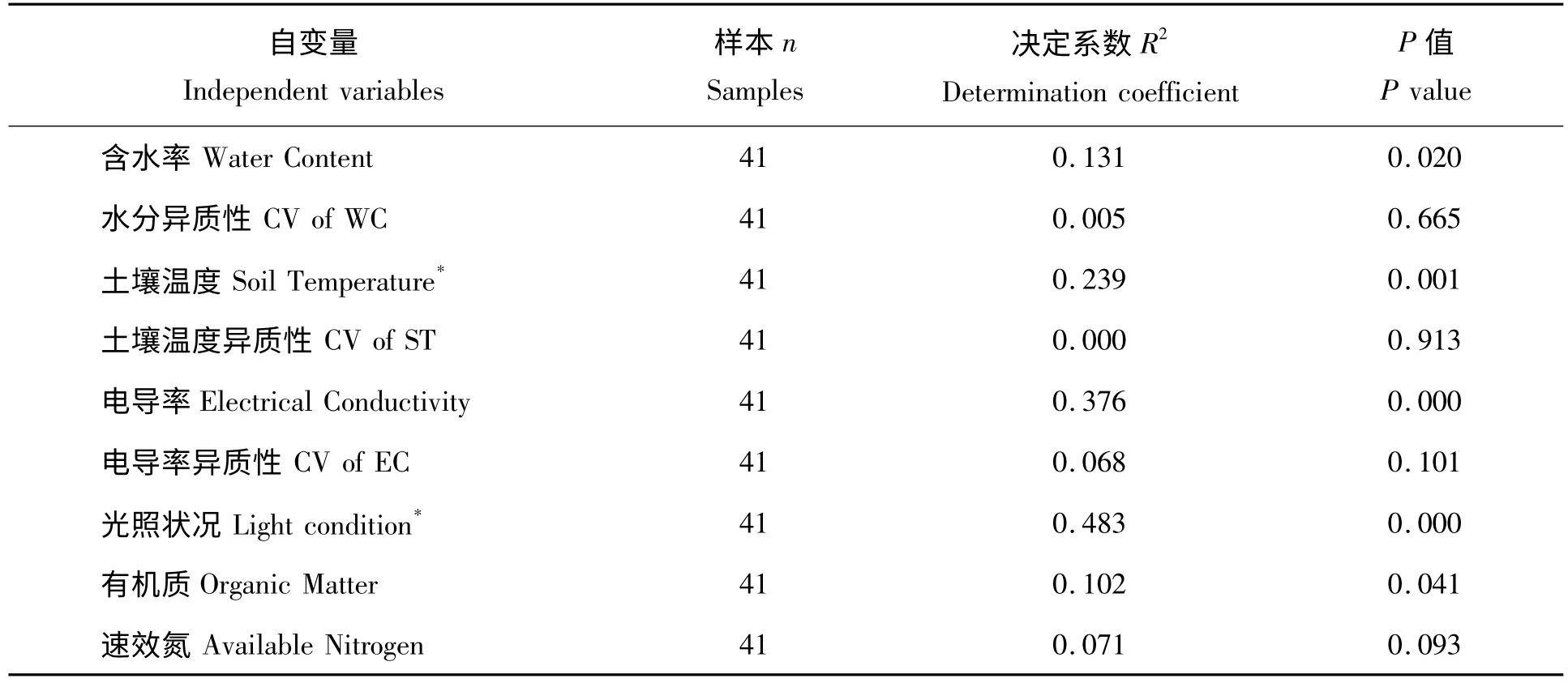
表1 华木莲胸径年生长量与环境因子的回归分析Tab.1 Regressions of environmental factors on annual growth of DBH of S.glauca
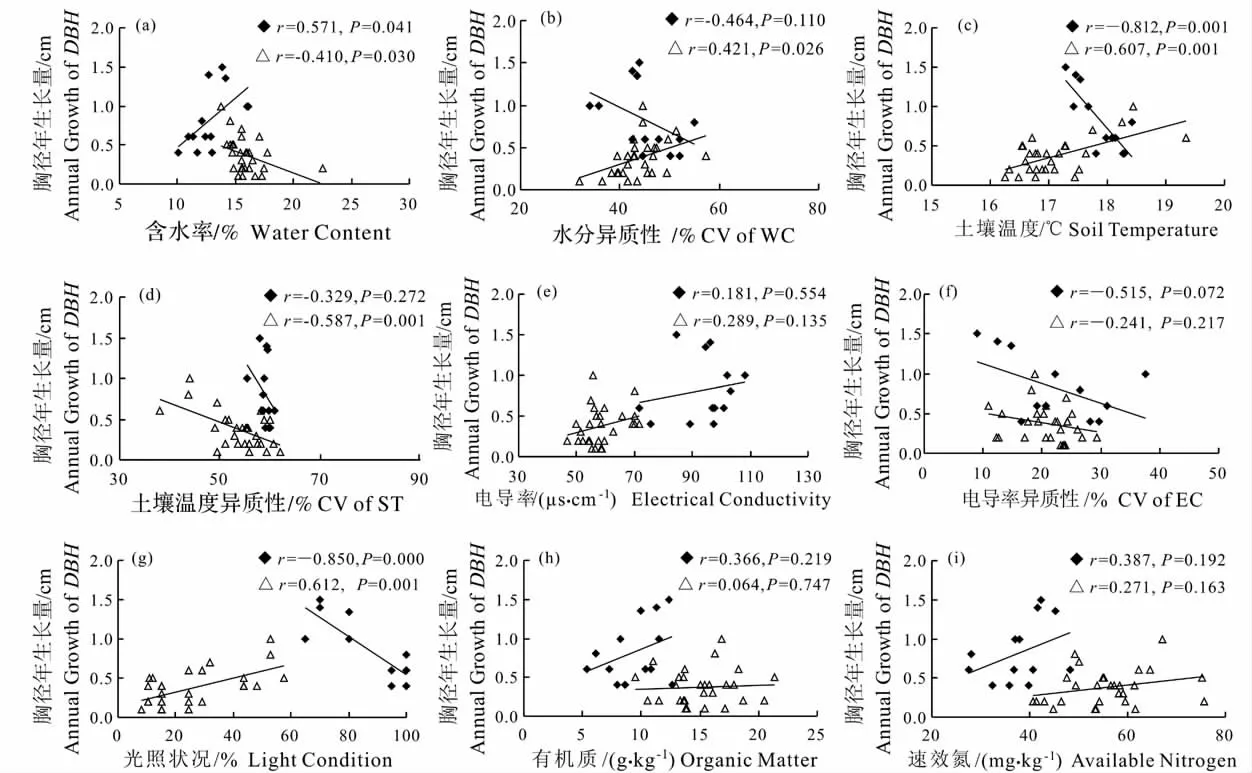
图2 华木莲胸径年生长量与环境因子的关系Fig.2 Relationship between AGDBH and environmental factors of S.glauca
2.3 胸径生长与生境因子关系
为考察华木莲胸径生长与环境的关系,将2012 年3 月的胸径与环境因子作分析,结果表明:华木莲胸径大小不受资源异质性的显著影响,而与光照、水分、温度和土壤养分的高低有显著相关关系(表2),而且土壤温度、光照状况与胸径呈明显二次曲线关系(图3c,3g)。将土壤温度和光照状况转化为线性与其他因子一起进行逐步回归,结果为:DBH= -4.652LC+0.039EC+1.067(R2=0.678,F=39.929,P=0.000),华木莲的胸径生长主要取决于光照状况和电导率。

表2 华木莲胸径生长与环境因子的回归分析Tab.2 Regressions of environmental factors on DBH of S.glauca

图3 华木莲胸径生长与环境因子的关系Fig.3 Relationship between DBH and environmental factors of S.glauca
同样,影响华木莲胸径生长的环境因子因不同生境而有较大差异(图3):小区生境的华木莲胸径随光照状况和土壤温度的降低而显著增大,其他因子则无显著影响;公园生境的华木莲胸径受多因子的影响,除与有机质含量、电导率异质性没有显著相关外,与其他环境因子均显著相关。逐步回归分析的结果为:在小区生境,光照状况和水分异质性是胸径的决定因子,DBH= -13.765LC +17.552CV of WC +12.216(R2=0.649,F=9.243,P=0.005);水分异质性虽与胸径不显著相关却进入了回归方程,而土壤温度与胸径显著相关却被排除出方程;通过构建通径分析模型(x2=0.497;RMSEA=0.000;GFI =0.979)发现,土壤温度主要是通过影响水分异质性而间接对胸径产生作用,作用系数为0.509;光照状况对胸径具有直接和间接作用(系数分别为-1.169 和0.505),总作用系数为-0.664;水分异质性对胸径仅具直接作用(作用系数0.667)(图4)。在公园生境,虽然显著影响胸径的因子很多,但是因其与光照状况存在多重共线性(rWC·LC= -0.436,P =0.020;rCVofWC·LC=0.445,P =0.018;rST·L=0.523,P =0.004;rCVofST·LC= -0.585,P =0.001;rEC·LC=0.377,P =0.048),因此光照状况成为决定胸径生长的因子,DBH=5.153LC+3.567(R2=0.504,F=26.427,P=0.000)。
