
干旱及复水对玉米叶片光合特性的影响
2012-05-07 11:07:26姚春霞张岁岐燕晓娟
水土保持研究 2012年3期
姚春霞,张岁岐,2,燕晓娟
(1.西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌712100;2.中国科学院 水利部 水土保持研究所,陕西 杨凌712100)
玉米(Zea mays L.)是极为重要的粮食、饲料、经济兼用作物。随着全球性的干旱日趋明显,水资源短缺也将会持续加剧[1]。而干旱作为限制作物产量提高的重要非生物胁迫因素,在各种环境胁迫因子中造成的损失最大[2]。干旱可以使作物从内到外发生一系列生理生态、生化及形态上的响应,其对植物地上部的影响主要是由于气孔关闭及二氧化碳供应的减少而引起叶片蒸腾作用的下降和光合作用的降低[3]。干旱胁迫还显著抑制了玉米根系的生长,减少了根系的吸收面积和活跃吸收面积[4]。白莉萍等[5]的研究表明,玉米各生育阶段受干旱胁迫将导致植株矮化,生长发育受阻,果穗性状恶化,以致于生物量和经济产量均大幅下降。但胁迫解除后作物生长会如何变化,胁迫时对作物造成的不利影响能否随胁迫的解除而消除尚不清楚。水分变动条件下,植物的生长存活和产量不仅仅取决于其对阶段干旱的忍耐能力,还决定于其对旱后复水的适应和对水分的利用能力[6]。苏佩等[7]发现玉米在多变低水条件下产量会减少,对叶片水分状况参数进行测定表明苗期干旱而拔节期复水增强了玉米对水分利用和其他生理过程的调节能力而表现出补偿效应。玉米苗期经干旱锻炼后复水,叶片和根部的水分状况得到改善,叶片可以较长时间内保持较强的渗透调节能力,这不仅有利于叶片的延伸生长,还使光合、气孔导度等生理代谢方面表现出有利植物生长的优势[8]。有研究表明,光合速率随水分胁迫加强而不断下降,是作物后期受旱减产的主要原因[9]。
水分胁迫导致PSII反应中心易产生光抑制现象,PSII的Fv/Fm降低是光合作用受抑制的显著特征。张林春等[8]发现干旱胁迫下,不同抗旱性玉米PSII的Fv/Fm、qP降低,复水后郑单958的光合、荧光迅速恢复,陕单902恢复缓慢。玉米叶片光合速率和叶水势在复水后也有不同程度的恢复[9]。以上研究多着重于苗期干旱后复水玉米补偿效应的变化,但对不同生育期干旱后复水,其补偿效应是否存在同样的变化有待于进一步研究。
干旱胁迫限制光合主要是通过气孔关闭还是通过代谢损耗,还存在比较大的争议[10]。以往对玉米的研究多侧重于苗期干旱及复水条件对叶片光合特性以及经济产量的影响[8]和干旱及复水对不同抗旱性玉米光合特性的影响 ,但对不同生育期干旱及复水对叶片光合特性的影响及其差异比较研究鲜有报道,尤其是对孕穗期光合特性的变化研究较少。因此,本实验选用玉米品种户单4号为试验材料,研究拔节期和孕穗期干旱及复水条件下玉米叶片光合参数和叶绿素荧光参数的变化特征,为玉米的补偿效应机制研究提供理论依据。
1 材料与方法
1.1 试验材料与方法
供试玉米品种采用户单4号,于2011年4—8月在中科院水土保持研究所防雨棚内完成。供试土壤为杨凌田间耕层土,毛管最大持水量为29%,土壤有机质、速效氮、速效磷、速效钾、全氮、全磷、全钾含量分别为8.25g/kg,30.27mg/kg,6.39mg/kg,137.03 mg/kg,0.53mg/kg,0.92mg/kg,13.52mg/kg。盆栽所用塑料桶直径为29.6cm,深度为28.5cm,每桶装风干土12kg。每桶施尿素3.50g,过磷酸钙3.28 g,硫酸钾1.37g,与土壤混匀后装桶。挑选大小一致的饱满种子在4月13日催芽并于4月15日进行播种,五叶一心定苗,每盆一株。
