
贵州瓮安陡山沱组碳同位素特征及古海洋意义
2012-07-06 07:17:30密文天王新利冯志强陈安清任才云周玉华
成都理工大学学报(自然科学版) 2012年3期
密文天 王新利 冯志强 陈安清 任才云 周玉华
(1.成都理工大学 沉积地质研究院,成都610059;2.中国地质大学 地球科学与资源学院,北京100038;3.浙江大学 海洋科学与工程学系,杭州310058;4.贵州瓮福磷矿,贵州 福泉550508)
贵州瓮安震旦系陡山沱组发现的瓮安生物群是目前发现的最早的动物化石群。较早的早期生命证据是澳大利亚太古宙3 500Ma B.P.的Warrawoona群微生物化石和南非的太古宙Fig Tree群中的化石[1];但直到新元古代末期以前,仍没有动物出现。1998年,Xiao在《Nature》上发表文章称在中国瓮安发现动物胚胎化石[2],这是迄今为止最早的动物化石记录。根据生物与环境协同演化的理论,推测埃迪卡拉纪陡山沱期的古海洋环境应发生了很大变化,有必要通过研究该时期的δ13C值变化,探讨其环境的变化。
重大地质事件发生时海相碳酸盐岩的碳同位素组成演化表现出一定的飘移规律。地质事件打破了原有的生物圈、水圈、大气圈之间的碳同位素动态平衡,导致各圈层内含碳物质的碳同位素组成有明显变化;而海相碳酸盐岩反映同期海水碳同位素组成,可以通过碳稳定同位素偏移来反映地质事件。例如,黄思静在泥盆系-石炭系、石炭系-二叠系、二叠系-三叠系界线附近发现δ13C值负漂移,认为与生物绝灭更替有关[3]。而震旦纪沉积地层记录的碳同位素异常变化幅度很大,反映了异常的海洋地球化学条件[4]。目前,整个华南震旦系台地相剖面共报道了4个碳同位素负异常[5],其中陡山沱期分辨出较为明显的2个负异常。最早的异常出现于陡山沱组底部盖帽碳酸盐岩中(约635Ma B.P.),并提出了“雪球地球”假说、上升流假说、甲烷逃逸和“淡水分层模式”假说来解释[6];第二个负异常出现在陡山沱组中部,出现的时间有580Ma B.P.和595Ma B.P.两种观点[4,5]。而在陡山沱组-灯影组界线附近的碳同位素异常被认为可以与阿曼Shuram异常对比[7]。
此外,磷是携带基因的DNA及RNA的主要化学成分,也是携带能量的三磷酸腺苷及细胞隔膜磷脂的核心成分。没有磷的参与,任何生物创新事件都是无法完成的。与后生动物同期发生的是地史时期第一次出现的成磷事件,而瓮安后生动物胚胎化石就发现于磷块岩中。研究δ13C值的变化对探索成磷事件及生物事件有一定的意义。
1 区域地质背景
瓮福磷矿位于贵州省中部瓮安县、福泉县境内,南北长约20km,东西宽2~4km(图1)。大地构造上属于扬子地台黔中隆起的北部。矿区地层属于白岩背斜构造,中部由小坝断层截断,北部为白岩背斜,南部为高坪背斜。背斜核部出露最古老的地层为前震旦系板溪群清水江组,向两侧依次出露震旦系、寒武系。

图1 贵州瓮安磷矿区域地质图Fig.1 Regional geological map of the Weng'an phosphorite deposit in Guizhou