3 结论与讨论
3.1 不同生境的华木莲生长及其影响因素
小区生境和公园生境的华木莲为同年栽植的同规格苗,然而到2012 年3 月两种生境的平均胸径分别为 7.93 ±1.62 cm 和 4.87 ±1.12 cm(图1a),前者是后者的1.63 倍,达极显著水平(P =0.000)。逐步回归分析结果表明,影响小区生境华木莲胸径生长的主要因素为光照状况和水分异质性,而影响公园生境华木莲胸径生长的主要因素为光照状况。虽然光照状况都是影响两类生境华木莲生长的重要因子,但是小区生境为开敞空间,而公园生境为乔木群落林缘,两者立地条件不同,在光照状况、土壤温度、含水率和电导率等方面均有明显差别(图3),由此导致了光照状况、土壤含水率、水分异质性和土壤温度对不同生境胸径生长的影响方向完全相反(图3a -c、g)。因此,环境因子的差异是造成不同生境华木莲胸径大小有显著差别的主要原因[8-11]。
此外,同一生境的个体生长亦有分化,小区生境胸径最大10.8 cm,最小5.1 cm;公园生境胸径最大7.6 cm,最小3.2 cm(图1a)。郭起荣等[6]研究认为华木莲不耐强光和高温干旱,所以小区生境中光照过强的植株生长受到影响,而公园生境光照不足的植株其生长同样也受到限制。从两类生境的光照梯度来看,华木莲大径植株主要分布在光照状况0.5 ~0.8(图3g),因此造成生境内部植株生长分化的主要原因是生境内光照的差异,这与其他研究得出微生境影响植株生长的结论相一致[17-18]。
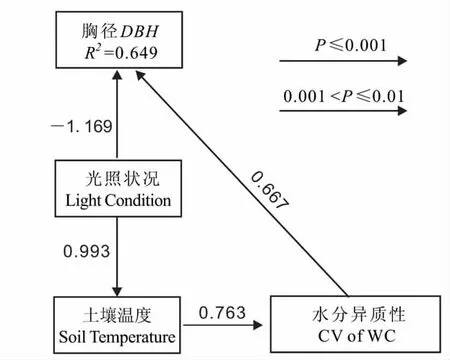
图4 小区生境华木莲胸径生长的通径分析Fig.4 Path analysis for DBH of S.glauca in residential district
3.2 环境影响华木莲生长的空间尺度效应
光照状况和水分异质性是决定小区生境华木莲生长的主要因子,而公园生境的胸径生长主要取决于光照状况。然而,当把环境的尺度放大,将两类生境全部植株一起分析时发现,电导率却和光照状况一起决定华木莲的生长。究其原因,小区生境和公园生境的环境因子有较大差异(图3),特别是光照状况、电导率、土壤含水率和土壤温度(图3a、c、e、g)表现得更加明显,两者互在高低两端形成了连续的环境梯度。当两类生境结合分析时,随着环境梯度的变大,环境因子与胸径的关系发生了改变,如光照、土壤温度和胸径就从线性关系转变为二次曲线关系(图3c、3g),电导率、有机质和速效氮等养分指标与小区生境或公园生境的胸径无显著相关而变成显著相关(表2,图3)。