
葱蝇对寄主植物的选择性研究
2012-08-10 02:22:24张云霞苏茂文赵海鹏马晓丹
中国蔬菜 2012年4期
张云霞 王 钲 苏茂文 赵海鹏 马晓丹 薛 明*
(1山东农业大学植物保护学院,山东泰安 271018;2青岛出入境检疫检验局,山东青岛 266000)
葱蝇〔Delia antique(Meigen)〕又称蒜蛆,属双翅目花蝇科,广泛分布于北半球温带地区,是世界性害虫(Poprawski et al.,1985)。葱蝇为寡食性害虫,为害洋葱(AlliuMcepaL.)、大葱(AlliuMfistulosumL. var.giganteumMakino)、大蒜(AlliuMsativuML.)和韭菜(AlliuMtuberosumRottl. ex Spr.)等百合科蔬菜,是重要的地下害虫,在我国以大蒜受害最重。该虫以幼虫蛀食寄主植物的地下部,造成地下部腐烂,整株地上部枯死;春、秋季均可为害,以春季为害最重,田间虫株率一般可达20%~30%,严重时达50%以上,是目前危害我国大蒜生产的重要害虫(王永卫和徐继明,1990)。
对葱蝇的研究,国外在生物学和生态学上研究较多(Ishikawa & Tsukada,1987;Ishikawa et al.,2000;Nomura & Ishikawa,2000,2001),国内在防治方面有一些报道(金运兰 等,2007;王玉桂,2010)。葱蝇为寡食性害虫,其发生程度与寄主植物有密切的关系,昆虫的产卵和取食选择是决定其发生程度的重要因素(刘廷明 等,1989)。本试验研究了葱蝇在 4种不同寄主植物上发生为害情况的差异,以明确成虫产卵选择和幼虫取食选择在其中所起的作用,为掌握该虫的田间数量动态和预测预报、加强田间葱蝇的针对性防治提供理论依据。
1 材料与方法
1.1 葱蝇的饲养
葱蝇于2008年5月采自山东省泰安市郊区范镇大蒜田,养虫室内用大蒜连续饲养多代。温度(20±2)℃,相对湿度60%~70%,光周期L/D=16 h/8 h。
将羽化出的成虫分别放入养虫笼内,笼内悬挂分别装有牛奶、10%糖水和清水的饲养盒。在养虫笼底部放入 1个装有湿沙的产卵盘,并插入新鲜大蒜瓣,以诱集成虫在沙中产卵。每天将产卵盘内的细沙倒入小盆中,倒入清水,轻轻搅拌后卵漂浮到水面上,将卵捞出后得到发育一致的卵。分批连续饲养,获得供试验用的发育一致的成虫和1龄、3龄幼虫。
试验选用的 4种寄主植物均为山东地区普遍种植的常规品种,韭菜品种为寿光独根红;大葱品种为章丘大葱;大蒜品种为金乡红皮蒜;洋葱品种为淄博红皮洋葱。
1.2 葱蝇对寄主植物的选择
