
台湾林檎(Malus doumeri(Bois.)Chev.)地理分布模拟1)
2012-07-02 00:03王雷宏杨俊仙郑玉红汤庚国
东北林业大学学报 2012年9期
王雷宏 杨俊仙 郑玉红 汤庚国
(安徽农业大学,合肥,230036)(江苏省中国科学院植物研究所)(南京林业大学)
苹果属(Malus Mill.)多胜海棠组(Section Docyniopsis)是属内一个独立的进化分支,表现出了独特的果心分离性状,是第三纪孑遗种,在东亚地区是最原始的类群,最新分类学研究认为本组只有乔劳斯基海棠(Malus tschonoskii(Maxim.)Schneid.)、台湾林檎(Malus doumeri(Bois)Chev.)2个种,前者分布在日本富士山,后者分布较广,其下包括原变种(Malus doumeri var.doumeri)、尖嘴林檎 (Malus doumeri var.melliana)、光萼林檎(Malus doumeri var.leiocalyca)[1-3]。现已发现的苹果属植物化石都在第三纪中新世之前[4-7],这些化石与现存苹果属较原始类群多胜海棠组植物亲缘关系仍存在较大争议[2,8],而且现存苹果属属下的组级类群间也没有直接的亲缘谱系关系,也就是说现存最原始的类群绝不是本属现存植物的祖先种[8]。在苹果属植物化石资料仍不能提供充足的证据条件下,研究苹果属较原始类群的分布格局对于揭示本属植物的起源和形成仍具有重要的科学意义,至少能揭示第四纪气候变化以来现存部分苹果属植物的地理分布和演化规律,然现有的标本记录数量有限,不能完整地反映台湾林檎的地理分布格局,其分布与生物气候因子之间的响应关系仍不明确[9]。笔者试图用生态位模型(BIOCLIM)模拟台湾林檎地理分布格局,分析影响其地理分布的生物气候主导因子,依据其生态学特性和分布规律在一定程度上试推测其地理分布形成的可能原因,为今后该植物的多样性起源以及种质资源利用提供基础资料。
1 材料与方法
1.1 分布数据来源和处理
以苹果属分类学研究的最新成果为依据[2],重新鉴定中国科学院植物研究所标本馆(PE)、中国科学院武汉植物园标本馆(HIB)、中国科学院昆明植物标本馆(KUN)、江苏省中国科学院植物研究所标本馆(NAS)、南京林业大学标本馆(NF)、南京大学标本馆(N)、中国科学院成都生物研究所植物标本室(CDBI)、西南农业大学植物标本馆(HWA)、西南林业大学植物标本室(SWFC)的标本,共获得台湾林檎有价值的分布记录243个,其中原变种105个,尖嘴林檎84个,光萼林檎54个,查阅中国科学院植物研究所标本馆地名数据库(http://beta.cvh.org.cn/cms/cn/node/42)获得分布记录地点的经纬度坐标,作为模拟分析的基础数据。
1.2 气象栅格数据和地理矢量数据