由此看来,制约华木莲生长的环境因素存在尺度效应,在不同空间尺度上影响胸径生长的环境因素是有差异的,这也验证了土壤养分与树木生长的关系与研究的空间尺度和环境梯度大小相关的论点[9]。此外,研究表明电导率是土壤养分、水分和盐分等密切相关的一个综合表征指标[19-20]。本研究中,土壤有机质、速效氮、土壤含水率、土壤温度和电导率都对华木莲的胸径生长有显著影响(表2),而电导率与上述因子均有显著相关关系(rEC·OM= -0.554,P =0.000;rEC·AN= -0.597,P =0.000;rEC·WC= - 0.566,P =0.000;rEC·ST= -0.630,P=0.000),这也就不难理解电导率和光照状况是决定华木莲生长的两个主要因子。
3.3 环境影响华木莲生长的时间尺度效应
不论是小区生境、公园生境的植株,还是全部植株,显著影响华木莲短期生长(胸径年生长量)和长期生长(胸径生长)的环境因子相同,其影响趋势也完全相同(图2、图3,表1、表2)。回归分析表明,光照状况和电导率都是决定华木莲短期生长和长期生长的两个主要控制因子,然而决定华木莲生长的因素在不同生境类型具有时间尺度差异:小区生境控制胸径年生长的是光照状况,而控制胸径生长的是光照状况和水分异质性;公园生境控制胸径年生长的是光照状况和土壤温度,而控制胸径生长的是光照状况。由以上分析可知,影响华木莲生长的环境因素具有尺度效应,而且时间尺度的影响会因空间尺度的变化而有差异。
3.4 华木莲生态适应性及其绿化应用启示
光照状况是影响华木莲生长最主要的环境因素,同时土壤理化性质(土壤养分、水分、温度、电导率等)对生长也具有重要作用。从华木莲的生长表现来看,小区生境光照过强和公园生境过荫蔽的植株长势均较弱,华木莲适宜生长于实际光照与全光照比值介于0.5 ~0.8 的生境,这与之前华木莲生态习性的研究结果相一致[3,6]。
华木莲小径植株在夏季基本上停止生长,而大径植株仍生长旺盛(图1d),原因主要为小径植株对夏季的高温强光适应能力弱,相比之下大径植株对环境适应能力强。因此,在园林上尽可能选用大径华木莲木绿化苗。影响华木莲生长的环境因素因不同生境而有差异的结果对不同类型生境的绿化栽植和管理具有明确的指导意义:在公园生境要透光,尽量满足华木莲的光照需求;而在小区生境应避免光照过强,在夏季还需浇水以降低高温的不利影响。此外,同一生境的华木莲个体间存在较大生长差异(图1a)的现象表明,华木莲的绿化栽植不但要做到适地适树,而且还要注重微生境的管理[17-18]。
致谢:林学0702 班冷丹凤、余雅婷、黄情、洪群、谢庚根、孟林林、胡亚琴同学,林学1002 班王雪岸同学参加了试验调查和测试分析工作,在此一并致谢。
[1]俞志雄.华木莲属——木兰科一新属[J].江西农业大学学报,1994,16(2):202 -204.