陡山沱组含磷岩系沉积在南沱冰期的南沱组冰碛岩之上,上覆地层为灯影组白云岩。陡山沱期该区处于瓮昭隆起向北东倾斜的浅水环境,最初沉积了陆源碎屑岩和下白云岩段;富磷海水侵入后在北部沉积了下磷层,后来海平面下降遭受剥蚀;最后海平面再次上升沉积了磷块岩层。研究区位于穿岩洞矿段及大塘矿段的陡山沱组剖面,大塘剖面自下而上大体分为4层:(1)下矿层段,下部为含泥磷块岩,厚1.16m;上部为白云质磷块岩及硅质磷块岩,厚4.64m。(2)夹层段,硅质岩,厚2.1m。(3)上矿层段,主要磷矿层,由下而上为硅质磷块岩、白云质磷块岩,厚2.8m;中上部为白云质砂屑磷块岩,厚5.25m。(4)底板段主要为海绿色泥岩及泥质白云岩,厚10.13m。样品分别采自大塘矿段和穿岩洞矿段。
2 分析方法及实验结果
2.1 干酪根碳同位素组成分析
采用化学及物理方法,除去岩石中的无机矿物及可溶有机质,并使不溶有机质富集。步骤:(1)酸处理,将岩样用蒸馏水浸泡,使泥质膨胀,除去上部清液;将6mol/L的盐酸加入样品,升温至60℃,搅拌反应2h,洗涤到弱酸性,除去清液;将样品用水浴锅蒸去水分,加入6mol/L的盐酸及质量分数为40%的氢氟酸,升温至60℃,搅拌反应2h,用稀盐酸洗涤3次。按上述步骤反复4次。(2)重液浮选,将样品烘干4h,磨碎后放入离心管,加入相对密度为2.0~2.1的重液,高速离心,取出上部干酪根,重复2次,用AgNO3检查有无卤离子。(3)将干酪根在-5℃冷冻6h,后进行干燥。(4)使系统真空至4Pa,通入氧气,调好-50℃酒精冷液并套到冷阱上,将有样品的瓷舟置于冷端,高温燃烧25min,开启真空泵至4 Pa,冷冻收集经冷阱脱水后的CO2。用气体同位素质谱仪测量的δ13C值(PDB标准),精度为±0.2‰。
2.2 无机碳、氧同位素组成分析
用饱和磷酸法获取样品反应后的CO2。纯灰岩在75℃、25℃时,反应时间分别为0.5h、4h,纯白云岩为6h、40h。用液氮法冷冻收集脱水后的CO2。实验条件为真空度2Pa,100%饱和磷酸,仪器为MAT252气体稳定同位素质谱仪,GBW04406为控制标准。精度为±0.2‰,以PDB为标准。
2.3 实验结果及检验
瓮安陡山沱组含磷岩系碳酸盐岩及干酪根的同位素组成列于表1和表2。干酪根碳同位素组成在-31.3‰~-32.32‰间变化,宜昌干酪根碳同位素为-28.97‰~-38.39‰。瓮安含磷岩系中白云岩的δ13C值为-2.45‰~1.75‰,δ18O值主要分布在-1.70‰~-8.82‰。
海相碳酸盐岩的碳同位素组成可能会受早期成岩作用、晚期成岩作用中流体的作用等影响[8],所测的碳同位素组成来自白云岩,其对海水信息的代表性需要讨论。海相碳酸盐的成岩蚀变过程伴随着 Mn的获取和Sr的失去[9,10],蚀变程度较低的海相碳酸盐有较低的Mn含量、更高的Sr含量。Derry认为Sr的质量分数>0.2‰时,能较好地代表海水信息[9];Kaufman等认为当碳酸盐中wMn/wSr<2~3时,能很好地保持原始海水的同位素组成[11];也有人认为 wMn/wSr<10表明碳酸盐岩未遭受强烈的蚀变,也可以作为碳酸盐岩保留了原始同位素的判别标志[12-14]。黄思静在秀山下古生界剖面研究中,认为微-粉晶白云石有较低的wMn/wSr比值,能代表海水信息,某些白云石Sr和Mn含量低的原因可能是沉积因素造成的[10]。本文的样品为采集的新鲜的结晶程度低的微-粉晶白云岩,避免采集重结晶作用强的样品,并回避碳酸盐脉,样品缺乏方解石胶结物,陆源碎屑也较少。总之,碳酸盐的结构特征以微-粉晶白云石为主。