采用当前和未来气候环境数据包括19个环境因子变量:bio1年均温、bio2平均日温差、bio3日均温差(bio2除以年均温差bio7)、bio4温度季节变化方差、bio5最热月温度、bio6最冷月温度、bio7温变年均差、bio8雨季均温、bio9干季均温、bio10暖季均温、bio11冷季均温、bio12年降雨量、bio13最湿月降雨量、bio14最干月降雨量、bio15季降雨量变异系数、bio16雨季降水量、bio17干季降水量、bio18暖季降雨量、bio19冷季降雨量,空间分辨率为30 arc s(相当于1 km空间分辨率),均来自WORLDCLIM[10](http://www.worldclim.org/)。从 http://nfgis.nsdi.gov.cn/,http://www.diva-gis.org 分别下载 1 ∶4 000 000的中国、亚洲矢量地图作为分析底图。
1.3 研究方法
采用 DIVA-GIS(http://www.diva-gis.org)软件,BIOCLIM 模型在该软件中自动实现[11-13]。显示预测图时,只涉及到少量的一些周边国家领土,领土所属和小岛屿的标注不属于本次研究范畴,未细化。模拟种级单位分类群时,把所有分布数据当作一个分类群,比较变种间的生态位间隔时,在软件设置面板上选择种下分类群,其余参数设置一律采用默认值。主成分分析采用SPSS 15.0。
2 结果与分析
2.1 标本记录分布和当前气候模拟分布
依据标本记录绘制的分布格局,简单显示本种分布在越南中部,中国西南、华南、华东以及台湾,表明台湾林檎分布北界已达江苏南京地区,最西在云南省新平县,最南达到越南昆蒿,最东在我国台湾岛。中国西南地区分布数较少,以浙江省、江西省、福建省、广东省分布的数量为多,然而这种分布格局与各地区标本的采集强度和频度是密切相关的,甚至一些标本可能不是原采地,反映的情况可能不全面。分布式样基本格局主要呈西南—华南—华东连续分布以及与台湾的间断分布。
用生态位模型(BIOCLIM)模拟地理分布,可以更一步客观显示各分布地区的分布密度和分布格局,用颜色深度(灰色到黑色)表示各分布区的分布值大小,轮廓显示分布的区域。结果显示,模拟分布与标本记录分布基本情况是吻合的,也出现少量的标本记录点与模拟分布区域不吻合,如四川省标本分布在成都,然模拟显示主要分布在宜宾到泸州的长江段;标本记录江苏南京钟山有分布,然模拟显示在江阴市长江段区域有分布。没有标本记录,但有模拟分布值的地区有:越南的北部;海南岛中南部;重庆的方斗山、七曜山;湖北省的西南部、七曜山的东坡,宜都市周边的青龙山、宋山,幕阜山的北坡,大别山西坡;安徽省九华山;韩国的通济岛及附近岛屿;日本岛的中南部,这些模拟分布区域都可视为潜在分布区,见图1。
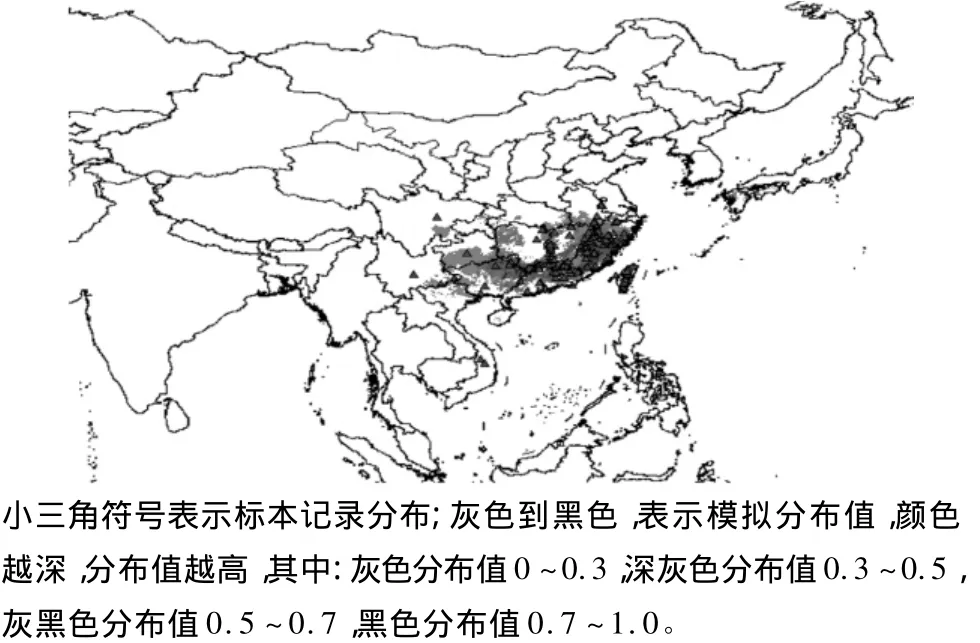
图1 台湾林檎的标本记录分布和模拟分布
2.2 分布与环境变量的关系
BIOCLIM模型中采用环境包络运算法则进行模拟。利用落在环境因子变量包络内的有效分布点作为分析对象,对其所在单元格环境变量做主成分分析,确定影响分布的主导因子。本次有效点共152个,19个主成分中的前3个主成分累计方差百分比达到了87.282 3%,能反映所有有效分布点生物气候变量信息量的87.282 3%(表1)。

表1 前3个主成分总方差解释
