
抗条锈小麦-非洲黑麦渐渗系的分子细胞学鉴定
2012-10-26 11:08贾举庆郭红媛李倩冉董娟杨武德杨足君
山西农业大学学报(自然科学版) 2012年4期
贾举庆,郭红媛,李倩冉,董娟,杨武德,杨足君
(1.山西农业大学 农学院,山西 太谷030801;2.电子科技大学 生命科学与技术学院,四川 成都610054)
小麦是世界上重要的农作物,在我国也是仅次于水稻的主要粮食作物之一,但同其它大多数作物一样,现代农业体系下,小麦的遗传多样性丢失极为严重,其对生物性和非生物性的环境协迫变得十分脆弱,限制了小麦产量的大幅度提高和品质的进一步改良。小麦族近缘植物中蕴含着丰富的优异基因,是小麦遗传改良可利用的重要资源。目前通过染色体工程方法,已将多种小麦族近缘物种的优异基因导入小麦遗传背景中,育成了一系列小麦外源染色体附加、代换和易位系等中间材料,在生产上得到广泛的应用[1]。
黑麦属(Secale L.)作为小麦属的近缘物种,以其优良的农艺性状,如高产、广适及对病虫害的抗性,成为小麦遗传改良的优异资源,也是小麦远缘杂交中利用最成功的远缘物种之一[2]。在系统分类学上黑麦属分为栽培黑麦(S.cereale L.)和野生黑麦,其中以栽培黑麦在小麦改良中的应用最为广泛[3,4]。但近年来,由于栽培黑麦中的抗病基因对新的病理小种失去抗性、1RS/1BL易位系高产特性已得到充分发挥、一些黑麦染色体上的劣质基因被认识、发现等现象的出现,导致各国对栽培黑麦的利用停滞不前[3~5]。目前被广泛利用的小麦-黑麦易位系的供体黑麦材料,也主要局限在几个如德 国 白 粒[6]、荆 州 黑 麦[7]、秦 岭 黑 麦[8]、威 岭 黑麦[9]等少数几个黑麦品种之中。因此,充分发掘野生黑麦中的优异基因,加强野生黑麦基因资源的利用,对小麦可持续性抗病遗传育种具有重要的意义。
非洲黑麦(S.africanum Stapf)是黑麦属的重要多年生野生物种,具有矮杆、异花授粉、抗多种小麦病害等优异性状,是小麦育种潜在的可利用资源。目前关于非洲黑麦的研究报道不多,主要集中在进化方面,认为非洲黑麦和森林黑麦(S.sil vestre)遗传距离较近,与其他栽培黑麦距离较远;非洲黑麦中的异染色质比栽培黑麦少[10]。在小麦育种利用方面,四川农业大学合成了小麦-非洲黑麦的双二倍体材料,为非洲黑麦基因向小麦中导入提供了桥梁材料[11]。其他关于非洲黑麦的核型和带型、小片段渐渗系的创制与利用还未开展。