1.2 试验设计
试验设三个水分处理,即正常供水(CK)、中度水分胁迫(MS)和重度水分胁迫(SS),其土壤相对含水量分别为盆土田间持水量的75%±5%,55%±5%,35%±5%。于八叶一心时开始控水,采用称重法控制土壤含水量。对照组土壤含水量始终保持在75%±5%,待干旱处理水分降至胁迫标准后,测定各项指标,随后复水至对照水平,连续测定5d。分别在拔节期和孕穗期进行复水处理。共10个处理,每处理设5次重复。
1.3 测定指标及方法
1.3.1 叶片光合参数的测定 选择植株上部第五片完全展开叶,于上午9:30—10:00采用Li—6400便携式光合测定系统(Li—Cor,USA)在1 300μmol/(m2·s)光强下,测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、并计算气孔限制值(Ls)(Ls=1-Ci/Ca,Ca为大气CO2浓度)和单叶水分利用效率(WUE)(WUE=Pn/Tr,Tr为蒸腾速率)等气体交换参数。每处理重复5次。
1.3.2 叶片叶绿素荧光参数的测定 测定完叶片气体交换参数后,叶片暗适应30min,采用Imaging-PAM(WALZ,德国)测定光系统Ⅱ(PSⅡ)的最大光化学效率(Fv/Fm),光化学猝灭系数(qP),每个处理重复5次。
1.3.3 叶水势的测定 于早上6:00取各处理上数第5片完全展开叶,用压力室法测定叶中部水势。
1.4 数据处理与统计分析
用Excel 2003进行数据计算,用SigmaPlot 10.0作图,用SAS统计软件进行显著性分析。
2 结果与分析
2.1 干旱及复水条件下玉米叶水势的变化
叶水势是反映植物水分状况的一个生理指标。水分胁迫下,图1a和1b的正常供水处理(CK)的叶水势最高,中度胁迫(MS)的次之,重度胁迫(SS)的最小。复水后,拔节期MS和SS处理的叶水势迅速升高,并超过对照;孕穗期不同水分处理的叶水势都相应升高,MS和SS处理的变化幅度比较平缓,复水第4天SS与对照间无显著差异。
2.2 干旱及复水过程中玉米叶片光合参数的变化
2.2.1 拔节期干旱及复水过程中玉米叶片光合参数的变化 由图2a和2b可以看出,干旱胁迫降低了叶片净光合速率(Pn)及气孔导度(Gs)。中度胁迫下叶片的Pn与对照间无显著差异,重度胁迫下Pn极显著低于对照。复水后,Pn和Gs有不同程度的升高,在第3天降低。复水第1天中度胁迫下Pn与Gs超过对照和重度胁迫处理,随后逐渐降低;在第2天时重度胁迫的Pn与Gs达最高值;复水第4天,中度和重度处理恢复至对照,甚至超过对照。各处理下,光合速率的恢复均以中度水分胁迫处理较快,重度胁迫处理较慢。
图2b和表1中可看出,重度水分胁迫的胞间CO2浓度(Ci)最低,中度水分胁迫的Ci最高,重度水分胁迫的Ci最低,气孔限制值(Ls)最大,Pn降低的主要原因是气孔限制;中度胁迫的Ci最高,Ls最小,说明Pn降低的主要原因是非气孔限制。复水后,Ci升高,Gs增加,Pn也逐渐升高,在第5天恢复至对照。
叶片水分利用效率反映了叶片瞬间CO2固定与水分散失之间的关系,也是评价植物抗旱能力的综合指标之一。由图2d可以看出,重度胁迫叶片的WUE高于对照,中度胁迫的 WUE最低,各处理间差异不显著。复水第4天,中度胁迫和重度胁迫处理的WUE均高于对照,表现出补偿作用。
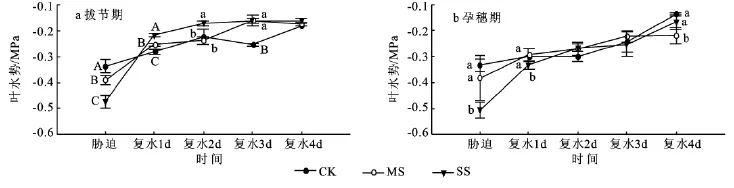
图1 拔节期和孕穗期干旱胁迫及复水条件下玉米叶水势的变化
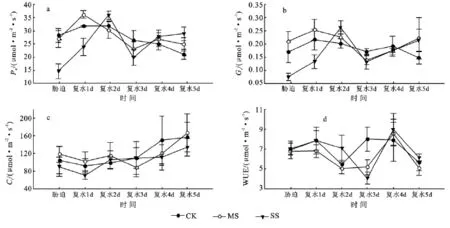