1.2.1 发生情况 2010年4月上旬,在葱蝇的发生期,在山东农业大学试验地中选取条件一致的试验小区4块,每小区20 m2。每块地十字型分为4个区域,分别移栽韭菜和苗期的洋葱、大葱、大蒜,罩尼龙网。然后每罩笼内接入 200头葱蝇成虫(雌∶雄=2∶1)。在罩笼内上方中间悬挂盛有脱脂棉的小盘,里面分别加牛奶和清水饲喂,注意及时补充、更换。待葱蝇幼虫发育至2~3龄时(接成虫后15 d),检查不同寄主植物地下根茎部幼虫的数量,计算虫口数量和所占比例。
1.2.2 成虫对寄主植物的产卵选择 采用纱网笼罩盆栽试验。分别在花盆(直径9 cm,高9 cm)中移栽韭菜、苗期的洋葱、大葱和大蒜4种寄主植物。洋葱、大葱和大蒜每盆4株,韭菜每盆8株,植株冠部大小和高度基本一致,土壤表层铺沙。在每个纱笼(60 cm×60 cm×40 cm)的4个角隅处分别放置种植4种寄主植物的花盆各3盆。然后每纱笼放入30头己经交配且卵巢已发育成熟的雌成虫,供其自由选择。注意减少光线不匀的影响,试验条件同上。6 d后采用漂浮法从不同寄主植物带卵的细沙土中漂浮出卵和卵壳,计数。养虫室温度(20±2)℃,相对湿度60%~70%,光周期L/D=16 h/8 h。
1.2.3 幼虫对寄主植物的取食选择 取韭菜和苗期的洋葱、大葱、大蒜4种寄主的地下茎,洗净,擦干,切成2 cm长的小块,两两相对呈十字形放置在铺有滤纸的玻璃培养皿(直径20 cm)的周围。各挑取孵化后第2天的葱蝇1龄幼虫100头,2日龄的3龄幼虫60头,放在培养皿中间,用黑棉布覆盖,每处理6次重复。接虫24 h后,检查不同寄主上幼虫的数量,计算选择率。
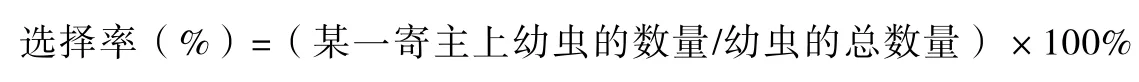
2 结果与分析
2.1 葱蝇在4种寄主植物上的发生数量
在不同寄主植物上葱蝇幼虫的数量,以洋葱最多,大蒜和大葱次之,韭菜上最少,分别占总虫数的40.0%、29.0%、27.0%和4.0%(图1)。
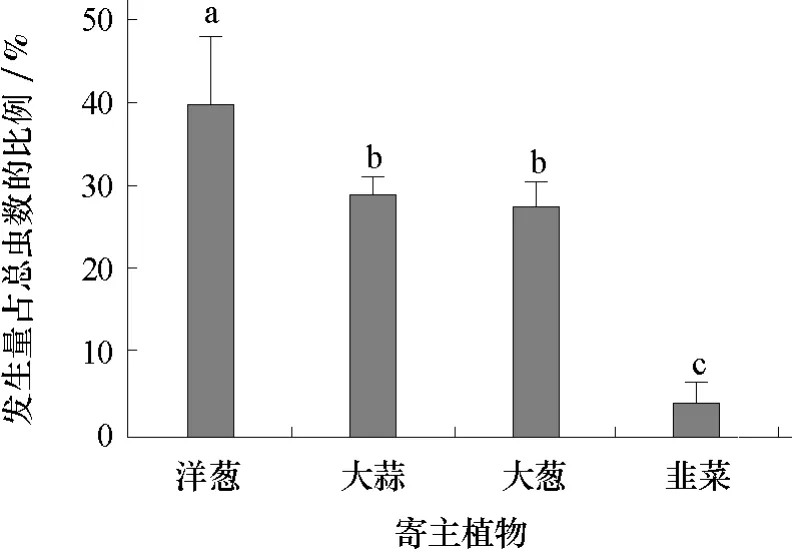
图1 葱蝇在4种寄主植物上的发生数量
2.2 葱蝇成虫对寄主植物的产卵选择
由图2可见,不同寄主上的落卵量依次是洋葱(1 297.3粒)>大蒜(936.0粒)>大葱(901.5粒)>韭菜(309.6粒),分别占总卵量的37.6%、27.2%、26.2%和9.0%。在洋葱上的落卵量最多,韭菜上最少,大蒜和大葱居中,差异不显著。

图2 葱蝇成虫对4种寄主植物的产卵选择
2.3 葱蝇幼虫对寄主植物的取食选择
2.3.1 葱蝇1龄幼虫对寄主植物的取食选择 由图3可见,葱蝇1龄幼虫对4种寄主植物的选择顺序依次为洋葱>大葱>韭菜>大蒜,取食选择率分别为76.4%、16.8%、4.2%和2.6%,其中大蒜和韭菜的差异不显著。
