
接种AM真菌对K326烟苗素质的影响
2012-06-08 07:54杨红武裴晓东龙世平
湖南农业科学 2012年19期
李 帆,陈 坤 ,杨红武 ,裴晓东 ,龙世平
(1.长沙市烟草公司浏阳分公司,湖南 浏阳 410300;2.湖南省农业生物资源利用研究所,湖南 长沙 410125)
国内外有学者研究表明,烟草对菌根有较强的依赖性,它能与某些特定AM真菌共生形成菌根,烟草接种AM真菌后对其增加养分的吸收、增强其抗逆性等方面有较为明显的作用[1-3]。研究表明烟草能否成功接种AM真菌,一方面,土壤中氮、磷的养分含量是重要的影响因子[4-6],同时其受生态条件的影响也比较大;另一方面,宿主植物与AM真菌存在着相互选择性[7-8]。因此,在开展规模化菌根技术育苗之前,为了更加深入了解烟草接种AM真菌后的菌根效应,先进行烟草幼苗接种AM真菌试验。笔者在前人[9]研究的基础上,选取适合浏阳生态条件的在烤烟生产上应用的2种AM真菌菌种,并在漂浮育苗阶段接种,研究了在不同基质肥力条件下其对烟株苗期生长与营养的影响,旨在为AM真菌在浏阳优质烤烟生产中规模化应用提供技术参考。
1 材料与方法
1.1 材料
供试烤烟品种为K326(Nicotiana tabacum L.CV K326),由浏阳市烟草公司提供。供试菌种为聚丛球囊霉(Glomuse aggregatum,BGC HUN02D)和摩西球囊霉(Glomusemosseae,BGC HUN01A),均由北京市农林科学院植物营养与资源环境研究所提供。烤烟育苗专用肥(氮150 g/kg,磷100 g/kg,钾180 g/kg)由浏阳市烟草公司提供。
1.2 方法
1.2.1 试验设计 共设12个处理,每处理重复3次,共36个小区,每小区1盘(200株烟苗)。基质A,V腐熟菌渣∶V泥炭∶V珍珠岩=3∶2∶1;基质 B,V腐熟芦苇渣∶V泥炭∶V珍珠岩=3∶2∶1;基质 C,V腐熟猪粪稻草∶V泥炭∶V珍珠岩=3∶2∶1;基质D为烟草育苗专用基质。接种情况见表1,后期管理同浏阳市烟草专卖局优质烤烟生产规程。
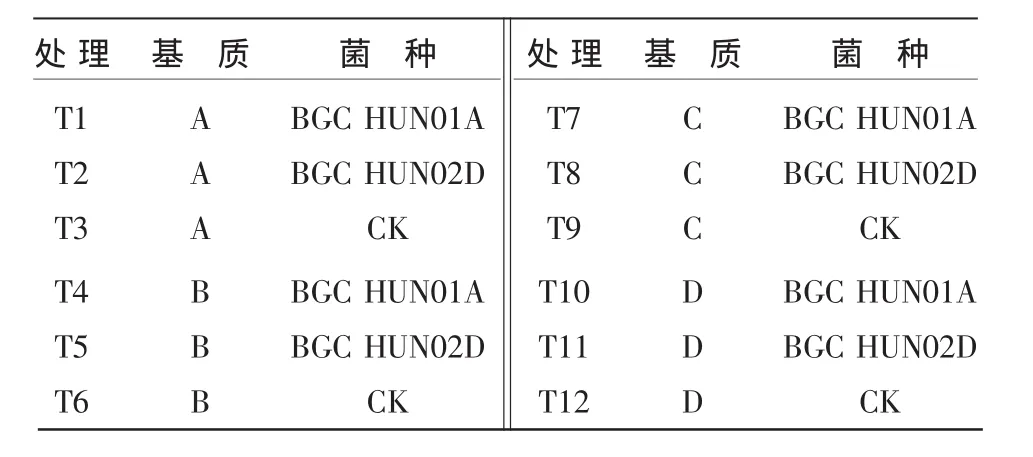
表1 各处理施肥及接种情况
1.2.2 播种、接种及管理 菌根烟苗的培育:在漂浮育苗条件下,播种期接种根内球囊霉(Glomus intraradices Smith&Schenck,BEG-141),培养液养分用量为1.2×Hoagland,由低到高,分次供应,成苗期镜检AM真菌侵染率≥25%。非菌根烟苗的培育:在漂浮育苗条件下,播种期接种等量灭活接种物,培养液养分用量为1.2×Hoagland,提供方式同菌根烟苗的培育,成苗期镜检未发现菌根结构。
