
苏北海滨湿地互花米草地上生物量动态*
2012-01-06 08:59:10徐伟伟王国祥刘金娥陈正勇吴晓东
海洋科学进展 2012年1期
徐伟伟,王国祥,刘金娥*,陈正勇,宋 康,吴晓东
(1.南京师范大学 地理科学学院,江苏 南京210046;2.江苏省环境演变与生态建设重点实验室,江苏 南京210046)
苏北海滨湿地互花米草地上生物量动态*
徐伟伟1,2,王国祥1,2,刘金娥1,2*,陈正勇1,2,宋 康1,2,吴晓东1,2
(1.南京师范大学 地理科学学院,江苏 南京210046;2.江苏省环境演变与生态建设重点实验室,江苏 南京210046)
在江苏盐城新洋港滩涂由海向陆建立样地:零星米草斑块(SAP)、稳定米草滩下边缘(SAFI)、2003年米草定居处(SAF03)、1989年米草定居处(SAF89),对互花米草(Spartina alterniflora)的生物量动态变化进行了研究。结果表明:1)除SAF03样地外,其他样地互花米草叶生物量随时间的推移都有不同程度的减少,SAF03叶生物量始终都显著高于其他样地(P<0.05);各样地茎和繁殖器官生物量均有所增加,繁殖前期SAFI处茎生物量增长最快,达18.97g·m2·d-1;离海越近繁殖器官生物量增加越大;11月份SAF89样地各器官生物量顺序为:叶>茎>繁殖器官,其他样地:茎>叶>繁殖器官。2)各次调查中,随着时间的推移各样地叶生物量分配减少,而茎生物量与繁殖器官生物量分配增多;由陆向海各样地叶生物量分配减少,而茎生物量与繁殖器官生物量分配增加。3)SAFI处互花米草地下/地上生物量的比值为2.39,极显著高于其他样地(P<0.01)。
外来种;生物量分配;生长特性;海滨湿地
湿地是介于水体和陆地之间的生态交错区,兼有陆生和水生生态的特点,是地球上生产力最高的生态系统之一[1]。而湿地植被是湿地生态系统的生产者,在生长过程中吸收CO2,形成光合产物并作为生物量固定储存起来,其含量多少显著地影响湿地生态系统的生产力,一直受到众多学者的高度关注。此外,湿地植被在稳定全球气候、减缓温室效应方面发挥了重要作用[2]。植物在不同环境与生长压力下具有不同的生长特性与物质分配规律[3],这是植物与环境条件共同作用的结果,反映了植物个体对环境条件的适应能力和生长发育规律,也反映了环境条件对植物的影响和饰变程度[4]。
互花米草(Spartina alterniflora)是一种原产美洲大西洋沿岸的多年生草本植物,主要生长于平均海平面至平均高潮位之间的广阔滩面以及河口湾,从加拿大的纽芬兰到南美的法属圭亚那潮间带均有分布[5]。互花米草能够保滩护岸,加快淤积,被许多国家引种,包括北美西海岸、欧洲、新西兰和中国沿海等,但由于在许多引种地快速蔓延[6]引起广泛关注,并且米草属植物已成为研究生物入侵生态学和遗传学特征的模式植物[7]。江苏沿海的互花米草在1990年前后形成较大面积群落,之后面积扩展迅速,成为优势盐沼植被,目前江苏沿海互花米草盐沼群落仍处于盐沼形成的初期[8]。
目前关于互花米草在生理[9-11]、生态功能[12-13]、扩散能力及引种繁殖[14-15]等方面的研究在国内外已有大量的文献报道;对于互花米草入侵河口湾和海湾潮间带生态系统的生态影响和后果,一些研究者已经进行过总结,并提出了管理对策[5,16-17];对克隆植物互花米草表型可塑性也有文献涉及[18]。而研究不同潮滩带种群生物量的动态变化,对于了解种群在不同生境的生物生态学特性和生产力状况及掌握植物种群对环境条件的适应能力有重要的意义。本研究对互花米草从快速生长前期到种子成熟期的调查与研究,就自然条件下不同潮滩带互花米草种群地上生物量动态变化作定量分析与研究,揭示成熟期互花米草生物量分配的规律,为互花米草在滩涂的扩张预防及恢复本地种提供科学依据,为防止互花米草在我国的进一步扩张以及对不同区域进行有效管理和控制提供参考,并为今后互花米草对碳储备的系统研究提供基础资料。