2.3.2 葱蝇3龄幼虫对寄主植物的取食选择 由图4可见,接虫24 h后不同寄主植物上的3龄幼虫的取食选择率依次为洋葱(62.1%)>大葱(23.3%)>韭菜(8.8%)>大蒜(5.8%),其中大蒜和韭菜的差异不显著。和1龄幼虫的取食选择相比较,随着虫龄的增大,葱蝇幼虫对不太喜食的寄主的适应能力有一定的增强。
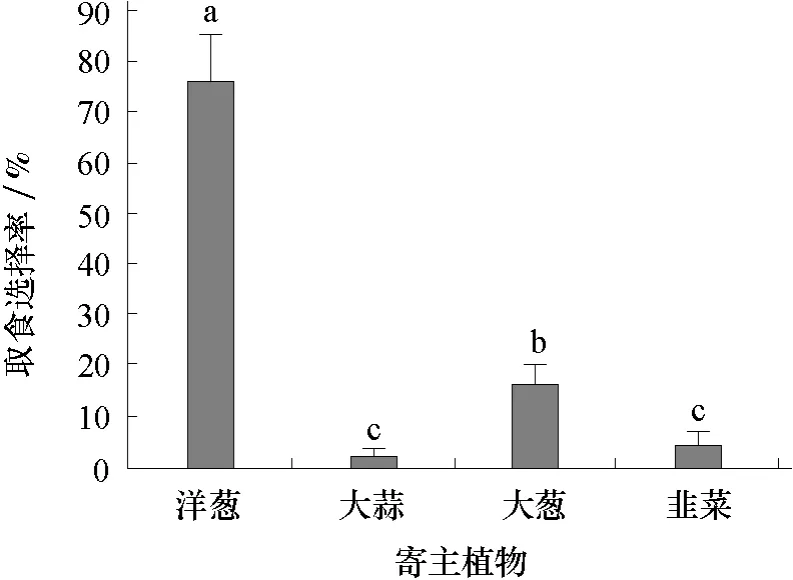
图3 葱蝇1龄幼虫对4种寄主植物的取食选择
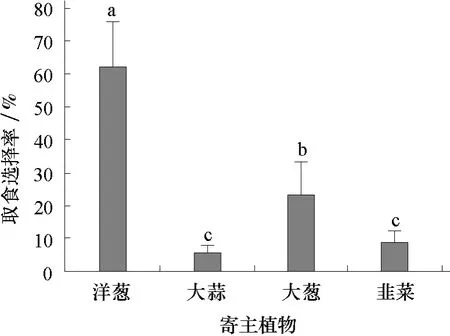
图4 葱蝇3龄幼虫对4种寄主植物的取食选择
3 结论与讨论
葱蝇虽然为寡食性害虫,但在田间不同种类的百合科寄主上的发生程度存在着明显差异。本试验结果表明,在田间罩笼的 4种寄主植物中,以洋葱上葱蝇的发生数量最多,其次为大蒜和大葱,韭菜上数量最少;且盆栽葱蝇成虫产卵选择试验结果与田间罩笼试验结果一致,说明成虫的产卵选择在决定葱蝇在不同寄主的发生数量上起主要的作用。葱蝇成虫的嗅觉系统很发达,(Honda & Ishikawa,1987)电生理实验证明,葱蝇触角嗅觉感受器对二丙二硫醚化合物有反应。据报道洋葱散发的气味中二丙基二硫醚及其相应的烷基硫化物等物质对葱蝇成虫具有引诱力并刺激其产卵(Vemon,1981);大蒜中也含有挥发性物质二烯丙基二硫及其氧化物,还有少量的二烯丙基三硫和二烯丙基四硫(周江菊,2010)。表明百合科中不同的寄主植物对葱蝇成虫选择产卵的引诱力大小不同,可能与含硫化合物的组分和含量不同有关。另外葱蝇对寄主植物的选择除嗅觉外,视觉也起着一定的作用(Ishikawa et al.,1985;Judd & Borden,1991)。
葱蝇幼虫为根蛆类,相对其他昆虫来讲,活动和感觉器官简单,仅能在小范围内选择取食。Soni和Finch(1979)试验结果表明,幼虫对寄主的选择可能利用醇、醛、酸、酯而非单纯利用硫化物作为寻找寄主的线索。所以成虫和幼虫在对寄主的选择喜好上可能存在着差异。而成虫的产卵选择在决定对寄主植物的危害中起着更重要的作用,幼虫的选择仅在同种寄主植株间的选择中起作用。葱蝇成虫和幼虫均更喜欢在有破伤的鳞茎上产卵取食为害(王玉桂,2010)。
葱蝇成虫羽化后需大量取食来补充营养以完成生殖的需求,且成虫寿命长,活动力强(王永卫和徐继明,1990),这也为田间诱杀成虫提供了良好的条件。葱蝇是为害大蒜的重要害虫之一,目前生产上主要在大蒜种植期采用药剂处理土壤和幼虫发生期田间灌根来防治葱蝇(陈凤春,2008),而长期大量使用化学农药导致害虫抗药性增强和产品的农药残留(Eckenrode &Nyrop,1995),因此利用成虫诱杀技术防治葱蝇是实现无公害治理的重要措施之一。
陈凤春.2008.无公害大蒜病虫害的综合防治.绿色植保,3(1):28-29.