表1 瓮安大塘剖面陡山沱组白云岩碳氧同位素组成Table 1 C and O isotope compositions of the dolomite from Doushantuo Formation in Weng'an

表2 瓮安陡山沱组样品干酪根碳同位素组成Table 2 δ13 Ckercompositions of the samples from Doushantuo Formation in Weng'an
白云岩样品wMn/wSr的元素比值均<10;底板段的 WD24、WD28的wMn/wSr比值较高,可能是因受锰矿化影响而具有高的Mn含量,Mn含量可与世界范围内其他地区同一时期的地层对比[15]。其余的样品 wMn/wSr基本达到了要求的标准,可以反映同期海水包含的信息。
碳酸盐岩的氧同位素组成对蚀变作用灵敏,水-岩交换作用能使原岩δ18O值降低,样品的氧同位素值均为>-10‰(PDB),表明样品很好地保存了原始海水的同位素组成。δ13C与δ18O二者是否存在线性相关关系,也能一定程度上判断样品是否遭受后期改造。Bathurst认为这是由成岩作用导致的[16]。瓮安δ13C与δ18O相关性很小,相关系数为0.16(图2)。
干酪根性质十分稳定,是耐降解大分子保存的结果。用它来测定样品中的δ13Corg值,比用相同样品中的总有机碳来测定更为准确[17]。有机碳同位素值与沉积环境和植物来源关系密切,但在成岩作用中,热化学反应可能导致有机碳同位素值改变;其他地层的可溶有机质对目标地层的混入污染,也可能使数值偏离[18]。干酪根的H/C原子比可以对干酪根保存程度进行评价,H/C原子比>0.2的样品,受到后期成熟作用的影响较小[17]。瓮安磷矿剖面样品的 H/C原子比均>0.2,表明样品受后期作用的影响很小。

图2 大塘剖面碳氧同位素组成相关图Fig.2 δ13 C-δ18 O plots for all tested samples from the Datang section
3 碳同位素组成特征
海水碳同位素组成受碳酸盐沉积物和有机碳的相对比例影响;海水中碳酸盐的沉积,不会引起碳同位素的分馏,因此与其形成介质的碳同位素组成相同[19,20]。而有机碳的同位素值很低,这与生物动力学分馏有关。当生物氧化/埋藏量发生变化时,由于有机质碳同位素值与海水同位素值相差较大[21],会影响到海水及其同期海相碳酸盐岩同位素组成。
3.1 大塘剖面碳酸盐岩碳同位素组成
瓮安陡山沱组含磷岩系大塘剖面底板段碳酸盐岩的δ13C值均为负值(图3)。从底板段向上至下矿层,δ13C值发生明显的正偏移,向上至下矿层下部δ13C值基本保持为正值,下矿层中上部δ13C值逐渐负偏移。在夹层段,δ13C值仍然较偏负;在上矿层,δ13C值又重新升高,保持在1‰~1.7‰。Hoffman等报道的陡山沱期早期的帽碳酸盐岩的δ13C值一般在-2‰~-3‰之间[22],世界其他大陆也有类似情况。瓮安陡山沱组底板段的碳酸盐岩可以与全球性的帽碳酸盐岩相对比,δ13C值维持在-1.85‰~-2.45‰;在大塘剖面底板段顶部,则上升到0‰左右。因此,在底板段δ13C值总体为上升趋势。到下矿层开始出现大规模磷块岩沉积时,δ13C值已经接近于1‰。

图3 瓮安大塘剖面陡山沱组碳酸盐岩碳同位素组成变化曲线Fig.3 Variation curves of C isotope in the carbonate rocks from Doushantuo Formation of the Datang section in Weng'an
