
抗浪鱼的核型研究
2012-08-14 08:01孔令富毕保良李莲军王俊相吴志蕾李永能
动物医学进展 2012年12期
孔令富,毕保良,李莲军,汪 霞,王俊相,吴志蕾,李永能,李 洁
(云南农业大学动物科学技术学院,云南昆明650201)
抗浪鱼(Anabarilius grahami)原产于中国第二深高原淡水湖云南省抚仙湖,隶属硬骨鱼纲(Actinopterygii),鲤 形 目 (Cypriniformes),鲤 科 (Cyprinidae),鲌亚科(Cultrinae)白鱼属(Anabarilius)。抗浪鱼是一种珍稀地方鱼种,20世纪90年代以后,由于过度捕捞及银鱼的引进放养,抚仙湖抗浪鱼的产量逐年下降,濒临灭绝。目前,关于抗浪鱼的研究主要是对其生物学特性的描述,包括形态特征、生活习性、生长特点、繁殖习性的描述和濒危原因的分析及对策。目前,除了对抗浪鱼线粒体DNA和遗传多样性有部分研究[1]外,对抗浪鱼的染色体的研究报道较少。
染色体是鱼类遗传的物质基础[2-3],其数目和形态结构具有该物种的特征,一定程度上也反映了生物进化的历史[4],在鱼类的遗传变异中,染色体起着重要的作用,对鱼类染体数量、形态与结构的研究,在遗传学中的基因定位、杂交以及多倍体育种等领域起着重要的作用[5-6]。核型是指把动物、植物、真菌等的某一种有机体或某一分类群的体细胞内整套染色体拍摄下来,按照它们相对恒定特征排列起来的图象。对核型作比较分析主要是确定它们之间在核型上的相同或不同特征,即核型的同源性和异源性,核型的共同特征常是鉴定不同物种或分类群之间近缘关系的重要依据,而对核型异质性的探究是区分不同分类群的重要依据。核型特征主要有染色体二倍数(2n)、染色体分组组成或称核型公式、染色体的臂数、染色体相对长度、臂比,次缢痕的数量等等。鱼类的染色体较小,数目偏多,研究工作难度较大,进展较为缓慢,截止2005年,已报道染色体核型的鱼类有2 100种左右,约占总数10%,且大多数为淡水鱼类,集中在鲤形目、鲶形目、鲈形目上。近年来,对鲤科鱼类染色体研究比较多,发现染色体数2n=100广泛存在于鲤科鱼类中,但核型却各不相同。本研究目的在于通过对抗浪鱼染色体组型的研究,为该鱼种质的细胞遗传学检测提供相应的参数,为抗浪鱼的系统演化、进化地位的确定提供基础资料,同时为抗浪鱼的选育种工作提供细胞生物学、细胞遗传学方面的理论依据。
1 材料与方法
1.1 材料
成年健康抗浪鱼8条,购自云南漫湾有限责任公司水产养殖场;甲醇、冰醋酸、KCl、生理盐水、Giemsa染色原液、植物血凝素(phytohaemagglutinin,PHA)和秋水仙素(colchicine)为宝生物工程(大连)有限公司产品。
1.2 方法
1.2.1 鱼肾细胞悬液的制备、低渗处理及固定PHA经小牛血清溶解后按10μg/g体重胸腔注射抗浪鱼,14h后按1μg/g体重注射秋水仙素,3h后放血取肾,于8g/L的生理盐水中清洗2遍~3遍,然后置于含少量生理盐水的培养皿中,充分剪碎后加入适量生理盐水,用吸管吹打数分钟,即可获得鱼肾细胞悬液。取鱼肾细胞悬液800r/min离心7min后,收集细胞,吹匀后加入预热的0.075mol/L KCl溶液,37℃低渗处理30min~60min。经低渗后的细胞吹匀后加固定液(甲醇∶冰醋酸=3∶1)固定30min,离心800r/min离心10min,重复固定2次。
