
池养脊尾白虾形态性状对体重的影响效应
2012-07-19 08:46吕敢堂王志铮任夙艺
浙江海洋大学学报(自然科学版) 2012年3期
杨 磊,赵 晶,杨 鹏,吕敢堂,王志铮,任夙艺
(浙江海洋学院水产学院,浙江舟山 316004)
形态性状既是水产经济动物种质遗传信息的外在综合体现,也是表征水产动物生长性能和生产性能的重要指标。利用形态性状构建基于估算重量性状的多元回归模型已日渐引起研究者们的兴趣,DEBOSKI等[1]运用形态性状建立了可估算大西洋鲑Salmo salar L.体脂肪含量的多元回归模型,刘小林等[2]研究了决定凡纳对虾Penaeusvannamei体重的关键形态性状组合,孙振兴等[3]定量分析了形态性状对扁玉螺Neveritadidyma软体部重的影响效应,王志铮等[4-5]在揭示了决定日本沼虾Macrobrachium nipponensis体重的关键形态性状组合存在明显性别差异的基础上,确定了决定该虾个体繁殖力的关键形态性状组合。因此,运用多元统计学方法精确找寻决定水产养殖动物目标经济性状的关键形态性状组合,对于指导该水产养殖动物的人工选育无疑具有重要的现实意义[2-12]。
脊尾白虾Exopalaemon carinicauda隶属于甲壳纲Crustacea、十足目Decapoda、长臂虾科Palaemonidae、白虾属Exopalaemon,俗称白虾、小白虾,是我国重要的养殖经济虾类之一,其因适应性较强、食性杂、养殖周期短、产量高、肌肉品质优良等特点,近年来逐渐为广大养殖业者和科研工作者所重视,是一种极具开发潜力的养殖虾类品种[14-17]。目前,有关脊尾白虾的研究主要集中于生长繁殖[14-16]、生理生态[17-18]、毒理病害[19-20]等方面,而关于形态性状对其体重影响效应的研究迄今尚未见报道。鉴于此,本文作者以人工养成的3月龄脊尾白虾同生群为实验对象,采用多元统计学方法研究了脊尾白虾形态性状对活体重的影响效应,以期为该虾人工选育提供基础资料。
1 材料与方法
1.1 样品来源及其生物学指标的测定
试验用脊尾白虾购自舟山市定海区文龙养殖场,于实验室暂养2 d后随机选取90尾肢体完整、健壮有活力的个体进行生物学测定。实验时,用滤纸吸净实验虾体表水分后用BS223S型电子天平(精度1 mg)称量其体重(BW);参照王志铮等[4]的测量方法用游标卡尺(精度0.02 mm)逐尾测量体长(BL)、头胸甲长(CL)、头胸甲宽(CW)、头胸甲高(CH)、腹部高(AH)、腹部宽(AW)、额剑长(RL)等 7项长度性状,并按赵晓勤等[21]的方法测量腹部长(AL)和尾扇长(FL)。
1.2 分析方法