从表2可以看出:第1主成分主要由雨季降雨量(bio16)反映,第2主成分主要由年均温(bio1)反映,第3主成分主要由最干月降雨量(bio14)反映。忽略生物气候变量间的相互作用,从生物气候变量的边际效应来看,可认为雨季降雨量,年均温,最干月降雨量是影响其分布的主导因子。

表2 相对于主成分各变量的权重系数
2.3 种下单位间的生态位间隔
尖嘴林檎、光萼林檎与原变种在形态上极为相似,生态位也可能相似,比较它们之间19个生物气候因子生态位,发现仅分析得出的3个主导因子生态位有明显的间隔,其它生物气候因子生态位明显交错重叠。如台湾林檎原变种和尖嘴林檎雨季降水量生态位有间隔(图2),以累积分布频数60%以上区别更为明显。

图2 台湾林檎和尖嘴林檎的累积分布频数与雨季降雨量的关系
2.4 分布区响应气候变化
对整个亚洲地区用BIOCLIM模型预测未来分布格局和潜在分布,见图3,分布值仍用灰色到黑色表示,与图1相同。比较当前和未来分布格局、潜在分布区的变化,发现整个分布范围的界限几乎无变化,然当前分布区内的各级分布区域普遍向北扩展程度较大,分布值加大,分布连续性加强,如,湖南省北部明显向北扩展,与湖北交界处壶瓶山地区分布值达到0.7~1.0,浙江北部的天目山,东部天台山分布值加大到0.7~1.0。以上分布区变化规律暗示,未来气候条件下,台湾林檎的分布区范围不会发生明显的南北或东西向迁移,也即气候变化对其分布范围影响不大。具体见图3。
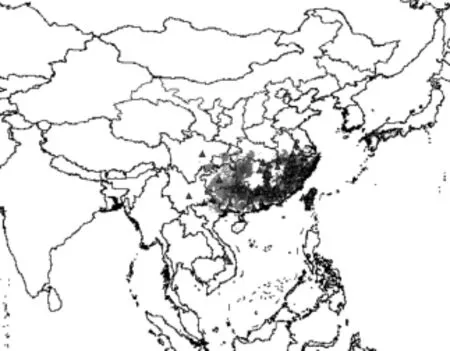
图3 台湾林檎未来分布格局和潜在分布
3 结论与讨论
3.1 台湾林檎地理分布
从标本记录和模拟分布区块的吻合程度来看,越南,中国的湖南省、贵州省、云南省、四川省、重庆市、湖北省的分布面积较少,主要分布在浙江省、福建省、江西省、广东省、广西省。江苏省的标本记录和模拟分布块也不吻合,江苏省中国科学院中山植物所后山的标本,可能是从安徽的黄山和浙江天目山等地引入栽培。运用GIS技术绘制物种的地理分布图,然后基于这种矢量数据,借用全球气候数据,采用生态位模型研究物种的地理分布变化,是植物地理演化研究的一个新方法、新途径,可操作性、科学性、准确性都较高[3]。传统植物区系学绘制植物分布图只是利用某几点的分布而示意了某个地区的存在,连线图也极易扩大植物的实际分布范围,不利于进一步深入分析。
3.2 台湾林檎地理分布与生物气候的关系
用BIOCLIM模型研究地理分布以及与生物气候因子的关系,可以从宏观上阐明物种分布生态或地理演化的规律。研究发现雨季降雨量、年均温、最干月降雨量是影响台湾林檎地理分布的主导因子,而且发现变种间大部分气候因子的生态位并不严格相隔,只有3个主导因子生态位有一定的间隔,可知种下单位外部形态特征应与气候因子的环境压力关系不大,偏向于物种居群内的基因型。在具体理解生物气候因子的生态位上有一定难度,实际上各生物气候因子间有内在的联系,理解主导因子时,可以理解为在其它生物气候因子在一定波动水平时,主导因子的略微变化对物种的地理分布影响程度较大。用BILCLIM模型建模时,采用了当前全球生物气候栅格数据(分辨率30 arc s),准确度较高。未采用中国地面气候数据标准值生成栅格数据,主要原因在于:一方面全球生物气候数据精确度已较高,它本身包含了我国气象台站的数据;另一方面,台湾林檎有一些分布点邻近其他国家(老挝,越南等),如采用中国的气候数据,就无法准确模拟台湾林檎的当前和未来的分布格局和潜在分布。