播种期分别接种2种AM真菌,不接种者为对照(CK)。将基质灭菌(121℃,1 h),先加超纯化水润湿后,装入烤烟漂浮育苗的专用漂浮盘(10×20穴/盘)孔内,每穴5 g。然后将2 g接种物埋入0.3~0.5 cm深的基质中,形成接种处理,对照为埋入等量灭活接种物。每穴播种3粒烤烟种子,出苗后留1株。试验在人工控制温度、光照和湿度的培养室中进行,幼苗生长期间,漂浮育苗池的水位保持在8 cm左右。光照强度14 000~15 000 lx,每天光照时间为12 h。昼夜温度保持20~31℃,湿度维持70%~80%。
1.2.3 测定项目与方法 从出苗第10天起,每5 d每小区取10株烟苗称量植株鲜重。幼苗根系经0.1%酸性品红乳酸甘油染色液处理染色后,在显微镜下观测真菌结构,估算AM真菌侵染率。在成苗期(出苗后55 d),每小区取20株烤烟幼苗,测量根长、株高、茎粗。然后将根系附着的基质抖落,用自来水和去离子水依次清洗,随机切取不同部位的鲜根,用根段频率标准法测定根系侵染率[6,10]。剩余样品取地上部分,105℃杀青30min,70℃烘干后粉碎样品,参考鲍士旦[11]方法用蒸馏法、钒钼黄比色法和火焰光度法分别测定烟株的氮、磷、钾含量。基质待自然阴干后,测定磷酸酶活性[12]。
2 结果与分析
2.1 接种AM真菌对烤烟K326幼苗侵染的影响
烤烟种子萌发后20 d镜检成苗期的烟苗根系发现,接种AM真菌后,根系内均出现真菌结构,接种AM真菌处理可在根系上观测到菌丝的侵入点、丛枝形成,而对照未见。这表明接种聚丛球囊霉、摩西球囊霉两种AM真菌均能成功侵染烤烟K326幼苗根系,形成菌根烟苗。
由图1可以看出,A、B、D三种基质成苗后BGCHUN02D与BGCHUN01A两个菌种侵染率均无显著差异,而C基质BGC HUN02D侵染率显著高于BGCHUN01A;此外,A基质中两种AM真菌的侵染率均低于16%,且明显低于B、C、D基质AM真菌的侵染率(26.8%~36%),表明基质A更有利于AM真菌侵染寄主根系。基质A接种BGCHUN02D的侵染率低于BGC HUN01A,而基质B、C、D接种BGC HUN02D侵染率则高于接种BGC HUN01A。此外,镜检还发现,BGCHUN02D侵染根系后,根外菌丝较多,BGCHUN01A的根外菌丝较少。
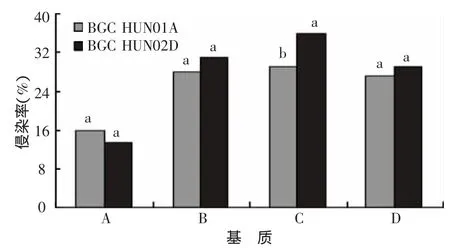
图1 不同施肥水平下两种AM真菌对烟株幼苗根系的侵染状况
2.2 接种AM真菌对K326幼苗根系的影响