整理所得数据,分别计算各性状的均值(M)、标准差(SD)和变异系数(CV%)。运用SPSS17.0软件在表型相关分析的基础上,根据通径分析原理建立相关矩阵正则方程组并得到通径系数(Pi)后,选取通径系数达显著水平(P<0.05)的性状,按相关系数组成效应(rxiy=Pi+∑rijPj),将这些性状与体重间的相关系数(rxiy)剖分为各性状对体重的直接作用(Pi)和通过其他性状影响体重的间接作用(rijPj),据此计算各形态性状对体重的决定系数(di)和共同决定系数(dij)。采用逐步导入剔除法进行偏回归分析,剔除偏回归系数未达显著水平的性状(P>0.05),建立形态性状对体重的多元回归方程,并进行拟合度检验。
2 结果
2.1 脊尾白虾生物学测定指标的参数估计值
由表1可见,脊尾白虾体重(BW)的变异系数最大,其值为所测形态性状的1.8倍以上,具较高的选育潜力,而所测各形态性状的变异系数则均较为接近。
2.2 脊尾白虾生物学测定指标间的相关性
由表2可见,除RL与AL间相关系数未达显著水平(P>0.05)外,其余相关系数均达极显著水平(P<0.01)。其中,宽度性状、高度性状与体重的相关系数值均较稳定,其范围分别为 [0.905,0.906]和[0.850,0.887],而长度性状与体重的相关系数值则波动较大,其范围为[0.607,0.943]。

表1 脊尾白虾生物学测定指标的参数统计(n=90)Tab.1 The apparent statistics of various trait for Exopalaemon carinicauda(n=90)
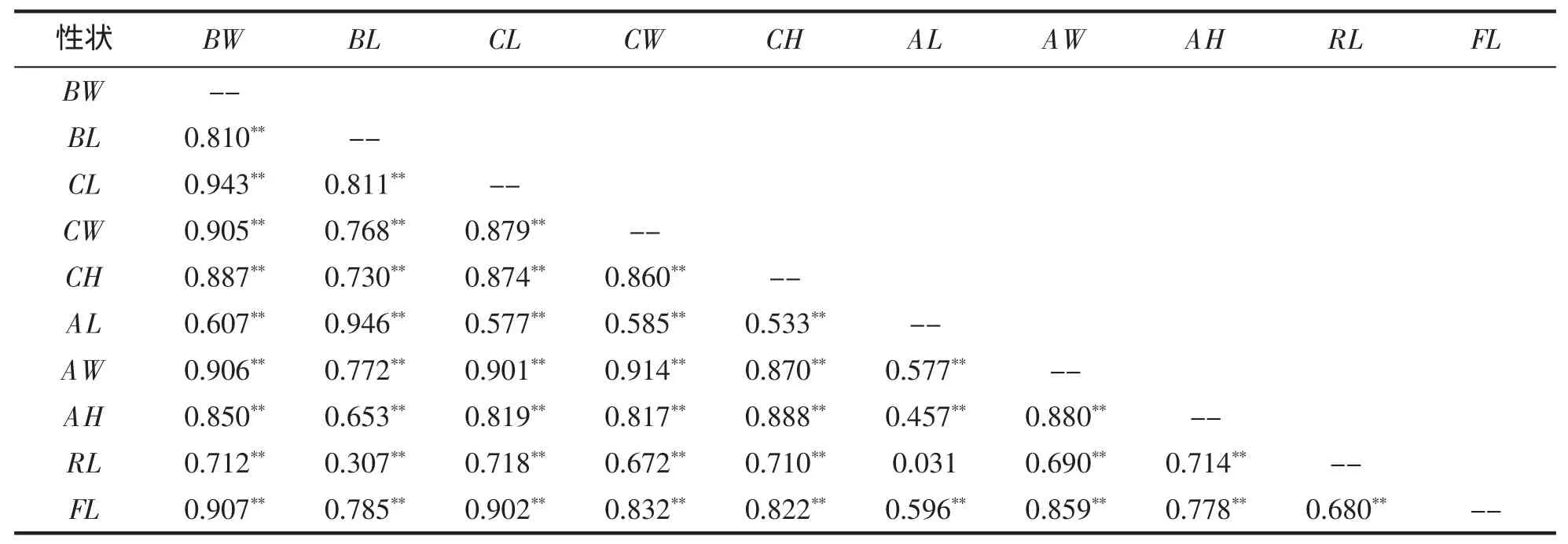
表2 脊尾白虾各性状间的相关系数(n=90,df=88)Tab.2 The phenotype correlation coefficient between the traits of E.carinicauda(n=90,df=88)
2.3 脊尾白虾形态性状对体重的通径分析
由表3可见,对体重的直接作用达到显著水平(P<0.05)的形态性状为CL、CW、FL和AH。它们对体重的直接作用呈CL>CW>FL>AH,与间接作用排序正好相反,且CW、FL、AH对体重的直接作用均小于通过CL对体重产生影响的间接作用,表明CL为决定体重的关键性状,其余3个性状为决定体重的主要性状。
2.4 脊尾白虾形态性状对体重的决定程度分析
由表 4 可见,就单一性状对体重的决定程度而言,呈 CL(0.175)>>CW(0.056)>FL(0.050)>>AH(0.020);就性状两两组合对体重的决定程度而言,以CL-CW(0.174)、CL-FL(0.168)为最大,且其决定系数值远高于其它组合,表明CL为决定该虾体重的关键变量,CW和FL为决定该虾体重的重要变量,AH为决定该虾体重的从属变量。