[2]吴征镒,孙航,周浙昆,等.中国植物区系中的特有性及其起源和分化[J].云南植物研究,2005,27(6):577 -601.
[3]俞志雄,廖军,林新春,等.华木莲植物群落的生态学研究[J].江西农业大学学报,1999,21(2):73 -77.
[4]侯伯鑫,易宏,林峰,等.湖南永顺县落叶木莲资源考察初报[J].湖南林业科技,2006,33(1):7 -10.
[5]龚德海,陈英,夏敏娟,等.华木莲在赣北的适应性初报[J].江西林业科技,2011,38(6):24 -25.
[6]郭起荣,俞志雄,施建敏.华木莲与木莲属两树种光合生理生态研究[J].江西农业大学学报,2003,25(5):645 -651.
[7]龚洵,张启泰,潘跃芝.濒危植物的区系性质与迁地保护[J].云南植物研究,2003,25(3):354 -360.
[8]郭文福.米老排人工林生长与立地的关系[J].林业科学研究,2009,22(6):835 -839.
[9]樊艳文,王襄平,曾令兵,等.北京栓皮栎林胸径——树高相关生长关系的分析[J].北京林业大学学报,2011,33(6):146 -150.
[10]Wang X P,Fang J Y,Tang Z Y,et al.Climatic control of primary forest structure and DBH - height allometry in northeast China[J].Forest Ecology and Management,2006,23(4):264 - 274.
[11]汪殿蓓,李建华,陈玉兰,等.青檀天然群落土壤成分及其对青檀生长的影响[J].生态环境学报,2010,19(10):2318-2324.
[12]俞志雄,林新春,李志强,等.华木莲生长过程的初步分析[J].江西农业大学学报,1999,21(1):95 -98.
[13]刘平,王玉涛,马履一.油松人工林单木胸径生长过程模拟及检验[J].西北林学院学报,2010,25(4):139 -143.
[14]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999.
[15]Shi J M,Ma K M,Wang J F,et al.Vascular plant species richness on wetland remnants is determined by both area and habitat heterogeneity[J].Biodiversity and conservation,2010,19(5):1279 -1295.
[16]李春喜,姜丽娜,邵云,等.生物统计学[M].3 版.北京:科学出版社,2005.
[17]杨俊,赵雨森,韩春华,等.微立地土壤物理性质差异及对脂松幼林生长的影响[J].东北林业大学学报,2008,36(6):24 -25.
[18]Titus J H,Moral R.Seedling establishment in different microsites on Mount St.Helens,Washington,USA[J].Plant Ecology,1998,134(1):13 -26.
[19]周红艺,何毓蓉,张保华,等.长江上游典型区水耕人为土的电导率与肥力评价探讨[J].西南农业学报,2003,16(1):86 -89.
[20]刘广明,杨劲松.土壤含盐量与土壤电导率及水分含量关系的试验研究[J].土壤通报,2011,32(S1):85 -87.
猜你喜欢
贵州林业科技(2022年2期)2022-06-22
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年2期)2021-07-13
水土保持研究(2019年6期)2019-10-19
新青年(2019年8期)2019-08-22
食品与健康(2018年8期)2018-08-23
中国水土保持科学(2017年3期)2017-07-05
浙江大学学报(工学版)(2016年2期)2016-06-05
创作评谭(2014年3期)2014-05-12

- 江西农业大学学报的其它文章
- 迁西板栗主栽品种授粉组合的优化