本研究拟对非洲黑麦核型、带型及非洲黑麦向小麦转育过程形成一系列的渐渗系为材料进行分子细胞学鉴定,并结合抗条锈病鉴定,筛选抗病优异材料,为利用这一优异基因资源提供理论和应用基础。
1 材料与方法
1.1 试验材料
非洲黑麦由美国密苏里植物园提供。安岳排灯麦-非洲黑麦双二倍体(AF)和波斯小麦-非洲黑麦双二倍体(BF)由四川农业大学蒋华仁研究员合成[12]。L系材料为小麦品种绵阳11与BF杂交的F6后代。安岳排灯麦、波斯小麦、Aurora(1RS/1BL品种)和中国春(CS)由电子科技大学分子细胞生物学实验室提供。
1.2 试验方法
1.2.1 C带分析
C分带参照任正隆[13]的操作流程,略加改动。
1.2.2 SCAR标记PCR扩增
PCR所用引物为非洲黑麦基因组特异引物O5 (5'-CCCAGTCACTACAACGAGAGT-3');O5 R(5'-GCTACAAGAGCTTCGTGCAG-3'),25 μL反应体系(包含50 ng基因组DNA,0.2μmol·L-1引物,200μmol·L-1d NTP,1×PCR buffer,2.0 mmol·L-1Mg Cl2和1 U Taq聚合酶);扩增条件为:94℃预变性3 min;94℃变性1 min,55℃退火1 min,72℃延伸1 min,共35个循环;72℃延伸10 min。产物由1%琼脂糖凝胶分离,紫外凝胶成像仪上观察并照相。
1.2.3 原位杂交分析
制片采用任正隆等[13]的方法,荧光原位杂交所用探针标记按DIG-Nick试剂盒(Roche公司)的操作方法,原位杂交参照Mukai的方法[14]。
1.2.4 醇溶蛋白分析
醇溶蛋白分析采用A-PAGE的方法,具体参照 Yang等的方法[15]。
1.2.5 抗条锈病分析
苗期接种与抗性鉴定于2009~2011年在山西省农业科学院作物研究所温室中进行,所用条锈菌系为国内目前流行小种CYR30和CYR31。条锈病接种采用苗期涂抹法,具体为:待感病品种SY95-71和供试小麦幼苗第一叶充分展开后,将混有滑石粉的锈菌夏孢子涂抹在已去蜡质的小麦叶片上;成株期采用喷施法接种,即将新鲜孢子悬浮液喷施于叶片上,待SY95-71充分发病时调查成株的表现型。在本试验中,按0~4级常规分级标准调查逐株记载实验材料的反应型即0、0;、1、2、3和4,其中0和0;为免疫和近免疫、1为高抗、2为中抗、3为中感、4为高感。
2 结果与分析
2.1 小麦-非洲黑麦双二倍体细胞学鉴定