图2 拔节期干旱及复水过程中玉米叶片光合参数的变化
2.2.2 孕穗期干旱及复水过程中玉米叶片光合参数的变化 不同水分条件下,叶片的净光合速率(Pn)表现为:MS>CK>SS。与拔节期不同的是,复水第1天,CK和SS处理的Pn降低,其后SS处理的Pn迅速升高并在第3天达到最大值。Gs与Pn变化一致,MS与CK无显著差异,且显著高于SS,各处理在复水后第4天恢复到最大。图3c和表1中,重度胁迫的Ci、Gs较低,Ls最高,说明Pn的降低是由气孔限制造成的。图3d表明,干旱胁迫下MS处理叶片的WUE较高,SS处理叶片的WUE最低;WUE在复水第1天达到最大值,随后逐渐降低。
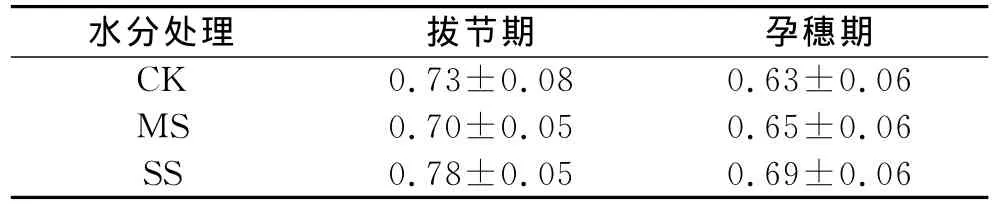
表1 玉米拔节期和孕穗期不同干旱胁迫下叶片气孔限制值(Ls)的比较
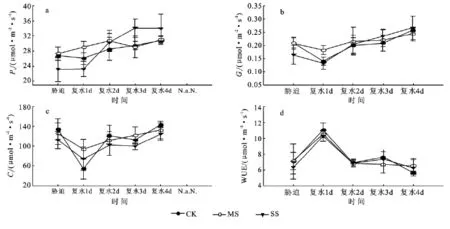
图3 孕穗期干旱及复水过程中玉米叶片光合参数的变化
2.3 干旱及复水条件下玉米叶片叶绿素荧光参数的变化
2.3.1 拔节期干旱及复水条件下玉米叶片叶绿素荧光参数的变化 Fv/Fm反映的是PSII原初光能转化效率及PSII潜在活性,该参数一般为0.75~0.85,但在逆境或受伤害时会明显降低。图4a中,重度胁迫叶片的Fv/Fm值显著低于正常供水(CK)和中度胁迫处理(MS),复水后逐渐升高。MS处理叶片的Fv/Fm值与CK间差异不显著,复水第3天各处理间无显著差异。
由图4b可看出,干旱胁迫下,光化学猝灭系数(qP)值降低,顺序为:CK>MS>SS。复水后三个处理逐渐升高,随后降低;复水第4天,SS处理的qP与CK和MS处理形成极显著差异。光化学猝灭系数(qP)值越大,说明PSII的电子传递活性越高。SS处理的qP值显著降低,表明重度胁迫条件下PSII反应中心的开放程度降低,光化学活性降低。复水第4天时干旱处理qP又继续升高,表现出滞后效应。
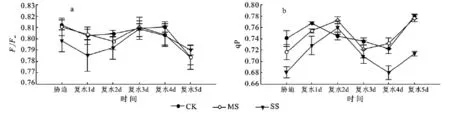
图4 拔节期干旱胁迫及复水条件下玉米叶片荧光特性的变化
2.3.2 孕穗期干旱及复水条件下玉米叶片叶绿素荧光参数的变化 图5所示是孕穗期玉米叶片Fv/Fm、qP的变化。与拔节期相比,重度胁迫(SS)和中度胁迫(MS)叶片的Fv/Fm值较低,复水后逐渐升高。复水第4天SS与CK、MS差异极显著。SS处理叶片的最大光化学效率(Fv/Fm)与其净光合速率(Pn)的变化趋势一致。光化学淬灭系数(qP)一定程度上反映了PSII反应中心的开放程度。干旱胁迫时,qP值降低(图5b),光合电子传递活性降低。复水后,逐渐升高。
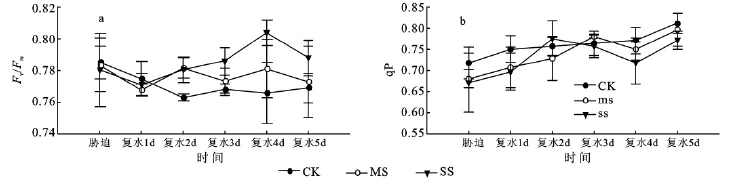
图5 孕穗期干旱胁迫及复水条件下玉米叶片荧光特性的变化
3 讨论
3.1 干旱及复水对玉米光合、荧光的影响
植物的光合作用可以将无机物转化为有机物,是地球上最重要的化学反应,也是绿色植物对各种内外因子的最敏感的生理过程之一。研究发现,干旱胁迫下植物光合速率降低的主要原因是受气孔的限制[11],而有些研究结果认为,水分亏缺条件下植物的光合速率降低的主要原因是受非气孔的限制。本实验中拔节期和孕穗期受水分胁迫后,中度水分胁迫(MS)和重度水分胁迫(SS)的净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci)降低,复水后,Ci和Pn升高,Gs增大,说明叶片在胁迫解除后有迅速恢复的能力。这与罗宏海[2]、张林春[8]等的研究一致。SS处理的Gs及Ci较低,气孔限制值(Ls)最高,说明Pn降低是气孔限制造成。这与其他研究结论有所不同[12],可能是由于试验条件和品种不同。