不同基质接种两种AM真菌可不同程度地增加烟株的根长和根重,而对根冠比的影响增减不一。如表2所示,接种BGC HUN01A和BGC HUN02D均对烟株幼苗根系生长有促进作用,能达到壮苗的目的,且A、B、C基质中接种BGCHUN01A的烟株幼苗根重、根冠比都高于接种BGCHUN02D幼苗,表明BGC HUN01A比BGC HUN02D在促进K326烟苗根系生长方面具有优势。其中基质A接种BGC HUN01A的烟苗根重显著高于CK,但根冠比与CK差别不大。基质C接种BGC HUN01A的幼苗的根冠比、根重都明显高于CK,根冠比和CK相比增加57.4%,比接种BGC HUN02D幼苗高21.2%。另两种基质B、D接种供试两种AM真菌的K326幼苗的根系生长未见明显优势。由此可见,接种AM菌种对烟株幼苗的生长发育有促进作用。
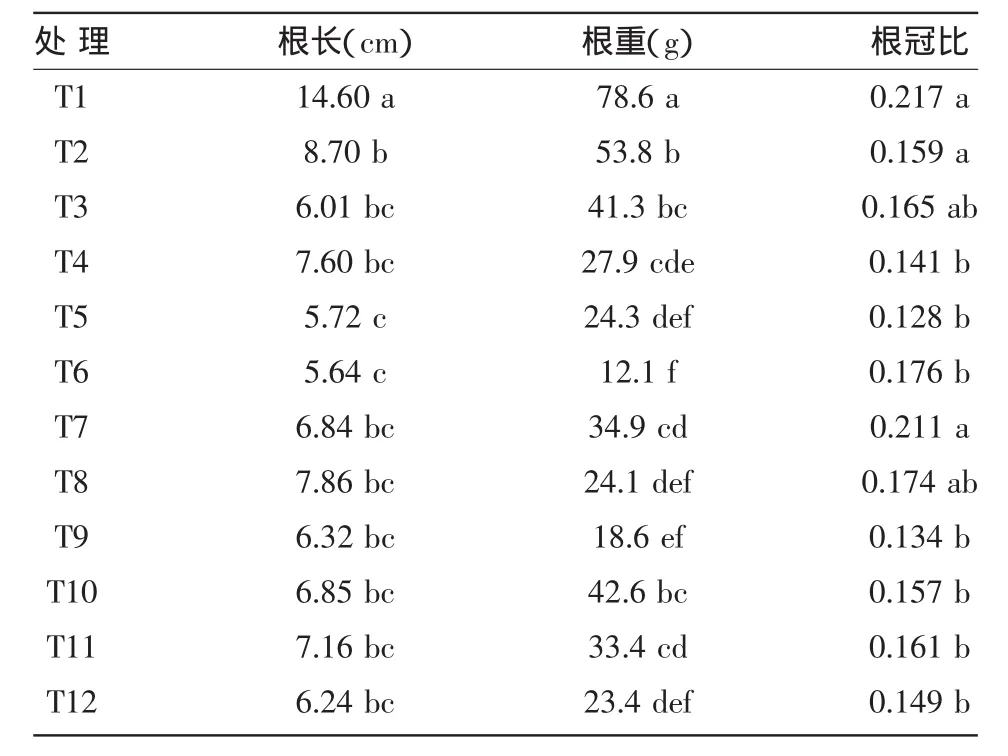
表2 接种AM真菌对烤烟幼苗根系的影响
2.3 接种AM真菌对K326幼苗养分含量的影响
如表3所示,和CK相比,K326幼苗接种两种AM真菌都能提高对氮的吸收,接种BGC HUN01A的K326幼苗含氮量比CK高13%,接种BGC HUN02D的菌根苗只比CK高0.03%,可见BGC HUN01A在增强K326幼苗吸氮能力上要比BGC HUN02D强。K326接种两种AM真菌后,K326菌根幼苗的含磷量都显著地提高了。这表明AM真菌能够促进烟苗对磷的吸收,增加烟株体内的磷含量,从而促进烟株的生长发育。接种两种AM真菌后烤烟幼苗之间吸磷能力存在差异,BGC HUN01A菌根烤烟幼苗的含磷量显著高于BGC HUN02D,说明接种BGC HUN01A的K326幼苗吸磷能力上优于BGC HUN02D。接种AM真菌能显著地提高K326幼苗的含钾量,接种BGC HUN01A的K326烟苗的含钾量显著高于BGC HUN02D,接种BGC HUN01A真菌的烤烟幼苗对钾的富集的作用,这对于优质烤烟生产意义重大。
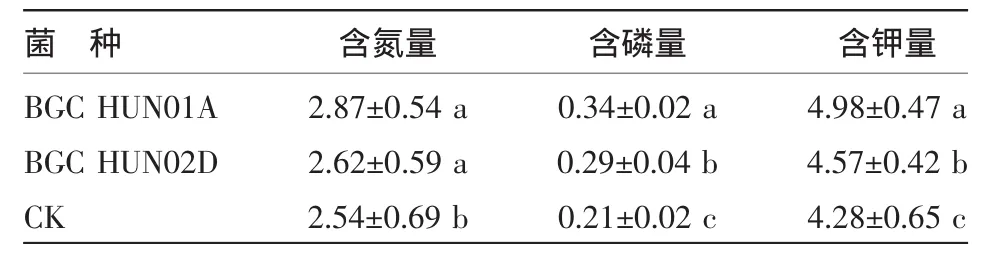
表3 接种菌种对烤烟幼苗氮、磷、钾含量的影响 (%)
2.4 AM真菌对基质磷酸酶活性的影响
如图2所示,接种不同AM真菌育苗基质中磷酸酶活性为:BGC HUN01A>BGC HUN02D>CK。4种接种BGCHUN01A的K326烟苗的基质,磷酸酶活性比对照处理分别高23.1%、29.5%、26.4%和30.1%。在4种施肥水平中,接种BGC HUN01A比接种BGC HUN02D的基质,其磷酸酶活性分别高10.4%、16%、10.9%和12.5%。由此可见,K326烟苗接种AM真菌显著提高了育苗基质中的磷酸酶活性,且不同菌株之间存在明显差异。两种AM真菌相比较,接种BGC HUN01A的烟苗基质能分泌出更多的磷酸酶。
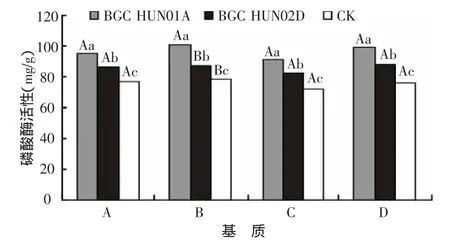
图2 接种AM真菌对基质磷酸酶含量的影响