以地高辛标记的非洲黑麦基因组DNA为探针,基因组原位杂交(GISH)发现,在BF双二倍体材料中鉴定出14条非洲黑麦染色体(图1 A),说明非洲黑麦染色体在双二倍体中得到很好的保存。利用染色体C带技术对同一个细胞中的染色体进行C分带,结合小麦ABD染色体的标准C带型及非洲黑麦的GISH图(图1 A和B)、参照Lukaszewski等[16]划分的栽培黑麦染色体标准C带图对这14条非洲黑麦染色体进行更明确的划分与标记,做出了非洲黑麦标准C带图(图1C)。通过将非洲黑麦与栽培黑麦的染色体C带对照比较发现,其中非洲黑麦的1Ra、2Ra、3Ra、5Ra和6Ra染色体的C带型与栽培黑麦的1R、2R、3R、5R和6R染色体的C带型相似,但4Ra和7Ra在长度和端部异染色质上都比栽培黑麦4R和7R染色体短和少。
2.2 双二倍体中染色体的重组
利用从非洲黑麦中分离出来的重复序列p Sa O5411作探针对32株BF后代材料进行荧光原位杂交(FISH)时发现,其中有9株中存在小麦染色体和非洲黑麦染色体发生小片段易位(图2 A);利用非洲黑麦基因组DNA探针对16株AF后代材料进行GISH时也发现了非洲黑麦染色体断裂及与小麦染色体发生小片段易位现象,并且这些易位中除了端部易位外还发现插入易位,在16株AF后代中发现了3株有非罗伯逊易位现象(图2B)。这些结果都说明在小麦-非洲黑麦双二倍体中小麦染色体和非洲黑麦染色体发生了大量的重组,并且重组呈现出多类型现象。
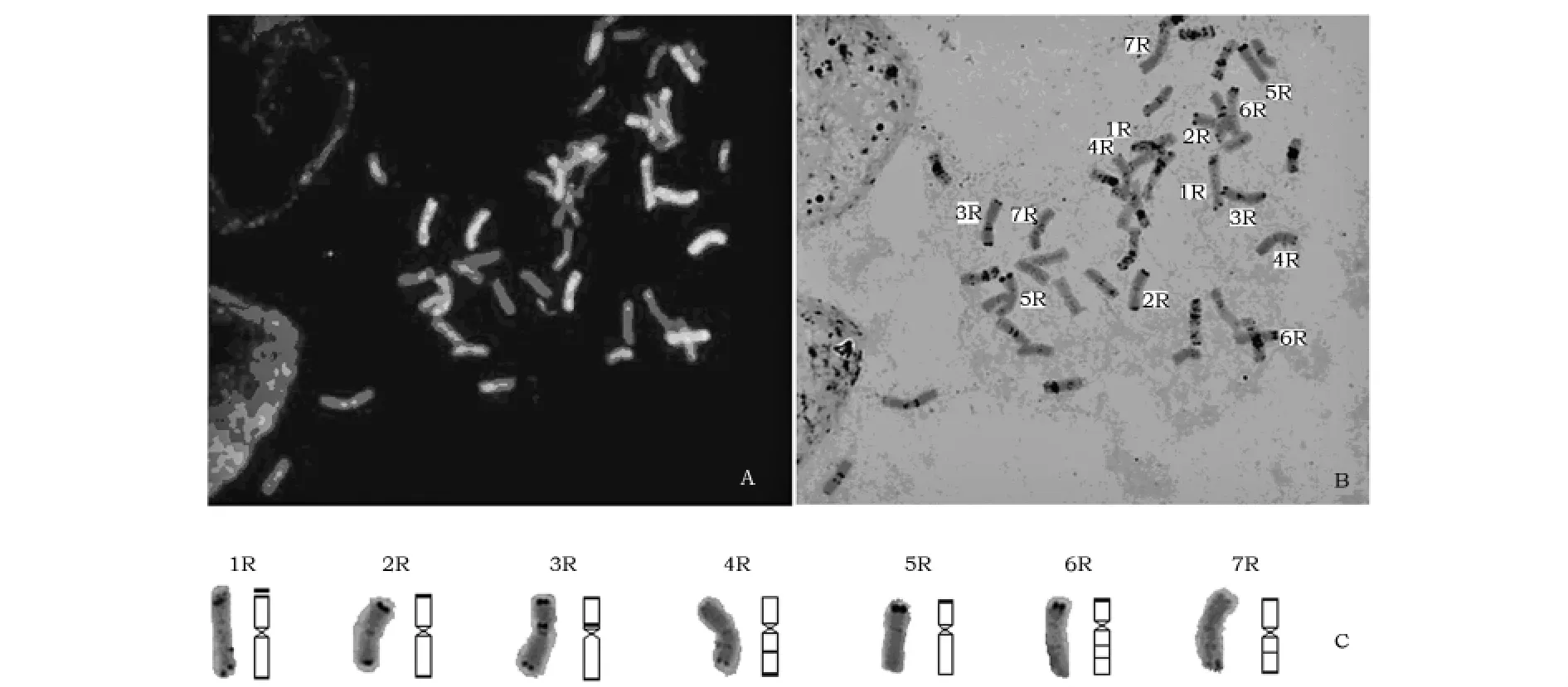
图1 非洲黑麦染色体鉴定及标准C带图的建立Fig.1 Identification of S.africaum chromoso mes and standard C band patter n
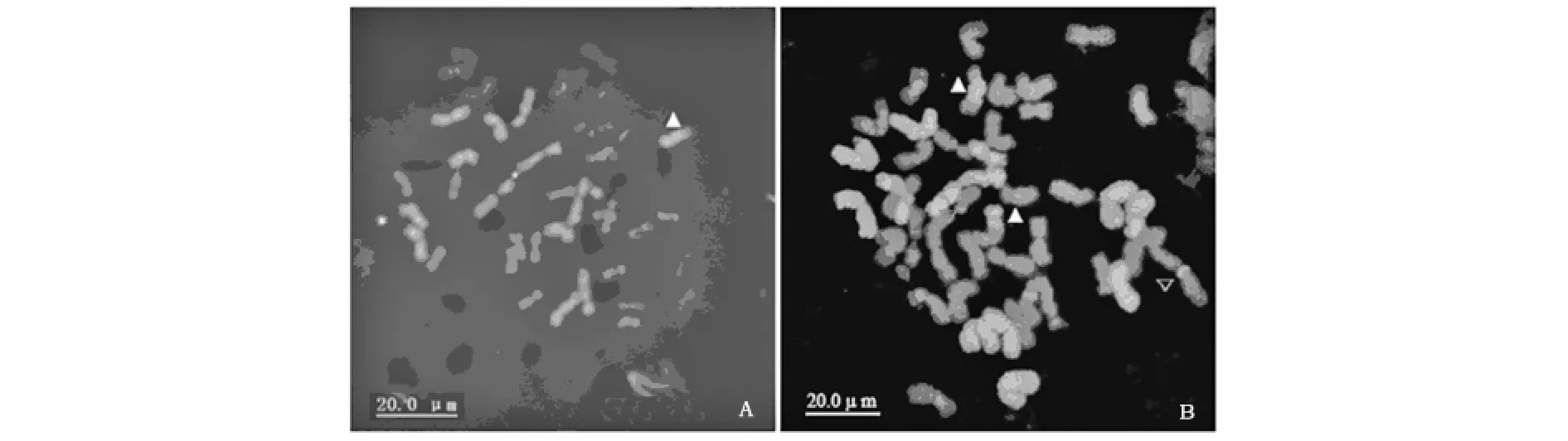
图2 有丝分裂期BF染色体FISH(A)和AF染色体GISH图(B)Fig.2 FISH(A)and GISH(B)pattern of AF mitotic metaphase chromosomes
2.3 抗条锈病渐渗系的筛选与鉴定
利用非洲黑麦基因组特异SCAR标记O5F/R对BF与MY11小麦杂交F6后103株后代材料中,检测出36株材料含有黑麦染色质(图3)。
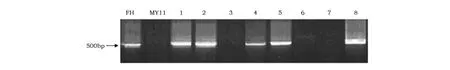