1.2.2 滴片 将鱼肾细胞悬液滴于预冷(4℃冰箱中预冷)载玻片上,每片1滴~2滴,自然干燥。染色待片干后,用100mL/L Giemsa液(pH 6.8,磷酸缓冲液配制)染色60min,用水充分冲洗,自然干燥后封片。1.2.3 染色体的观察与计数 选取100个不同个体的鱼肾细胞,经显微镜进行观察统计,确定染色体2n数目。在油镜下观察计数100个中期分裂相,在确定抗浪鱼为二倍体的基础上,选取10个左右进行拍照、测量。
1.2.4 核型分析 首先目测照片上每条染色体长度,按长短顺序初步编号,写在每条染色体短臂的一端,同时确定主缢痕的位置,用分规和直尺逐条测量短臂和长臂长度。根据测量的数据,计算染色体的相对长度,臂比和着丝粒指数。
相对长度=(每个染色体的长度/全部染色体长度)×100
臂比(率)=长臂长度/短臂长度
着丝粒指数=(短臂长度/该染色体长度)×100
着丝粒的位置:一般来说,每条染色体着丝粒的位置是恒定的,染色体的两臂常在着丝粒处呈不同程度的弯曲。着丝粒位置的测定常用的方法是以染色体的长臂(L)和短臂(S)的比值来表示(表1)。
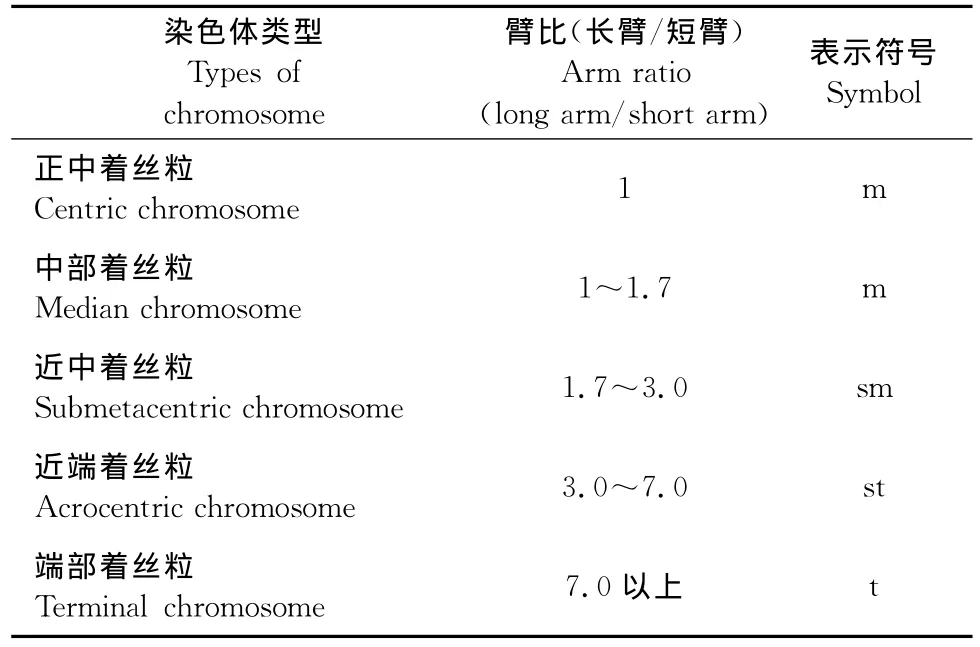
表1 染色体着丝粒位置确定Table 1 The position of chromosome centromere
2 结果
2.1 抗浪鱼的染色体二倍体数目
观察并计数了抗浪鱼的100个清晰的中期分裂相,其染色体分布见表2。
表中统计数字表明,抗浪鱼的细胞染色体数目在46~50之间。染色体总数为48占75%,少于或多于48的染色体占25%。由此可以确定抗浪鱼2n数为48,这同已报道的抗浪鱼2n数一致。
2.2 抗浪鱼的染色体核型
按Levan A等的标准对染色体进行分类、命名,得到抗浪鱼的染色体的相对长度、臂比、类型和着丝粒指数的数据(表3)。
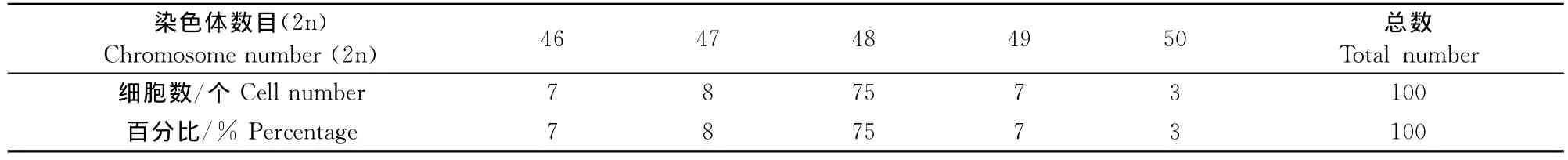
表2 抗浪鱼染色体数目Table 2 Chromosome number of Anabarilius grahami

表3 抗浪鱼核型分析结果Table 3 The results of Anabarilius grahami karyotype

续表3

图1 抗浪鱼染色体中期分裂相Fig.1 The chromosome metaphase of Anabarilius grahami
3 讨论
与其他鱼类相比较,作为云南省重要的地方鱼种,抗浪鱼的分布不广且受到了其他鱼种的冲击,再加上其种质资源的研究开展也较少,并且研究也不够系统,所以抗浪鱼染色体组型的研究成果尚不多见、也不够全面,因此抗浪鱼的核型研究具有较大意义。
3.1 抗浪鱼的核型
根据表3数据可把抗浪鱼的染色体分为3类:A类(编号1~14):共有染色体7对,都为中部着丝粒染色体。B类(编号15~34):共有染色体10对,都为近中着丝粒染色体。C类(编号35~48):共有染色体7对,都为近端着丝粒。因此,可以确定抗浪鱼的核型公式为2n=14m+20sm+14st,染色体的总臂数NF=82。与安晶等[7]在抗浪鱼的二倍体数目、核型公式和染色体的总臂数NF的研究结果一致,试验结果又一次确定了抗浪鱼的核型,为抗浪鱼的进一步研究提供了依据。

图2 抗浪鱼染色体核型Fig.2 The karyotype of Anabarilius grahami
3.2 抗浪鱼与其他鲤科鱼类的核型比较