2.5 脊尾白虾形态性状与体重间多元回归方程的构建
经多元回归分析(表 5~7),保留偏回归系数达到显著水平(P<0.05)的 CL、CW、FL 和 AH,得到可准确估算脊尾白虾体重的多元回归方 BW=1.847 CL+1.657 CW+1.110 FL+0.954 AH-3.825(R2=0.931,P<0.01);经检验,所建方程对体重的估计值与实测值间无显著差异。

表3 脊尾白虾形态性状体重影响的通径分析Tab.3 The effect of morphometric traits on body weight of E.carinicauda
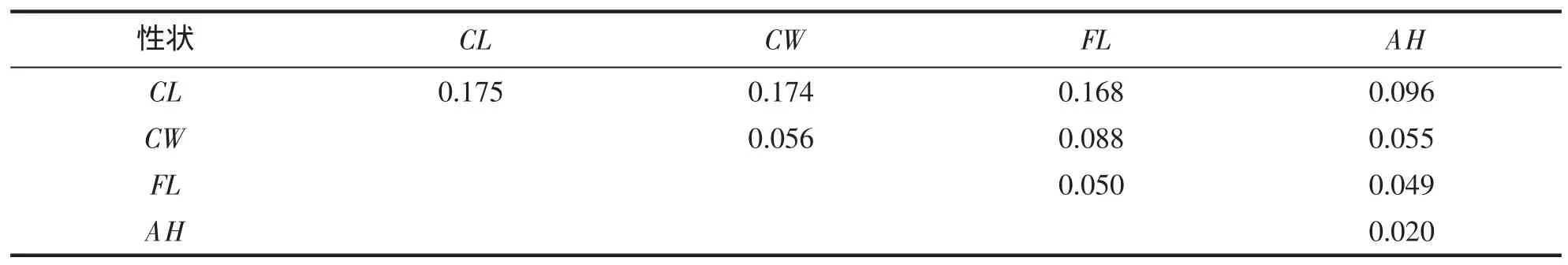
表4 脊尾白虾形态性状对体重的决定系数Tab.4 Determinant coefficients of morphometric traits on body weight of E.carinicauda
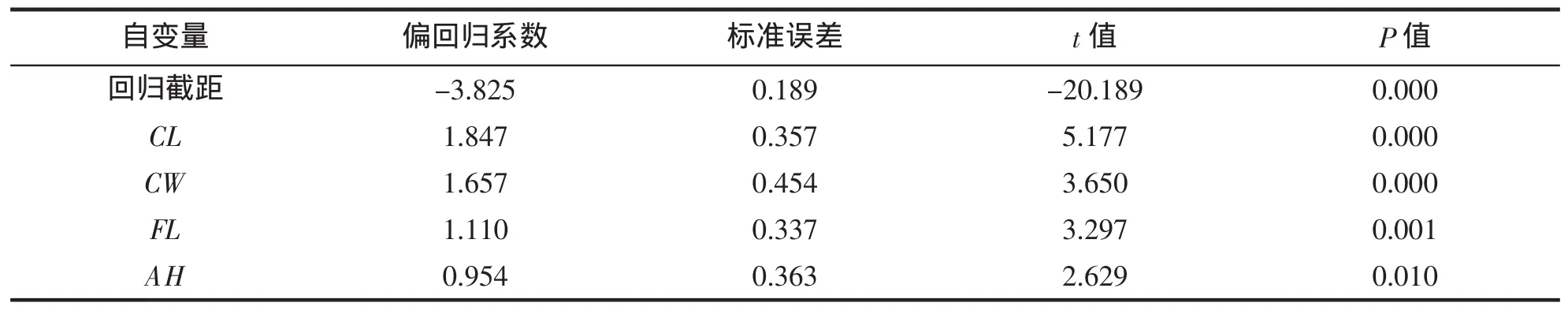
表5 偏回归系数和回归常数显著性检验Tab.5 Test significant of partial regression and intercept
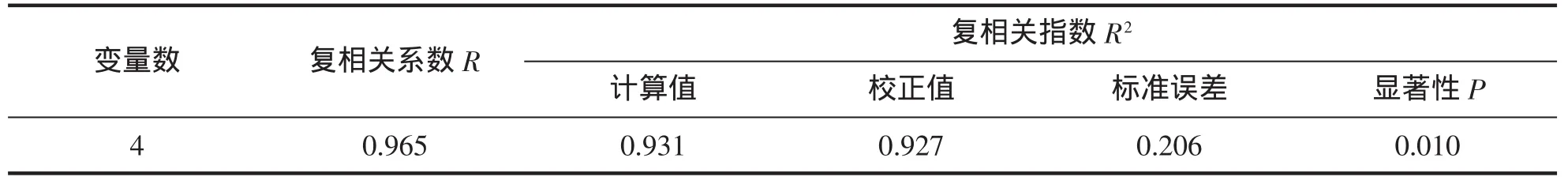
表6 脊尾白虾形态性状与体重的复相关分析Tab.6 The multiple-correlation coefficients of morphometric traits to the weight of E.carinicauda
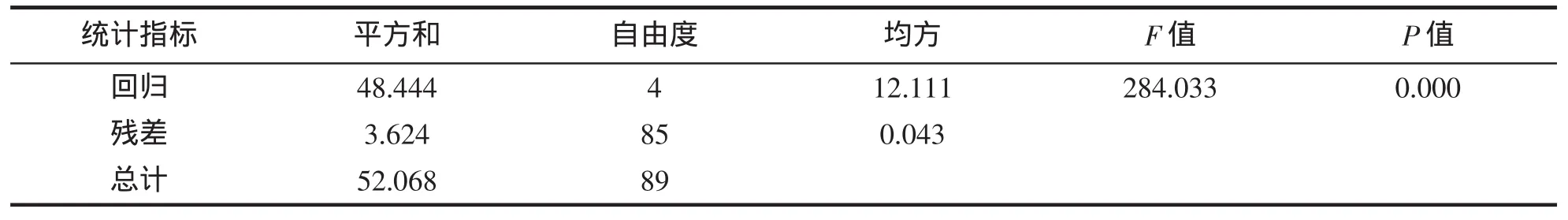
表7 多元回归方程的方差分析Tab.7 Analysis of variance of the multiple regression equations for E.carinicauda
3 讨论
3.1 决定脊尾白虾体重的关键形态性状组合的确定
通径分析是一种可揭示变量间的相互关系的有效数学手段,在水产经济动物良种选育中,其被广泛运用于确定决定水产经济动物体重的关键形态性状组合[2,4-10]。已有的研究表明,虾类重量性状的变异系数均明显高于其形态性状,说明重量性状具较高的选育潜力,这与本研究的结果(表1)相吻。由表2可见,脊尾白虾体重与形态性状间的相关性均达到极显著水平(P<0.05),但并非所有性状均可融入决定其体重的关键性状组合中,而找寻决定其体重的关键性状组合尚需依赖多元回归分析得以实现。