金运兰,汪彩云,史慧英.2007.蒜蛆的综合防治.安徽农学通报,(11):160.
刘廷明,刘文英,黄厚英.1989.葱蝇发生规律及防治技术的研究.山东农业科学,(4):15-20.
王永卫,徐继明.1990.葱蝇的发生及防治.新疆农垦技术,(3):17-18.
王玉桂.2010.葱地种蝇的危害及无公害防治技术.吉林蔬菜,(5):64-65.
周江菊.2010.顶空固相微萃取气质联用分析大蒜挥发性风味成分.中国调味品,(9)35:95-99.
Eckenrode C,Nyrop J.1995.Onion maggot managemant in New York,Mchigan and Wisconsin.New York’s Food and Life Sciences Bulletin,144:41-42.
Honda I,Ishikawa Y.1987.Ultrasturcture of the larval cephalic sensory organs of the onion and seed-corn flies,Hylemya antiquaMeigen andH.platuraMeigen(Diptera:Anthomyiidae).Applied Entomology and Zoology,22(3):325-334.
Ishikaws Y,Tanaka S,Matsumoto Y.1985.Color preference of the onion fly with reference to ultraviolet reflection.Applied Entomology and Zoology,20(1):20-26.
Ishikawa Y,Tsukada S,Matsumoto Y.1987.Effect of temperature and photoperiod on the larval development and diapause induction in the onion fl y,Hylemya antiquaMeigen(Diptera:Anthomyiidae).Applied Entomology and Zoology,22:610-616.
Ishikawa Y,Yamashita T,Nomura M.2000.Characteristics of summer diapause in the onion maggot,Delia antiqua(Diptera:anthomyiidae).Insect Physiology,46:161-164.
Judd G J R,Borden J H.1991.Sensory interaction during trap-finding by female onion flies:imp lications for ovipositional host-p lant finding.Entomologia Experimentalis et Applicata,58:239-249.
Nomura M,Ishikawa Y.2000.Biphasic effect of low temperature on completion of winter diapause in the onion maggot,Delia antique.Insect Physiology,46:373-377.
Nomura M,Ishikawa Y.2001.DynaMic changes in cold hardimess,high-temperature tolerance and trehalose content in the onion maggot,Delia antiqua(Diptera:anthomyiidae),associated with the summer and winter diapause.Applied Entomology and Zoology,36:443-449.Poprawski T V,Robert P H,Maniania N K.1985.Susceptibility of the onion maggot to the mycotoxin destructive.Applied Entomology and Zoology,20(7):801-802.
Soni S K,Finch S.1979.Laboratory evaluation of sulphur-bearing chemicals as attractants for larvae of the onion fly,Delia antiqua(Meigen)(Diptera:Anthomyiidae).Bulletin of Entomological Research,69:291-298.
Vemon.1981.Candidate EHV technologies for the post-1980s.Vehicular Technology Conference,4:49-60.
猜你喜欢
英语世界(2023年11期)2023-11-17 09:24:46
小星星·阅读100分(低年级)(2018年4期)2018-05-07 07:25:40
乡村地理(2018年4期)2018-03-23 01:53:36
小猕猴智力画刊(2017年9期)2017-10-19 01:01:57
现代园艺·综合版(2017年1期)2017-07-14 16:37:59
江苏农业科学(2017年7期)2017-05-23 23:10:25
红领巾·萌芽(2017年2期)2017-03-09 15:22:59
农业与技术(2016年19期)2016-12-12 02:23:50
中国蔬菜(2015年9期)2015-12-21 13:04:42
新作文(小学中高年级版)(2015年5期)2015-04-12 06:27:42