新元古代公认有2期冰川事件,即Sturtian冰期和 Marinoan或Vrangerian冰期[23],而南沱冰期可以与Marinoan冰期相对比。有机地球化学证据表明南沱冰期时古海洋中的光合作用很受限制,生物生产率异常低下[24];冰期后的陡山沱早期古海洋生态系统开始重建,藻类复苏使生物生产率开始恢复。因此,海洋浮游生物的光合作用产生的同位素分馏及其有机碳埋藏率的变化,可能导致了底板段δ13C值的升高。所以,陡山沱早期的新属种对有机质的生产量逐渐增加,使同期海相碳酸盐岩的δ13C值呈现正偏移,反映古海洋生物圈重新恢复生机。下矿层获得的δ13C值(WD16)继续升高,但与宜昌田家园子同层位对比[15],δ13C值较低,可能是区域环境差异对同位素差异的影响所致。而下矿层的中上部δ13C值有一定的负漂移,显示了生物生产率的降低,这反映了陡山沱期古海洋环境的不稳定性,气候与环境都有一定的波动与反复。
到陡山沱期中期,δ13C值仍然有明显的负偏移,在大塘剖面的夹层段δ13C值(WD24)为-0.08‰。夹层段出现的明显的沉积间断面及古喀斯特现象的发育表明这一时期发生了地表暴露事件,可能与小规模的Gaskiers冰期(约580Ma B.P.)有关,或其他地质事件导致了生物生产率的又一次下降。该期的δ13C值负偏移表明当时处于生态相对萧条时期。经过陡山沱中期的环境的剧烈变化之后,δ13C值又开始上升,获得的陡山沱组上磷矿层中上部的δ13C值为1.75‰~0.84‰。这说明生物进入新的复苏与繁盛期,瓮安生物群化石中的后生动物胚胎化石就是在上矿层下部发现的。Lambert等对扬子地区震旦系陡山沱组碳同位素的测定后发现碳同位素也有数次波动[25],与大塘剖面的δ13C值的数据特征相符合。这些波动反映了该时期全球的岩石圈、水圈和生物圈的快速变化。而环境与生物是协同演化的,环境的快速变化刺激了生物的进化。
3.2 穿岩洞剖面有机碳同位素组成
Lewan把显生宙时期沉积岩中无定形干酪根的碳同位素组成分成相对富12C的L型和相对富13C的H型的2种无定形干酪根[26]。瓮安穿岩洞剖面获得的干酪根δ13C数值低于-26‰,应属于L型无定形有机质;且δ13Corg数值大多低于-30‰(表2),即使在L型无定形有机质中,它们更富12C。与李任伟等获得的陡山沱期黑色页岩δ13Corg值十分一致[27]。
δ13Corg值与海水溶解碳的碳同位素组成和总量变化有关[28],生物初级生产率、生物量、沉积速率、大气CO2浓度及风化等都可对其造成影响。生产率较高时,营光合作用的浮游生物等从水体中的CO2摄取碳引起生物细胞内外CO2不平衡,使浮游生物δ13Corg值增加;生产率较低时,浮游生物碳需求量降低,引起水溶液中溶解的CO2累积,导 致δ13Corg值 的 降 低[29]。 上 矿 层 样 品 的δ13Corg值自下而上数值逐渐降低(图4),浮动达到1.02‰,这显示了生物生产率逐渐地有小幅度降低,但起伏较小。