3 讨论与结论
3.1 两种AM真菌对K326烟苗菌根侵染率的影响
AM真菌侵染率的高低和很多因素有关,其中介质养分的浓度、有效性是重要的影响因素,养分过低或过高都不利于AM真菌对烤烟的侵染。该试验表明,基质 B、C、D接种 BGC HUN01A和BGC HUN02D的侵染率均高于基质A,可能是因为基质A营养相对比较充足,导致K326烟苗对菌根真菌的侵染有排斥作用,高养分对AM真菌的侵染不利。
3.2 AM真菌对烤烟幼苗营养的影响
AM真菌的侵染是产生菌根效应的基础,菌丝进入烤烟幼苗根系后以不同方式影响植物的代谢过程。AM真菌的菌丝作用,扩大了植物根系对土壤中氮素的吸收面积和与土壤的接触位点,增强了植物的吸收利用。另外,AM真菌能将NH4+运给寄主植物,通过提高硝酸还原酶活性,有助于加强寄主植物氮代谢的有效性,增加菌根植物的生物量。烟草和其他作物一样,磷素临界营养期通常出现在苗期,所以烟草幼苗含磷量高对提高烤烟幼苗质量相当有利,能使烟苗移栽后迅速还苗,甚至能提高整个生育期的磷素营养。该研究证明,烤烟K326接种AM真菌能显著提高烤烟幼苗的磷素营养,对幼苗的生长有促进作用。这可能是因为AM真菌扩大了磷素的吸收领域,菌丝强大的吸收和运输能力以及AM真菌能够活化难溶性磷酸盐等。钾是优质烟叶生产中重要的化学元素。试验也表明,B、C、D基质接种AM真菌能显著地提高烟株幼苗的含钾量,而且接种BGC HUN01A的烟苗的含钾量显著高于BGCHUN02D,接种BGC HUN01A真菌的烤烟幼苗对钾的富集的作用,这对于优质烤烟生产意义重大。
3.3 AM真菌对基质磷酸酶活性的影响
接种不同AM真菌都能不同程度地影响K326烟苗的生长,B、C、D基质接种AM真菌都能增加基质酸性磷酸酶的活性。其主要原因是菌根菌丝能生长到根系不能到达的基质中吸收磷素,并运输给烟苗根系,以满足烤烟幼苗的营养需求。不同的菌根到达外界的范围不一样,所获得的养分空间也不一样,所以造成了吸收和运输给烟苗的磷素的差异。该研究中,K326接种BGC HUN01A的烟苗基质能分泌出更多的磷酸酶。
总体上讲,该研究表明,接种AM真菌可不同程度提高烟苗综合素质,而且接种BGC HUN01A的烟苗素质优于接种BGC HUN02D的烟苗。基质A幼苗根系生长相对旺盛,幼苗素质也相对较好,但其接种AM真菌后根系侵染率相对较低。
[1] Zhao L L,Li B,Zhao FG.Effects of VA mycorrhizal fungion the physiological and biochemical characters of tobacco[J].Acta A-gric.BorealiOccidentalis Sin,1999,8(6):136-138.