1 研究地区与研究方法
1.1 研究区概况
研究区位于江苏省盐城市新洋港滩涂(32°34′~34°28′N,119°48′~120°56′E),该区是温带和亚热带的过度地带,受海洋性与大陆性气候的共同影响,季风盛行,四季分明,年平均气温13.8℃,日均光辐射总量116~121 kcal·cm-2,年均降雨量1 023.8mm,降水集中在夏季。水源有陆地水和海洋水,海水淹没潮间带间隔7~12 h,高潮位1.27~4.61m。研究区新洋港是受到人类干扰较少的潮滩湿地,保留着较完整的潮滩植被演替序列,由海向陆依次为:光滩,互花米草滩,盐地碱蓬(Suaeda salsa)滩及由藨草(Scirpus trqueter)、獐毛(Aeluropus littoralis)、白茅(Imperata cylindrical)、芦苇(Phragmites australis)组成的禾草滩。
1.2 研究方法
1.2.1 样地选择与野外取样
在江苏盐城新洋港滩涂的互花米草盐沼区域,按照扩散前沿区和不同发育年限的互花米草种群稳定区的序列建立4个野外观测带,分别为零星米草斑块(S.alterniflorapatch)、稳定米草滩下边缘(S.alternifloraflat infra-edge)、2003年米草定居处(S.alternifloraflat 2003)、1989年米草定居处(S.alternifloraflat 1889),文中各样地分别用SAP,SAFI,SAF03和SAF89代表(表1)。于2009-04-24GPS定位各样地进行动态监测,并测量出距海堤距离及高程。2009-08-16,09-23,11-11统计调查互花米草的生长、繁殖及生物量指标,每个样地设置3个重复。各次调查取样在设置的每个样地随机选择3处25×25cm2小样方,齐地面剪下,在2009-11-11采集了地下生物量,做好标记带回实验室测定。3次采样分别代表了开花前快速生长期、开花期和种子成熟期。

图1 新洋港滨海湿地采样点分布Fig.1 Locations of sampling in the Xinyanggang coastal wetland
1.2.2 生物量及理化指标的测定
测定指标包括根、茎、叶、小穗、种子以及全株生物量。将每株的根、茎、叶、小穗、种子分装,杀青后用80℃恒温烘干,称量。土壤含水量采用烘干法测定,全盐采用电导法测定[19]。
1.2.3 数据处理
根据野外调查数据和室内的分析结果,采用SPSS 11.5软件,选择单因素方差分析(One-Way ANOVA)进行差异显著性检验。
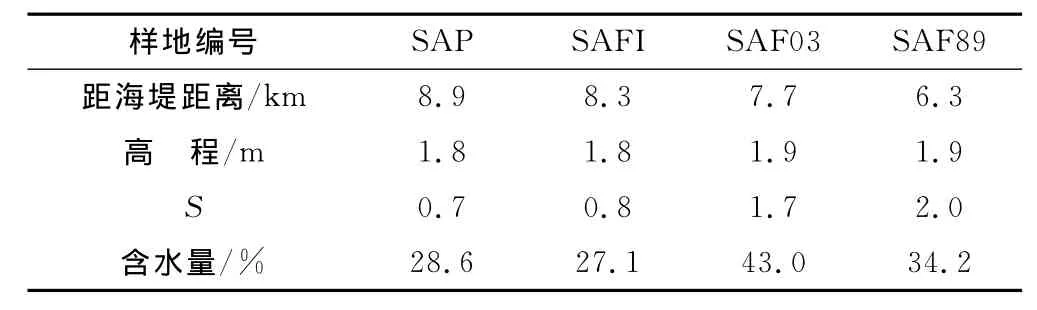
表1 样地描述Table 1 Descriptions of the sample plots
2 结果与分析
2.1 不同潮滩带互花米草的地上生物量组成
相对8月份来说,除11月份SAF03样地外,互花米草叶生物量随时间的推移都有不同程度的减少(图2),除8月份SAF03与SAF89无显著性差异外(P>0.05),SAF03叶生物量一直都显著高于其他样地(P<0.05);而茎生物量随时间的推移都有不同程度的增加,到11月份,以SAF03样地增加的量最多,总量达1 042.88g/m2,而SAF89处茎生物量一直没有变化。繁殖器官生物量在9月份时无显著性差异,到11月份互花米草种子成熟时,SAP样地处单位面积繁殖器官生物量最大,而SAP89处最小,二者间存在极显著性差异(P<0.01),SAP样地和其他样地间有显著性差异(P<0.05)。8月份生物量大小顺序都是:叶>茎,除9月份SAF03和SAF89样地及11月份的SAF89顺序为:叶>茎>繁殖器官,其他:茎>叶>繁殖器官。互花米草地上生物量各次采样中都是以SAF03样地处最高,表明此样地其叶面积总量大、茎粗壮,这些特点有利于植株同化足够的养分及种群进一步扩展,不同潮滩带地上生物量的变化趋势不同也表征着他们生长特性及发育成熟阶段的不同。