杨晓青等[13]研究发现,水分胁迫下小麦Fv/Fm、qP显著降低,NPQ升高。本研究中,拔节期和孕穗期干旱胁迫下最大光化学效率(Fv/Fm)值降低,说明叶片PSII反应中心的光化学活性降低,同时导致过剩的激发能的增大。拔节期重度胁迫的Ls与对照相比增加了6.85%,Fv/Fm值降低了1.72%,孕穗期重度胁迫的Ls增加了9.52%,Fv/Fm值降低了0.61%,表明两个时期叶片Pn降低主要都是气孔限制造成的。光化学淬灭系数(qP)表示PSII天线色素吸收的光能用于光化学反应的份额,一定程度上反映了PSII反应中心的开放程度[14]。干旱处理下的光化学淬灭系数(qP)低于对照,表明干旱胁迫导致PSII反应中心的开放度降低,光合电子传递活性下降,导致PSII反应中心的光能过剩,对光合机构起到了一定的保护作用。复水后,Fv/Fm、qP这两个参数值基本恢复到对照水平。表明在干旱胁迫得到解除后,受到伤害的PSII反应中心可以在一定程度上得以恢复。
单叶水分利用效率(WUE)表征植物对自身蒸腾耗水量的利用能力,是提高大田水分利用效率的生理基础[15]。研究表明,前期干旱锻炼能提高玉米的水分利用效率,有利于农业增产和节水[16]。本研究中,重度胁迫叶的WUE在胁迫及复水过程中均高于对照,表明干旱胁迫提高了水分利用效率;中度胁迫叶的WUE在复水第4天时超过对照,与徐国山[12]等的研究结果一致。孕穗期三个处理叶片的WUE在复水第1天达到最大值,随后趋于降低。可能是因为复水初期,补充的水分主要用于恢复自身生长,消耗掉的水分相对减少。
3.2 外界环境因素对复水后光合恢复的影响
复水后植物水分状况及生理活性的恢复与干旱胁迫程度及复水后的外界条件有关。范雯雯等[17]在小麦实验中发现,高肥力土壤有利于复水后光合速率的恢复;Boyer[18]比较了玉米、向日葵叶片光合速率和水势的关系,发现低光强下,即使干旱时叶水势低于-1.7MPa,光合也可在复水后15h内恢复;但在高光强下,叶水势即使低于-1.2MPa,复水三天后光合也不可能完全恢复。本实验中拔节期光合速率在复水后逐渐增加,在第3天骤降,主要原因应为阴雨天气影响光照弱,温度低,空气相对湿度增大,气孔导度明显减小,进入叶片内二氧化碳浓度降低,以致光合速率显著减小。
4 结论
玉米受旱后生理代谢发生一系列适应性变化,一旦解除干旱并恢复供水,同样发生一系列生理代谢等方面恢复和修复作用。复水后,叶片水分状况得到改善,气孔完全张开,气孔导度增大,光合速率也增大。拔节期Pn、Gs在复水后第2天达到最大值,由于外界环境影响,Pn下降;而孕穗期Pn、Gs复水后第4天达最大值。干旱胁迫伤害光合机构PSII,PSII主动调节电子传递速率和光化学效率,减轻过剩光能对其系统的损伤。孕穗期叶片的Fv/Fm值低于拔节期,且两时期的Fv/Fm值在复水第1天不能立即恢复,这是由于胁迫程度加剧复水后叶片的潜在光合能力未被激发出来,在复水第2天逐渐恢复。两时期干旱处理在复水条件下,qP变化趋势一致,基本恢复至对照。这是由于Gs增加了叶内部二氧化碳的量,提高了叶片内碳同化的效率,从而使光合电子传递活性增强。本研究证明了拔节期和孕穗期光合特性变化存在差异,但全生育期干旱及复水过程中玉米光合特性和叶水势具体变化过程以及各参数之间的内在联系还有待于更深入的研究。
[1] 山仑.植物水分利用效率和半干旱地区农业用水[J].植物生理学通讯,1994,30(1):61-66.