瓮安磷矿上磷矿段可以分为上下两个层位,下部的黑色碳质磷块岩层与其上的灰白色白云质磷块岩层,分别对应于深水低能的局限环境和浅水高能的开放环境。实验获得的δ13Corg值均来自于黑色碳质磷块岩层,是整个瓮安生物群化石产出最丰富的层位。陡山沱中-晚期,瓮安由于受海侵作用而位于陆表海盆地中部[30],这种极低的δ13Corg值可以用Lewan提出的海洋模式来解释[26,27]:在浅水限流的陆表海盆地,有机质被微生物降解产生的CO2成为光合作用的碳源;又由于海水循环在此沉积环境中受限制,导致生物降解产生的富12C的碳源不断在陆表海盆地增加,当被瓮安生物群中大量的浮游生物在光合作用下利用时,产生的有机质将富集12C,最后其有机碳具有低的δ13Corg值(图5)。
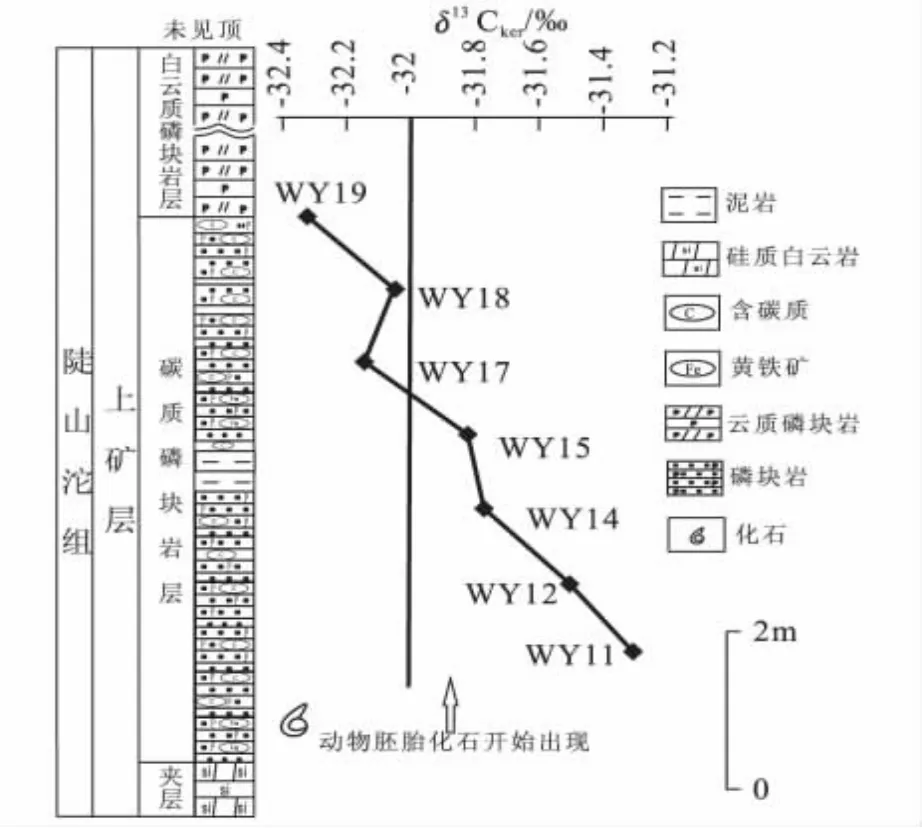
图4 瓮安穿岩洞剖面陡山沱组有机质碳同位素演变曲线Fig.4 Variation curves of the organic carbon isotope from Doushantuo Formation of the Chuan-Yandong section in Weng'an
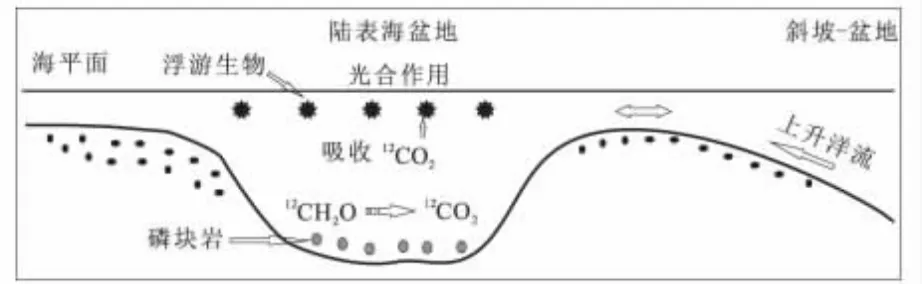
图5 瓮安陡山沱组L型无定形有机质成因Fig.5 The genesis of L-type amorphous organic matter from the Doushantuo Formation of Weng'an
此外,Dean通过实验发现在藻类光合作用过程中,CO2含量对其碳同位素分馏具有重要的分馏作用[31],高含量的CO2将导致同位素分馏系数的明显增大,使海相有机质相对于海水溶解碳更富集12C。Hoffman等认为“雪球地球”后[22],大气CO2含量很高,导致温室效应,因此,藻类进行光合作用过程中对CO2的分馏系数较大,海相有机质相对于海水溶解碳更富集12C,使有机碳同位素值明显偏低。
δ13Corg值的逐渐降低,可能与该期海平面变化有关。上矿层底部为沉积间断面,代表海平面下降时暴露出水面遭受侵蚀;向上为碳质磷块岩沉积物,代表开始又一次海侵。此时,瓮安生物开始大量产出,δ13Corg值维持在-31‰,海水深度不大,透光带内生物生产率较高;随后,δ13Corg值逐渐减小,反映海平面处于上升阶段,最低值达到-32.32‰,代表海水深度较大。由于海平面上升,下部水体的生物的光合作用受到限制,底部海水缺氧状态加剧,生物生产率下降导致δ13Corg值降低。
此外,宜昌陡山沱组第2岩性段(页岩与白云岩互层段)可以与瓮安磷块岩下矿层进行对比,获得的δ13Corg值为-28.97‰,反映了较高的生产率水平,与该层燧石结核中发现的疑源类、球状和丝状蓝菌、多细胞藻类相吻合。在陡山沱组顶部第4岩性段中的黑色页岩获得的δ13Corg值呈现极低的数值,为-38.39‰,显示了在陡山沱末期生物生产率的下降。
4 讨论
综合许多学者对新元古代末到早寒武世碳酸盐岩的δ13C变化趋势的研究,发现有相似的变化趋势,碳同位素表现为正偏移,但有几次负偏移[8,32]。获得的总体趋势与本研究相一致:在冰期结束后的初期,即陡山沱早期δ13C值较低,而在陡山沱中-后期表现为明显的正偏移;从长期趋势看,一直到前寒武纪/寒武纪界线附近,总体表现为正偏移。与此对应(图6),全球开始了大规模的磷块岩沉积,沉积的磷矿分布在除南极洲外几乎所有的大陆,生物圈也发生了巨大变化。磷矿的沉积与δ13C值正漂移有一定的相关性。