3.3 台湾林檎的潜在分布
从当前气候下的潜在分布结果来看,中国海南岛、日本琉球岛、韩国南部及其一些小岛屿都可能有分布,这些地区分布值主要是0.3~0.5,极少数地方达到了 0.5 ~ 0.7,然从 GBIF(http://data.gbif.org)查找记录信息,韩国、日本未有记录,海南岛也没有分布记录,这些地区仍不存在台湾林檎。日本出现了另一个地理替代种乔劳斯基海棠。从形态来看,二者的区别主要在于叶的质地、叶缘裂与不裂及锯齿类型,ITS序列系统发育分析也表明二者亲缘关系密切[4]。据此,可推测乔劳斯基海棠和台湾林檎是由它们的共同祖先地理隔离形成的新类群。总体来看,本次分布模拟较为准确,所模拟的当前分布应该接近了台湾林檎真实地理分布,结合未来分布图的变化特征反映了其基本的生物气候生态需求,未来大气CO2浓度增高时,引起的生物气候变化对本种的地理分布范围影响不大,只是分布区域内各级适生区范围都扩展和分布值升高。
3.4 台湾林檎的起源
依据台湾林檎地理分布格局和对气候变化的响应特征,可简单推测它的起源和迁移路线,首先这一物种是东亚第三纪孑遗种,而苹果属植物化石显示到中新世未时较台湾林檎更原始的植物广布欧、亚、美洲,显然台湾林檎是在上新世形成的,考虑到进化的时间,最晚在上新世末更新世初,也就是距今200万年前就形成了,按照它的生态习性,它在亚洲起源只能是年均温度高,雨季多雨的地方,而且上新世以来气温的下降变化对这一地方气候影响不大,而且本种群极易在水平方向扩张,所以在现亚洲大陆亚热带南缘和热带北缘的区域起源可能性较大,而且在这个时期未受到反复海侵作用的高海拔或者海拔上升较快的地方[14],推测起源地就在云南省、广西、老挝和越南交界处。依据最新中国台湾、日本琉球岛构造理论,可知2岛从更新世到全新世冰期前仍在海底[15],所以这2岛台湾林檎及地理替代种的起源,当是在第四纪冰期时,海水下沉后,它们的祖先种通过大陆架桥进行的扩展。总体来看祖先起源后,是向东进行扩展,可能是第四纪未次大冰期的气候条件奠定了基本格局,最后演化成现有的2个种以及地理分布式样。
3.5 模拟分布的局限性
模拟物种地理分布影响因素较多:影响较大的因素仍然是标本鉴定的正确性和分布记录的准确性以及标本采集方式是不是近似于随机采样[16-17];其次由薄盘光滑样条函数插值法所得来的高分辨率生物气候变量数据也难以保证与复杂地形区域的气候完全吻合[9],未来CO2浓度升高后,气候条件改变,分析时假设了景观背景上各种土地利用类型以及沿海陆地未发生变化。
总之,基于分布与环境生态位理论的BIOCLIM模型简单实用,模拟结果较为保守谨慎,模拟出的分布范围界限与标本记录分布较为吻合,有利于研究植物和环境的关系,从软件中可以方便提取有效分布点的生物气候变量数据,利用这些数据可进行主成分分析,有利于从生态位理论的角度揭示影响物种地理分布的主导气候因子,提示物种气候尺度的基本生态需求。
[1]谷粹芝,李朝銮.台湾林檎及其近缘种的研究[J].广西植物,1990,10(1):11-14.