图3 部分杂交后代材料以SCAR标记PCR扩增结果Fig.3 The PCR amplification results by SCAR mar ker in partial materials
对36株含非洲黑麦染色质的材料及其亲本小麦和非洲黑麦及小麦-非洲黑麦双二倍体接条锈菌小种发现,在36株中有19株材料(L1-L19)对条锈菌表现出不同程度抗性(表1)。
综合利用生化、细胞生物的方法对这19株材料进行鉴定。利用顺序基因组原位杂交和染色体C带技术鉴定发现,L3材料中黑麦染色体插入易位到小麦染色体中,至于插入片段具体来源,由于插入片段过小,通过C带图还无法确定(图4 A和B);通过1RaS/1BL易位染色体特征GISH图,可以认为L1为只携带一条1RaS/1BL易位染色体的易位系材料(图4C和D);结合C带图,可以认为L16材料是小麦3BS染色体端部与非洲黑麦3Ra染色体发生了易位(图4E和F);利用GISH技术,认为材料L17为1Ra代换系(图4 G和H),至于代换了小麦哪条染色体,由于缺乏C带证据还无法判断;利用醇溶蛋白电泳(图4I)(箭头所示为黑麦碱特征条带)和GISH技术(图4J)分析认为,L9-15共7个材料为携带了一对1RaS/1BL易位染色体的易位系材料。其他其他携带非洲黑麦染色质的抗条锈病材料利用GISH没有检测到杂交信号。
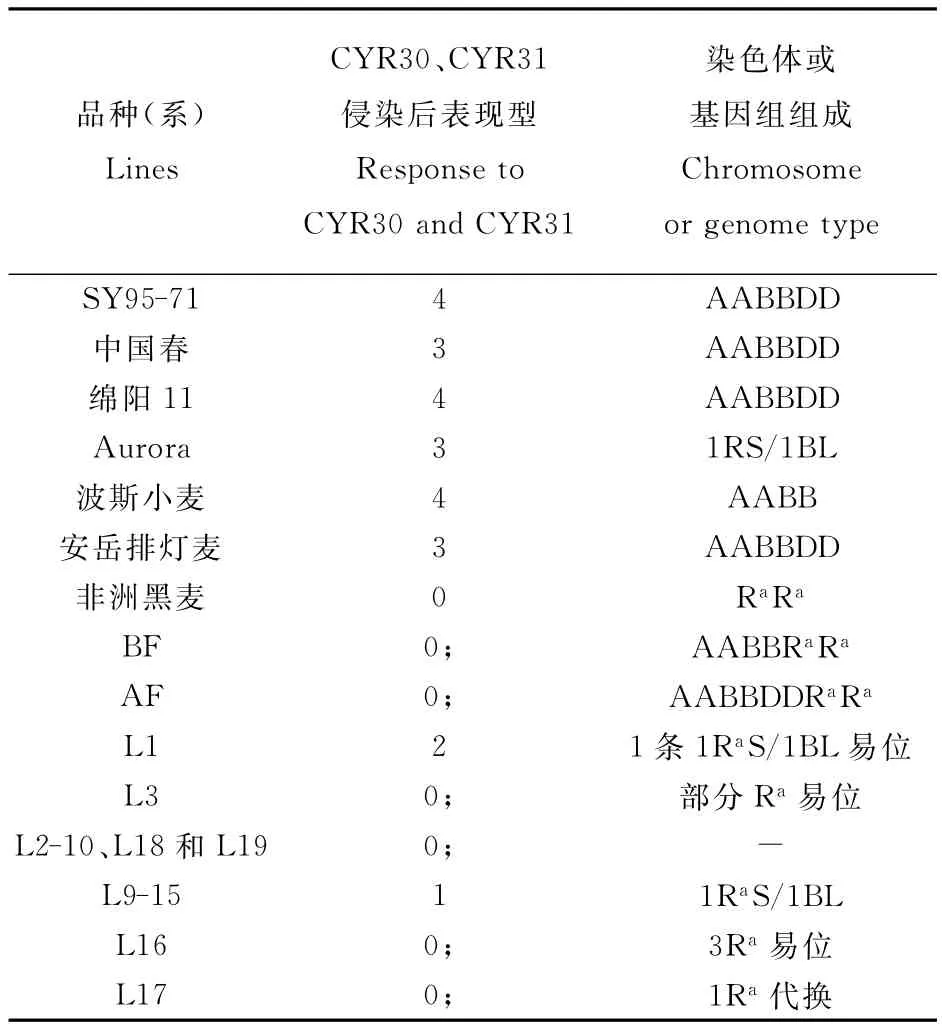
表1 试验材料的条锈病的侵染型及染色体组成Table 1 Rust response to P.striifor mis f.sp.tritici and genomic structures of parents,amphiploids,stripe rust resistant derivatives and controls

图4 部分L系材料的C带、GISH和A-PAGE图Fig.4 C-banding(A)and GISH(B)patter n and A-PAGE patter n of a part of L lines
3 结论与讨论
3.1 非洲黑麦染色体标准C带型的建立
染色体带型分析是辨别染色体的一种简单快速的方法[17]。在小麦族物种的研究中应用最广的是C带技术,但在小麦族植物远缘杂交中如要鉴定外缘染色体必需要有染色体的标准带型加以对照识别,因此Gill等用普通小麦中国春为材料,做出了小麦分带核型[18];Lukaszewski等建立了黑麦染色体C-带标准带型[16]。但研究认为非洲黑麦的异染色质跟栽培黑麦有一定的差别,这也在本研究中得以证实,如4Ra和7Ra在长度和端部异染色质上都比栽培黑麦4R和7R染色体短和少(图1)。本研究通过参考栽培黑麦染色体C带带型,建立非洲黑麦的标准C带,为今后小麦背景中快速识别非洲黑麦染色体提供依据。