Tucker认为磷块岩的沉积一般与高的生物生产率和大洋海水的流通性相对应[33]。冰期后,气候转暖,陡山沱期开始大规模的海侵,导致浅海台地面积剧增,为浅海生物的繁盛及磷块岩的沉积提供了场所。同时,物理及化学风化作用使得陆源磷质输入海中,海水营养水平得以提高,为生物圈提供充足的供应。海洋也从滞留的封闭体系逐渐向开放体系演化,大洋环流作用增强,有利于上升洋流的发育[35],提供了携带富磷海水运移到扩大了的浅海台地附近的动力。另一个关于海水流通性提升的证据来自于硫同位素。Cook对该时期的蒸发岩硫酸盐及磷块岩中晶格硫酸盐的δ34S值的大幅度升高现象研究后发现,δ34S值的偏移是由深部洋水和浅部洋水的混合导致的[36]。深部洋水中厌氧的硫酸盐还原细菌的作用使得δ34S值的偏移并富集磷;而浅水地区相反,当海水流动性加大时,使得深部与浅部水体混合。

图6 埃迪卡拉纪-早寒武世的碳同位素和磷块岩丰度演化曲线略图[33,34]Fig.6 Generalized trend of C isotope and the abundance of phosphorites in Ediacaran-Early Cambrian carbonate rocks
此外,一般认为大洋缺氧事件发生时,会导致海相碳酸盐岩碳同位素δ13C值正偏移。溶解氧含量低的缺氧时期,生物死亡后的躯体得以保存,有机质(富12C)大规模埋藏势必造成海水中相对富13C[21]。Hiroto等发现前寒武纪末存在广泛缺氧的浅海环境[37];Donnelly等也认为,世界各地新元古代末海相碳酸盐岩δ13C值正偏移是由于大洋缺氧事件造成的[38]。含磷有机质在沉降过程中伴随着磷质释放,而缺氧条件下释放效率比氧化条件下高很多[39],因此海洋缺氧事件促使缺氧水体内磷的聚集,为磷块岩的最终沉积提供了条件。
在对古海洋的生物圈及生物演化的作用方面,磷可通过海水溶液方式对生态系统提供支持。对于匮乏磷的生态系统,其生物群及生产率受到严重制约。如现代的太平洋中的Kauai火山海岛,即是匮乏磷的生态系统,只能靠风力传输含磷灰尘来维持[40]。而在目前上升洋流活跃且有磷矿沉积的秘鲁、智利及纳米比亚外海,都具有高的有机质生产率,这必然与海水中的磷含量的增高有关。因此,陡山沱期的以瓮安生物群为代表的生物圈繁盛(如后生动物的出现)、生物有机质的成磷作用均是对古海洋环境重大变化的响应。
在海水的流通性提升后,经洋流上涌而影响磷的分布,进而影响生物圈这一系列反应后,促使了生物圈内部出现生命创新事件。因此,磷可能为这一事件的生物进化提供了动力。由于生物种类及数量的增加,可能在空间的利用、牧食及防御等方面产生压力及竞争[36,41];或者如Stanley的“收成原理”认为的,掠食性的生物以微生物及原始藻类为食,导致后者大量繁衍以避免灭绝,同时推动捕食者也随之进化[42]。
5 结论
瓮安陡山沱组大塘剖面的碳酸盐岩碳同位素特征与其他地区同期碳同位素变化趋势相似,显示了冰期后δ13C值正飘移的趋势,这与冰期后生物生产率的提高有关。瓮安穿岩洞剖面上磷矿段的黑色碳质磷块岩中的L型无定形有机质的低δ13Corg值是由于限流盆地中积累的12C源被浮游生物吸收导致,从另一个方面体现了生物有机质聚磷作用。大洋中的缺氧事件使底部海水富磷质,随着古海洋流通性的提高,海水中磷质被上升洋流带到浅海并沉积成矿。而由于有了磷质的供应,生物圈也开始繁盛,并引起了生物的进化。
[1]张同钢,储雪蕾,陈孟莪,等.新元古代全球冰川事件对早期生物演化的影响[J].地学前缘,2002,9(3):49-54.