[2] 赵丽莉,李 斌,赵方贵.接种VA菌根真菌对烟草某些生理生化特性的影响[J].西北农业学报,1999,8(6):136-138.
[3] 江 龙,黄建国,袁 玲.不同丛枝菌根真菌对烟苗生长营养状况和生理活性的影响[J].贵州农业科学,2009,37(12):53-58.
[4] Koide R T,LiM.On host regulation of the vesicular—arbuscular mycorrhizal symbiosis[J].New Phytol,1990,114:59-96.
[5] Balaz M,Vosdtka M.Vesicular-arbuscularmycorrhiza of Calamagrostis villosa supplied with organic and inorganic phosphorus[J].Biol.Plantarum,1997,39(2):281-288.
[6] Liu R J,Luo X S.A new method to quantify the inoculum potential of arbuscular mycorrhizal fungi[J].New Phytologist,1994,128:89-92.
[7] 刘延荣,丁兆龙,方宇澄.烟草上优良丛枝菌根真菌的筛选[J].山东农业大学报,1997,28(3):269-273.
[8] 方宇澄,黄 镇,刘延荣.烟草VA菌根菌区系研究[J].中国烟草学报,2000,6(4):26-30.
[9] 李竹玖.AM真菌对烤烟幼苗生长和营养的影响[D].重庆:西南大学,2008.
[10] 靳冬梅,叶协峰,刘国顺,等.不同浓度营养液对烤烟漂浮育苗烟苗生长及生理特性的影响[J].华北农学报,2005,20(6):15-19.
[11] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2003.
[12] 关松荫.土壤酶及其研究法[M].北京:中国农业出版社,1986.
猜你喜欢
江西农业(2022年2期)2022-02-25
园林科技(2020年2期)2020-01-18
西南农业学报(2017年4期)2017-05-17
中国烟草学报(2016年1期)2016-11-16
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年4期)2016-05-17
湖南农业科学(2014年5期)2014-02-27
环境与可持续发展(2013年6期)2013-03-11
中国烟草学报(2012年6期)2012-04-09
植物营养与肥料学报(2011年3期)2011-10-24