鲌亚科中的星云白鱼(Anabarilius andersoni)、大鳞白鱼(Anabarilius macrolepis)、取自杞麓湖的银白鱼(Anabarilius alburnops)、取自滇池的银白鱼(Anabarilius alburnops)、黑尾近红鲌(Ancherythroculter nigrocauda)、海南华鳊(Sinibrama melrosei)、银飘鱼(Pseudolaubuca sinensis)、广东鲂(Megalobrama terminalis)的染色体总数是一致的,都是2n=48,而核型公式和染色体的总臂数NF变异范围很大[8-10]。鲌亚科鱼类和抗浪鱼同属鲌亚科的黑尾近红鲌、海南华鳊、银飘鱼、广东鲂只有染色体总数与抗浪鱼相同,在核型公式和染色体的总臂数NF方面的差距较大,表现出了较大的进化差异性。与抗浪鱼同属鲌亚科白鱼属的星云白鱼、大鳞白鱼、银白鱼的染色体总数均为2n=48,与抗浪鱼相同,但在核型公式和染色体的总臂数NF方面又与抗浪鱼略微有所不同,展现出了抗浪鱼在进化上的独特性,体现了不同环境给鱼类进化所带来的不同影响;同时星云白鱼、取自杞麓湖的银白鱼在染色体总数、核型公式、染色体的总臂数NF方面是极其相似的,体现它们在进化上的同源性,这个结论与先前杨君兴的报道中对这3种鱼共有一个最晚近的共同祖先的论述相互印证。从研究结果来看,想要将核型分析的结果做为抗浪鱼种资检测的指标是远远不够的,若能进一步运用染色体带型分析技术、光谱核型SKY技术、分子核型分析技术对抗浪鱼的染色体进行研究,其结果将可能为提高抗浪鱼的研究的准确性提供更有力的辅助。
[1] 杨 博,陈小勇,杨君兴.抗浪白鱼线粒体DNA控制区结构和种群遗传多样性分析 [J].动物学研究,2008,29(4):279-385.
[2] 周丽青,杨爱国,吴 彪,等.波纹唇鱼染色体制备及核型的初步研究 [J].渔业科学进展,2010,31(1):54-58.
[3] 蔡 岩,周永灿,谢瑞敏,等.布氏石斑鱼的染色体核型、银染和 C-带 [J].水产学报,2012,36(5):647-651.
[4] 杨春英,贺一原,郭沐林,等.洞庭湖水系沅水和澧水2种黄颡鱼的形态及染色体组型 [J].湖南文理学院学报:自然科学版,2011,23(4):57-61.
[5] 周丽青,杨爱国,吴 彪,等.波纹唇鱼染色体制备及核型的初步研究 [J].渔业科学进展,2010,31(1):54-58.
[6] 蔡 岩,周永灿,张 本,等.海南野生三斑石斑鱼染色体核型分析 [J].热带生物学报,2011,2(2):98-100.
[7] 安 晶,张桂贤,李运东,等.鲤鱼冻血细胞培养及染色体制备条件优化研究 [J].现代农业科技,2012(2):308-309.
[8] 吉华松,周永灿,蔡 岩,等.六带石斑鱼染色体核型和银染研究 [J].水产科学,2011,30(8):465-466.
[9] 孙增田,宋文华,丁 峰,等.萘醌化合物对斑马鱼肝脏染色体损伤研究 [J].云南大学学报:自然科学版,2012,34(2):238-241.
[10] 牟洪善,王昌留,李金萍.青岛文昌鱼(Branchiostoma belcheri tsingtauense)染色体G类带型研究 [J].海洋与湖沼,2011,42(4):597-602.
猜你喜欢
小学生作文(低年级适用)(2022年12期)2022-12-28
生物技术进展(2022年1期)2022-02-11
宁夏医学杂志(2020年3期)2021-01-21
当代水产(2019年2期)2019-05-16
科学之谜(2018年9期)2018-12-17
阅读与作文(小学低年级版)(2017年10期)2017-10-27
哈尔滨医药(2015年2期)2015-12-01
伴侣(2015年7期)2015-07-16
中国当代医药(2015年9期)2015-03-01
遗传(2015年4期)2015-02-04