通径分析表明,头胸甲长(CL)、头胸甲宽(CW)、尾扇长(FL)和腹部高(AH)等4项形态性状对该虾体重的直接作用均达到了显著水平(P<0.05),它们对该虾体重的总决定程度高达 93.1%(∑d=R2=0.931),高于刘小林等[2]所指出的决定系数下限(R2=0.85),说明决定该虾体重的关键形态性状组合已经找到。
3.2 CL、CW、FL和AH为影响脊尾白虾体重的关键形态性状组合的原因分析
头胸甲是甲壳动物的摄食与运动中心,故头胸部相关形态性状对体重具决定作用。已有的研究表明,决定中国明对虾Fenneropenaeus chinensis、凡纳对虾、日本囊对虾Marsupenaeus japonicus和日本沼虾体重的关键形态性状组合均包含CL和CW,与本研究的结果相吻。沈嘉瑞等[22]指出,脊尾白虾有跳跃运动的习性主要是借腹部的急剧屈曲和尾扇向下拨水使身体腾空来实现的;沈辉等[23-24]进一步观察发现该虾最主要的防御方式就是运用尾扇拨水向后弹开,且具相残行为。动物的防御行为可分为初级防御(Primary defence)和次级防御(Secondary defence)[25]。虾类在自然生境中以潜底、隐蔽等初级防御为主,而在养殖环境中则以连续的弹跳次级防御为主导[26-28]。据此可认为,养殖环境中脊尾白虾因空间拥挤程度的加剧,使得次级防御行为的发生频率也大大提高,故不难理解其次级防御行为的关键部位,即尾扇和腹部的发达程度对体重有着重要的决定作用,致使FL和AH融入决定其体重的关键性状组合中。
[1]DEBOSKI P,DOBOSZ S,ROBAK S,et al.Fat level in body of juvenile Atlantic salmon (Salmo salar L.),and sea trout(Salmo trutta M.trutta L.),and method of estimation from,orphometric data[J].Archives of Polish Fisheres,1999,7(2):237-243.
[2]刘小林,吴长功,张志怀,等.凡纳对虾形态性状对体重的影响效果分析[J].生态学报,2004,24(4):857-862.
[3]孙振兴,常林瑞,徐建鹏.扁玉螺(Neverita didyma)表型性状对体重和软体部重的影响效应分析[J].海洋与湖沼,2010,41(4):513-518.
[4]王志铮,吴一挺,杨 磊,等.日本沼虾(Macrobrachium nipponensis)形态性状对体重的影响效应[J].海洋与湖沼,2011,42(4):612-618.