[2]Xiao S,Zhang Y,Knoll A H.Three-demensional preservation of algae and animal embryos in a Neoproterozoic phosphorite[J].Nature,1998,391:553-558.
[3]黄思静.上扬子地台区晚古生代海相碳酸盐岩的碳、锶同位素研究[J].地质学报,1997,71(1):45-53.
[4]蒋干清,张世红,史晓颖,等.华南埃迪卡拉纪陡山沱盆地氧化界面的迁移与碳同位素异常[J].中国科学:D 辑,2008,38(12):1481-1495.
[5]Zhu M Y,Zhang J M,Yang A H.Integrated Ediacaran(Sinian)chronostratigraphy of South China[J].Palaeogeography Palaeoclimatology Palaeoecology,2007,254:7-61.
[6]王新强,史晓颖.华南伊迪卡拉纪碳同位素时空变化及其对生物演化的影响[J].中国科学:D辑,2010,40(1):18-27.
[7]Fike D A,Grotzinger J P,Pratt L M,et al.Oxidation of the Ediacaran Ocean[J].Nature,2006,444:744-747.
[8]Derry L A,Kaufman A J,Jacobsen S B.Sedimentary cycling and environmental change in the Late Proterozoic:Evidence from stable and radiogenic isotopes[J].Geochimica et Cosmochimica Acta,1992,56:1317-1329.
[9]Derry L A,Keto L,Jacobsen S,et a1.Sr isotopic variations in Upper Proterozoic carbonates from Svalbard and East Greenland[J].Geochimica et Cosmochimica Acta,1989,53:2331-2339.
[10]黄思静,石和,毛晓东,等.早古生代海相碳酸盐的成岩蚀变性及其对海水信息的保存性[J].成都理工大学学报:自然科学版,2003,30(1):9-17.
[11]Kaufman A J,Knoll A H,Awramik S M.Biostratigraphic and chemostratigraphic correlation of Neoproterozoic sedimentary successions:Upper Tindir Group,northwestern Canada,as a test case[J].Geology,1992,20:181-185.
[12]Kaufman A J,Knoll A H.Neoproterozoic variations in the C isotope composition of seawater:stratigraphic and biogeochemical implications[J].Precambrian Research,1995,73:27-49.
[13]Kaufman A J,Konll A H,Narbonne G M.Isotopes,ice ages and terminal Proterozoic earth history[J].Preceedings of the National Academy of Sciences,1997,95:6600-6605.
[14]Derry L A,Brasier M D,Corfield R M,et al.Sr and C isotopic in Lower Cambrian carbonates from the Siberian craton:A paleoenvironmental record during the Cambrian explosion[J].Earth and Planetary Science Letters,1994,128:671-681.
[15]Zhang T G,Chu X L,Zhang Q R,et al.Variations of sulfur and carbon isotopes in seawater during the Doushantuo Stage in late Neoproterozoic[J].Chinese Science Bulletin,2003,48(13):1375-1380.
[16]Bathurst R G.Carbonate Sediments and Their Diagenesis[M].Amsterdam:Elsevier,1975:1-439.
[17]Strauss H,Des Marais D J,Hayes J M,et al.Proterozoic organic carbon-its preservation and isotopic record[C]//In Early Organic Evolution:Implication for Mineral and Energy Resources.Berlin:Springer Verlag,1992:203-211.
[18]Hayes J M,Poppa B N,Takigiku R,et al.An isotopic study of biogeochemical relationships between carbonates and organic carbon in the Greenhorn For-mation[J].Geochimica et Cosmochimica Acta,1989,53:2961-2972.
[19]Delaney M L.Extinctions and carbon cycling[J].Nature,1989,337:18-19.
[20]周传明.贵州瓮安地区上震旦统碳同位素特征[J].地层学杂志,1997,21(2):124-129.
[21]胡修棉,王成善,李祥辉.大洋缺氧事件的碳稳定同位素响应[J].成都理工学院学报,2001,28(1):1-4.
[22]Hoffman P F,Kaufman A J,Halverson G P,et al.A Neoproterozoic Snowball Earth[J].Science,1998,281:1342-1344.
[23]Condon D J,Prave A R.Two from Donegal Neoproterozoic glacialepisodes on the northeast margin of Laurentia[J].Geology,2000,28(10):951 -954.