[2] 罗宏海,张亚黎,张旺锋,等.新疆滴灌棉花花铃期干旱复水对叶片光合特性及产量的影响[J].作物学报,2008,34(1):171-174.
[3] 李文娆,张岁岐,山仑.水分胁迫对紫花苜蓿根系吸水与光合特性的影响[J].草地学报,2007,15(3):206-211.
[4] 韩希英,宋凤斌.干旱胁迫对玉米根系生长及根际养分的影响[J].水土保持学报,2006,20(3):170-172.
[5] Bai L P,Sui F G,Sun C H,et al.Effects of soil water stress on morphological development and yield of maize[J].Acta Ecologca Sinica,2004,24(7):1556-1560.
[6] Reynolds J F,Kemp P R,Oqle K,et al.Modifying the‘pulse-reserve’paradigm for deserts of North America:precipitation pulse,soil water,and plant responses[J].Oecologia,2004,141(2):194-210.
[7] 苏佩,山仑.多变低水环境下高粱、玉米籽粒产量及水分利用效率变化的生理基础研究[M]∥植物生理学与跨世纪农业研究.北京:科学出版社,1999:199-202.
[8] 张林春,郝扬,张仁和,等.干旱及复水对不同抗旱性玉米光合特性的影响[J].西北农业学报,2010,19(5):76-80.
[9] 赵天宏,沈秀瑛,杨德光,等.水分胁迫及复水对玉米叶片叶绿素含量和光合作用的影响[J].杂粮作物,2003,23(1):33-35.
[10] 王磊,胡楠,张彤,等.干旱和复水对大豆叶片光合及叶绿素荧光的影响[J].生态学报,2007,27(9):3631-3635.
[11] Lakso A N.Seasonal changes in stomata responses to leaf water potential in apple[J].Am.Soc.Hort.Sci.,1979,10(4):58-60.
[12] 徐国山,邓丽娟,李雪丽,等.土壤干旱胁迫及复水对紫叶矮樱光合特性的影响[J].河北林果研究,2010,25(2):116-120.
[13] 杨晓青,张岁岐,梁宗锁,等.水分胁迫对不同抗旱类型冬小麦幼苗叶绿素荧光参数的影响[J].西北植物学报,2004,24(5):812-816.
[14] 宋莉英,孙兰兰,舒展,等.干旱和复水对入侵植物三裂叶蟛蜞菊叶片叶绿素荧光特性的影响[J].生态学报,2009,29(7):3713-3720.
[15] Du Z C,Deng X P,Zhao C X.Anti-oxidation capacities of different wheat genotypes under water stress[J].Acta Botanica Boreal-Occident Sinica,2005,2(8):1672-1676.
[16] 黄占斌,山仑.水分利用效率及其生理生态机理研究进展[J].生态农业研究,1998,16(4):19-23.
[17] 范雯雯,王志强,林同保,等.不同土壤肥力下旱后复水对冬小麦光合特性及水分利用效率的影响[J].麦类作物学报,2010,30(2):362-365.
[18] Boyer J S.Leaf enlargement and metabolic rates in corn,soybean,and sunflower at various leaf water potentials[J].Plant Physiology,1970,46(2):233-235.
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
今日农业(2021年6期)2021-11-27 08:05:59
食品安全导刊(2020年33期)2020-12-26 07:38:20
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
山东农业科学(2017年4期)2017-05-18 17:07:16
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
动物营养学报(2015年10期)2015-12-01 06:45:19
江苏农业科学(2014年10期)2014-11-22 13:23:56
江苏农业科学(2014年9期)2014-11-15 02:14:26