3.2 双二倍体中染色体重组
远缘杂交过程中,由于染色体的不亲和往往导致出现减数分裂不稳定现象,并且容易形成非整倍体[10],这在本试验中也有所体现,在对BF后代材料中的细胞染色体计数时发现,染色体数目存在40至42条不等现象(数据未列出),这种不稳定性恰恰为育种提供了丰富的变异类型以供选择。此外,在本试验中,不管是AF还是BF中都发现了多种类型的染色体易位现象,包括小麦染色体端部与非洲黑麦染色体整条染色体或染色体片段融合、染色体的非罗伯逊易位甚至有非洲黑麦染色体片段插入到小麦染色体中的现象(图2)。而这种频繁且多类型的重组现象在栽培黑麦形成的八倍体或六倍体小黑麦中很少出现,如并没发现有栽培黑麦染色体插入易位到小麦染色体中的现象[19]。非洲黑麦与小麦染色体这种频繁的重组可能是非洲黑麦异染色质少有关。Yang等用从栽培黑麦中分离到的串联重复序列弥散状分布于栽培黑麦所有染色体上,但用该序列杂交非洲黑麦时发现,这种重复序列除了杂交非洲黑麦的6Ra染色体外,其他染色体上都没有分布[19],而研究认为,重复序列及异染色质化在植物性染色体重组中会起到抑制作用[20]。
3.3 小麦-非洲黑麦渐渗系中小片段易位现象
远缘杂交遗传改良小麦的过程中往往会产生染色体附加、染色体代换和染色体易位等现象。在本研究中,无论是AF还是BF在于MY11小麦杂交F6后代材料中我们都是发现了染色体的代换和易位(图4),但还没发现染色体的附加现象,并且代换也只是发现一对1Ra代换系材料(L17)(图4G和H),这可能与试验材料的数量不足有关。在染色体发生易位的材料中,出现了各种易位类型,如一对1RaS/1BL易位系材料(L9-15)(图4I和J);一条3Ra染色体在端部与半条小麦染色体发生易位的L16材料(图4E和F);L1和L3中的小片段易位(图4 A、B、C和D)等。有趣的是,本试验中L2-10、L18和L19等11株材料中的黑麦染色质只能通过分子标记检测(图3),而无法利用GISH技术检测(检测无杂交信号,结果没列出),这可能因为渗入到小麦中的非洲黑麦染色体易位片段过小,超出了GISH技术所能检测的最小片段范围[14]。对于这些小片段易位材料的鉴定则需要在今后试验中开发非洲黑麦的单染色体标记来加以检测。虽然这些小片段易位系难以检测,单这些小片段易位材料能很好的克服大片段易位所带来的外源不利基因,如黑麦碱(Sec1),对小麦品质的影响[21],并且小片段易位对后代材料染色体的正常配对与稳定遗传都有积极作用。因此这些材料更是今后育种中要重点利用的对象。