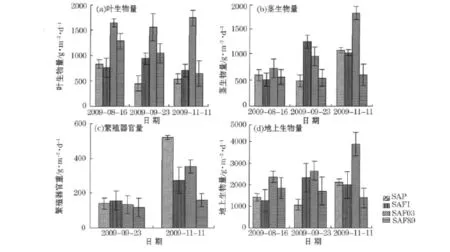
图2 不同潮滩带互花米草的地上生物量变化Fig.2 Variations of the above-ground biomass of S.alterniflorain different intertidal flats
2.2 不同潮滩带互花米草地上各器官生物量生长动态
互花米草各器官生物量随时间变化趋势与单位面积地上生物量变化趋势基本一致。2009-08-16—09-23,叶生物量除SAFI样地增加4.78g·m-2·d-1,其他样地都是减少的,减少量最大的是SAP样地,为10.07g·m-2·d-1,茎生物量中SAP与SAF89分别减少2.95和0.45g·m-2·d-1,而SAFI与SAF03处分别增加18.97和5.93g·m-2·d-1;2009-09-23—11-11,叶生物量SAP与SAF03处分别增加1.96和3.90g·m-2·d-1,SAFI与SAF89分别减少4.86和8.39g·m-2·d-1,茎生物量除SAFI减少4.39 g·m-2·d-1外,其他均呈正增长,最大为SAF03处的17.03g·m-2·d-1;繁殖器官生物量都呈正增长的趋势,从大到小为SAP,SAF03,SAFI和SAF89,分别为7.92,4.57,2.39和0.88g·m-2·d-1。
2.3 不同潮滩带互花米草各器官地上生物量的比例动态
各器官生物量在个体生物量中所占的比例代表着同化产物向不同器官的分配比例和终生生长过程中各器官协调关系[4]。各个样地的叶生物量分配随时间的推移都有不同程度的减少,不同时期各潮滩带互花米草不同器官生物量分配大小顺序几乎没有变化(图3)。各次调查中叶生物量分配呈现出潮滩带越低生物量分配越多的趋势,茎生物量和繁殖器官生物量相反;到11月份,叶生物量以SAP样地处减少量最多,达33%,而茎生物量分配都有不同程度的增加,在SAF03处增加最大,增加了15%。从8月种群进入繁殖期,植株开始将存储的能量更多地转入到繁殖器官中,以利于种群完成繁殖过程[20]。繁殖器官生物量分配都有明显增加,以SAP处生物量分配最大并且增加的最快,增加了11%,与其他样地有显著性差异(P<0.05),但总的来说,繁殖器官生物量占地上生物量的比例很小。

图3 不同潮滩带互花米草不同器官地上生物量分配动态Fig.3 Dynamics in the above-ground biomass proportion of different organs of S.alterniflorain the different intertidal flats
2.4 不同潮滩带互花米草地下部分与地上部分生物量构成
植物各部分是一个统一的整体,地上部分对地下部分的生长发育有重要的影响,是地下部分生物量的能量来源,又依靠地下部分吸收生长所需要的水分和营养物质。地下/地上生物量的比值是描述地上部分生物量与地下部分生物量之间关系的一个重要参数,它反映了分配给地下部分光合产物的比例,其变化反映了光合作用物质在植物体内的分布和转移[21]。图4是2009-11采集的数据,可知SAFI处互花米草地下部分生物量是地上的2.39倍,SAF89处地下生物量略大于地上生物量,而SAP和SAF03地下生物量分别是地上生物量的0.64和0.40倍。地下生物量SAFI处与其他各样地都有极显著差异(P<0.01)。