[2]钱关泽.苹果属(Malus Mill.)分类学研究[D].南京:南京林业大学,2005.
[3]王雷宏,杨俊仙,郑玉红,等.苹果属山荆子地理分布模拟[J].北京林业大学学报,2011,33(3):70-74.
[4]Robinson J P,Harris S A,Juniper B E.Taxonomy of the genus Malus Mill.(Rosaceae)with emphasis on the cultivated apple,Malus domestica Borkh.[J].Plant Systematics and Evolution,2001,226(1/2):35-58.
[5]Hu H H,Chancy R W.A miocene flora from Shantung Province,China[M].Washington:Carnegie Institution of Washington Publication,1940:10,17,47,48,124,127,135.
[6]中国科学院植物研究所南京地质占生物研究所《中国新生代植物》编写组.中国植物化石:中国新生代化石[M].北京:科学出版社,1978:98-99.
[7]Britton N L,Brown A.An illustrated flora of the Northern United States and Canada:Vol.2[M].2 ed.Newyork:Dover Publications,1913:234-236.
[8]王雷宏,汤庚国,夏海武,等.山荆子叶脉序的研究[J].南京林业大学学报:自然科学版,2008,32(2):39-42.
[9]邓毓芳,刘显旋,钟海雁,等.林擒生物学特性及开发利用前景的研究[J].中南林学院学报:1996,16(4):24-27.
[10]Hijmans R J,Cameron S E,Parra J L,et al.Very high resolution interpolated climate surfaces for global land areas[J].International Journal of Climatology,2005,25:1965-1978.
[11]Hijmans R J,Spooner D M.Geographic distribution of wild potato species[J].American Journal of Botany,2001,88(11):2101-2112.
[12]Jarvis A,Ferguson M E,Williams D E,et al.Biogeography of wild Arachis:assessing conservation status and setting future priorities[J].Crop Science,2003,43(3):1100-1108.
[13]Jarvis A,Guarino L,Williams D,et al.Spatial analysis of wild peanut distributions and the implications for plant genetic resources conservation[J].Plant Genetic Resources Newsletter,2002,131:28-34.
[14]黎先耀,刘思孔.生物史图说[M].北京:科学出版社,1990.
[15]戴昌凤.台湾地区生物礁及其生境[J].古地理学报,2010,12(5):565-576.
[16]王运生,谢丙炎,万方浩,等.ROC曲线分析在评价入侵物种分布模型中的应用[J].生物多样性,2007,15(4):365-372.
[17]王利松,陈彬,纪力强,等.生物多样性信息学研究进展[J].生物多样性,2010,18(5):429-443.
猜你喜欢
数学小灵通·3-4年级(2021年6期)2021-07-16
少儿科学周刊·儿童版(2018年12期)2018-01-26
少儿科学周刊·少年版(2018年12期)2018-01-26
少儿科学周刊·儿童版(2017年7期)2017-09-29
江苏农业科学(2017年5期)2017-04-15
科技与创新(2016年18期)2016-11-04
湖南水利水电(2015年2期)2015-12-24
湖北农业科学(2014年21期)2014-12-22
新疆农垦科技(2014年7期)2014-02-28
自然资源遥感(2014年3期)2014-02-27