[5]王志铮,杨 磊,施建军,等.日本沼虾(Macrobrachium nipponensis)池养越冬抱卵亲青虾表型性状对个体繁殖力的影响效应[J].海洋与湖沼,2012,43(1):166-173.
[6]孙成波,邓先余,李镇泉,等.北部湾野生日本囊对虾(Marsupenaeus japonicus)体重和形态性状的关系[J].海洋与湖沼,2008,39(3):263-268.
[7]董世瑞,孔 杰,万初坤,等.中国对虾形态性状对体重影响的通径分析[J].海洋水产研究,2007,28(3):15-22.
[8]安 丽,刘 萍,李 健,等.“黄海1号”中国明对虾形态性状对体质量的影响效果分析[J].中国水产科学,2008,15(5):779-786.
[9]吴立峰,张吕平,沈 琪,等.凡纳滨对虾不同家系的形态性状对体重的影响[J].海洋湖沼通报,2010(2):37-48.
[10]何吉祥,丁凤琴,宋光同,等.池养克氏原螯虾形态性状对体质量的影响[J].安徽农业大学学报,2011,38(3):352-357.
[11]刘春雷,常玉梅,梁利群,等.转大麻哈鱼生长激素基因鲤表型性状与体质量的相关性及通径分析[J].应用生态学报,2011,22(7):1 893-1 899.
[12]刘 磊,李 健,高保全,等.三疣梭子蟹不同日龄生长性状相关性及其对体重的影响[J].水产学报,2009,33(6):964-971.
[13]刘瑞玉.中国北部经济虾类[M].北京:中国农业出版社,1995:48-49.
[14]李明云.池养脊尾白虾的繁殖、生长及其最大持续轮捕量的初步探讨[J].水产学报,1994,18(2):85-92.
[15]王兴强,阎斌伦,马 甡,等.脊尾白虾生物学及养殖生态学研究进展[J].齐鲁渔业,2005,32(8):21-23.
[16]徐君义.浙江乐清湾脊尾白虾的繁殖和世代的初步研究[J].动物学杂志,1990,25(6):3-7.
[17]罗会明,黄厚哲.脊尾白虾幼体对饵料的摄食与吸收[J].厦门大学学报:自然科学版,1980,19(4):100-107.
[18]王兴强,曹 梅.低盐和低温对脊尾白虾生长和能量收支的影响[J].水生态学杂志,2010,3(2):66-71.
[20]王李宝,万夕和,朱 彬,等.二硫氰基甲烷对脊尾白虾的急性致毒实验[J].海洋水产研究,2008,29(5):120-123.
[21]CAO Mei,WANG Xing-qiang,YAN Bin-lun,et al.Effects of salinity fluctuation and dietary traditional Chinese herbal medicines on survival,growth and immunity of Exopalaemon carinicauda[J].Agricultural Science&Technology,2010,11(1):77-81.
[22]赵晓勤,倪 娟,陈立侨,等.日本沼虾4种群的形态差异分析[J].中国水产科学,2006,13(2):224-229.
[23]沈嘉瑞,刘瑞玉.我国的虾蟹[M].北京:中国青年出版社,1976:11-36.
[24]沈 辉,万夕和,王李宝,等.3种肉食性鱼类的捕食及脊尾白虾防御行为比较[J].水产养殖,2010,31(1):40-42.
[25]沈 辉,万夕和,许 璞,等.脊尾白虾的行为学观察研究[J].海洋科学,2010,34(10):53-56;61.
[26]尚玉昌.行为生态学[M].北京:北京大学出版社,1998:253-269.
[27]于赫男.环境胁迫对罗氏沼虾和凡纳滨对虾行为、生长及生理活动的影响[D].广州:暨南大学,2007.
[28]DANIEL T L,MEYHOFER E.Size limits in escape locomotion of carridean shrimp[J].J Exp Biol,1989,143(2):245-265.
[29]DALL W,HILL B J,ROTHLISBERG P C.Biology of the Penaeidae[M]//BLAXTER J H S,ed.Advances in Marine Biology.London:Aeademic Press,1990.
猜你喜欢
当代水产(2022年8期)2022-09-20
食品工业(2022年7期)2022-08-04
当代水产(2022年5期)2022-06-05
黑龙江气象(2021年2期)2021-11-05
当代水产(2021年8期)2021-11-04
当代水产(2021年4期)2021-07-20
当代水产(2020年3期)2020-06-15
数学学习与研究(2019年20期)2019-11-30
课程教育研究(2018年4期)2018-03-16
餐饮世界(2014年6期)2014-08-16