3.4 非洲黑麦中的抗条锈基因
黑麦中携带许多优异的抗性基因,特别是1RS染色体携带的抗条锈病基因Yr 9得到广泛的利用,在小麦遗传育种上产生了重要的影响[21]。研究发现不同黑麦属种同一染色体上携带的抗性基因也有所不同,如来源于Insave黑麦中的Pm17与Pet kus黑麦抗白粉病基因Pm8,虽然二者都是位于黑麦1RS染色体上,但其对白粉病的抗谱并不相同;同样来源于Petkus黑麦1RS染色体上的Sr 31与Imperial黑麦1RS染色体上Sr R也不尽相同;来源于不同黑麦后代L155和R12中也存在两种不同类型的1RS/1BL易位系,那些对Yr 9有毒性的条锈病生理小种在这两种黑麦及其后代材料上表现出不同的侵染类型[22]。本试验中也发现1 RS/1BL易位系材料Aur ora在对条锈病失去抗性时,L9-15和L17这些携带1RS染色体的材料却对条锈病表现出良好的抗性(表1)。因此,在传统抗条锈基因的抗病性逐渐丧失的同时,非洲黑麦1 RS染色体上仍存在优异的抗条锈新基因。此外,在本试验中还发现非洲黑麦其他染色体中也携带有优良的抗条锈基因(L16为3 Ra染色体与小麦染色体易位),这都说明非洲黑麦中携带多种抗条锈病基因,因此可以成为小麦抗条锈遗传育种的宝贵资源。
[1]张正斌.小麦遗传学[M].北京:中国农业出版社,2001:1-3.
[2]Zeller F J.1B/1 R wheat-ryechromoso me substitutions and translocations[C].In:Proceedings of the 4t h Inter national Wheat Genetics Symposium.Columbia:E.R.Sears and L.M.S.Sears.Missouri Agricultural Experiment Station,University of Missouri,1973:209-221.
[3]任正隆.黑麦种质导入小麦及其在小麦育种中的利用方式[J].中国农业科学,1991,24(3):18-25.