[24]Wang T G,Li M J,Wang C J,et al.Organic molecular evidence in the Late Neoproterozoic Tillites for a palaeo-oceanic environment during the snowball Earth era in the Yangtze region,southern China[J].Precambrian Research,2008,162:317-326.
[25]Lambert I B,Walter M R,Zang W L,et al.Paleoenvironment and carbon isotope stratigraphy of Upper Proterozoic carbonates of the Yangtze Platform[J].Nature,1987,325:140-142.
[26]Lewan M D.Stable carbon isotopes of amorphous kerogen from Phanerozoic sedimentary rocks[J].Geochimica et Cosmochimica Acta,1986,50:1583-1591.
[27]李任伟,卢家烂,张淑坤,等.震旦纪和早寒武世黑色页岩有机碳同位素组成[J].中国科学:D辑,1999,29(4):351-357.
[28]Cao C,Wang W,Jin Y,et al.Carbon isotope excursions across the Permian-Triassic boundary in the Meishan section,Zhejiang Province,China[J].Chinese Science Bulletin,2002,47(13):1125-1129.
[29]陈兰,钟宏,胡瑞忠.湘黔地区早寒武世黑色页岩有机碳同位素组成变化及其意义[J].矿物岩石,2006,26(1):81-85.
[30]周传明,薛耀松,张俊明.贵州瓮安磷矿上震旦统陡山沱组地层和沉积环境[J].地层学杂志,1998,22(4):308-313.
[31]Dean W E,Arthur M A,Claypool G E.Depletion of13C in Cretaceous marine organic matter:source,diagenetic,or environmental signal?[J].Marine Geology,1986,70:119-157.
[32]Knoll A H,Walter M R.Latest Proterozoic stratigraphy and earth history[J].Nature,1992,356:673-678.
[33]Tucker M E.The Precambrian-Cambrian boundary:seawater chemistry,ocean circulation and nutrient supply in metazoan evolution,extinction and biomineralization[J].Journal of the Geological Society,1992,149:655-668.
[34]Cook P J.Phosphogenesis around the Proterozoic-Phanerozoic transition[J].Journal of the Geological Society,1992,149:615-620.
[35]Kaufman A J,Jacobsen S B,Knoll A H.The Vendian record of Sr and C isotopic variations in seawater:implications for tectonics and paleoclimate[J].Earth and Planetary Science Letters,1993,120:409-430.
[36]Cook P J,Shergold J H.Phosphorus,Phosphorites,and Skeletal Evolution at the Precambrian-Cambrian Boundary[J].Nature,1984,308:231-236.
[37]Hiroto K,Yoshio W.Oceanic anoxia at the Precambrian-Cambrian boundary[J].Geology,2001,29(11):995-998.
[38]Donnelly T H,Shergold J H,Southgate P N,et al.Events leading to global phosphogenesis around the Proterozoic Cambrian boundary[C]// Phosphorite Research and Development.London:Geological Society Special Publication,1990,52:273-287.
[39]Ingall E D,Jahnke R.Influence of water-column anoxia on the elemental fractionation of carbon and phosphorus during sediment diagenesis[J].Marine Geology,1997,139:219-229.
[40]Kurtz A C,Derry L A,Chadwick O A.Accretion of Asian dust to Hawaiian soils:Isotopic,elemental,and mineral mass balances[J].Geochimica et Cosmochimica Acta,2001,65:1971-1983.
[41]袁训来,肖书海,周传明.新元古代陡山沱期真核生物的辐射[C]//生物的起源、辐射与多样性演变——华夏化石记录的启示.北京:科学出版社,2006:13-27.
[42]Stanley S M.An ecological theory for the sudden origin of multicellular life in the late Precambrian[J].PANS,1973,70(5):1486-1489.
猜你喜欢
化工矿产地质(2022年2期)2022-07-21 03:17:14
科技与创新(2020年15期)2020-08-12 06:01:42
沉积与特提斯地质(2019年1期)2019-07-16 08:41:12
当代贵州(2017年45期)2017-11-16 08:38:51
乡村地理(2017年4期)2017-09-18 02:53:58
当代贵州(2017年28期)2017-08-01 00:17:51
小康(2014年15期)2014-12-15 02:22:30
陕西学前师范学院学报(2014年5期)2014-07-31 18:12:12
紫光阁(2014年3期)2014-03-07 17:09:31
中国矿业(2013年1期)2013-11-20 05:05:20