[4]符书兰,张怀琼,任正隆.黑麦的不同抗条锈病特性在小麦育种中利用价值的评价[J].四川农业大学学报,2005,23(4):379-382.
[5]晏本菊,张怀琼,任正隆.黑麦碱基因(Sec-1)表达缺失的1RS/1BL易位系的鉴定[J].遗传,2005,27(4):514-517.
[6]胡含.非Robertsonian类型小黑麦易位系的研究[J].遗传,2001,23(1):62-63.
[7]李爱霞,亓增军,裴自友,等.普通小麦辉县红荆州黑麦异染色体系的选育及其梭条花叶病抗性鉴定[J].作物学报,2007,33(4):639-645.
[8]宋运贤,李春莲,陈耀锋,等.小麦抗条锈新种质的创制Ⅲ——小麦抗条锈新种质细胞遗传学初步鉴定[J].西北农林科技大学学报,2003,31(6):9-13.
[9]张怀渝 任正隆.威岭栽培黑麦抗白粉病特性导入小麦的研究[J].分子细胞生物学报,2007,40:31-40.
[10]Bennett M D,Gustafson J P,Smit h J B.Variation in nuclear DNA in the genus Secale[J].Chr o moso ma,1977,61:149-176.
[11]蒋华仁,戴大庆,孙东发,等.新合成的小麦遗传资源——几种多倍体小簇麦[J].中国农业科学,1992,25(1):89.
[12]Jiang H R,Dai D Q,Xiao S H.Pr oduction of wheat special ger mplas m[J].J Sichuan Agric Univ,1992,10:255-259.
[13]任正隆,张怀琼.一个改良的染色体C带技术[J].四川农业大学学报,1995,13(1):1-5.
[14]Mukai Y,Nakahara Y,Yama moto M.Si multaneous discri mination of the t hree geno mes in hexaploid wheat by multicolor fluorescence in situ hybridization using t otal geno mic and highly repeated DNA pr obes[J].Geno me,1993,36(3):489-494.
[15]Yang Z J,Li G R,Jiang H R,etal.Expression of nucleol us,endosper m storage pr oteins and disease resistance in an amphiploid bet ween Aegilops tauschii and Secale silvestre[J].Euphytica,2001,119:317-321.
[16]Lukaszewski A J,Gustafson J P.Cytogenetics of triticale[J].Plant Breeding Reviews,1987,10:41-93.
[17]姚启伦,戴玄,李昌满,等.遗传标记在植物遗传育种上的应用[J].涪陵师范学院学报,2004,20(5):86-88.
[18]Gill B S,Friebe B,Endo T R.Standard Karytype and no men-clature system for description of chro moso men bands and str uct ural aberrations in wheat(Triticu m aestivu m)[J].Geno me,1991,34:830-839.
[19]Yang Z J,Li G R,Jia J Q,etal.Molecular cytogenetic characterization of wheat-Secale af ricanu m a mphiploids and the introgression lines for stripe r ust resistance[J].Euphytica,2009,167(2):197-202.
[20]高武军,谢璐,卢靖雯,等.重复序列及异染色质化在植物性染色体重组抑制中的作用[J].遗传,2010,3(1):25-30.
[21]任正隆,张怀琼.小麦-黑麦染色体小片段易位的诱导[J].中国科学(C辑),1997,27(3):258-263.
[22]Luo P G,Zhang H Y,Shu K,etal.Diversity of stripe r ust(Puccinia striifor mis f.sp.tritici)resistance in wheat genotype wit h 1 RS chr o moso mal translocations from different rye lines[J].Can J Plant Pat hol,2008,30:254-259.
猜你喜欢
中国卫生标准管理(2022年21期)2023-01-03
湖北植保(2022年4期)2022-08-23
中国农学通报(2022年21期)2022-08-22
今日农业(2021年7期)2021-07-28
农民致富之友(2019年16期)2019-07-01
现代家庭·生活版(2019年1期)2019-01-24
西藏农业科技(2018年4期)2018-04-25
医学研究杂志(2015年2期)2015-06-10
三联生活周刊(2015年21期)2015-05-21
俄罗斯问题研究(2012年1期)2012-03-25