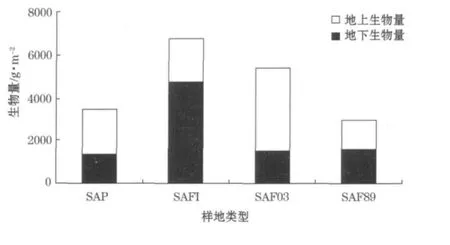
图4 不同潮滩带互花米草地下部分生物量与地上部分生物量Fig.4 The above-and under-ground biomasses of S.alterniflorain the different intertidal flats
3 讨 论
3.1 生物量变化
江苏盐城新洋港滩涂内的互花米草,从8月开始进入开花繁殖期,此后,除SAF03处互花米草单位面积叶生物量略有增加外,其他样地均出现负增长,而茎生物量是有不同程度的增长,说明8月后不同潮滩带互花米草叶组织逐渐枯萎,光合产物减少,但并没有完全抑制茎生物量的增加,茎生物量的增长在SAF03样地处表现的最为明显,原因可能是该处生境土壤含水量较高、盐度适中,潮侵频率所带来的淹水胁迫及受海浪直接冲刷等因子影响较小,从而有利于植物生长。SAF03处叶生物量一直最高,其地上生物量也一直最高,营养枝叶生殖分配的高比例有利于增大种群的光合面积,而增加光合产物的积累,既保证有性生殖的生长,也为营养生长提供了保障。经实验数据分析得知,互花米草个体茎的生长发育和地上生物量动态趋势基本一致,而部分样地不同调查时间段叶生物量和地上生物量动态趋势相反,主要因为适宜的潮汐、水分和盐分条件有利于叶生物量的积累,但叶生物量的变化与叶的枯落有很大关系,这方面的实验设计有待进一步完善。叶生物量在SAP和SAF89样地下降最为多,说明该处的互花米草较先枯萎,结合野外对互花米草开花及结实的观测,我们发现SAP和SAF89样地的互花米草较先成熟,这跟该生境中不利植物生长的条件有关[22],SAP因近海受潮水淹没时间相对较长,而SAF89处是互花米草的最初引种地,沉积导致高程增加,降低了局部地带的潮浸频率,出现局部高地的返盐现象,使沉积物含盐量增加,且土壤含水量较低(表1),这两处生境互花米草为生长繁殖相对提前进入成熟期。
3.2 生物量分配
在不同的潮汐、水分和盐分条件下,植物总是对生长、繁殖和维持等功能间进行资源分配的权衡(trade-off),使其各功能分配达到最佳状态[23-24],植物的能量分配格局反映了植物在生长发育过程中对环境的响应规律和能量分配对策[25]。各次调查中,基本呈现离海越近,即在越低的潮滩带,叶生物量分配越少,而茎生物量分配越多,这说明了离海越近互花米草植株需要更大生物量的茎组织来适应低潮滩长时间、高深度及涨落潮流动海水的淹水环境。
有性繁殖在对不同环境的适应能力方面存在着优越性。在相同环境条件下,土壤条件,水热条件基本相同时,植物同化的资源是一定的,此时生殖枝的生殖构件穗和营养构件叶、茎之间的生物量分配具有高度拮抗作用,分配到营养构件的生物量多,必然导致分配到生殖构件的就少,有研究发现入侵种能通过调节植株大小和种子产量等而使其在不同演替阶段的群落中立足,占据不同发育阶段的滩涂[26-27]。植物生物产量是通过2个生理过程形成的:1)根的吸收作用;2)叶片的光合作用。通常叶片光合产物积累量远远超过根系的矿物质积累量,而且生物产量是籽粒产量的基础,种子生殖分配比例的多少直接影响下一代的生长发育和种群竞争力[28-29],没有高额的生物产量便不可能有高额的籽粒产量[30]。SAP处繁殖器官生物量分配在9月和11月都是最大并增加的最快,且繁殖器官生物量也是最大;繁殖器官的生物量是随着地上生物量的增加而增加的,繁殖器官生物量的增加是以减少其他器官生物量分配为代价的,但SAP地上生物量偏小,而离海最近,较大繁殖器官生物量分配可能意味着近海互花米草更有通过种子扩散的可能性及对有性繁殖的趋向。互花米草地下部分生物量与地上部分生物量比值的变化在很大程度上是由地上部分生物量的变化引起的,互花米草把吸收的大部分营养物质投资到地上部分茎、叶的生长上,以利于植株对光资源的摄取和加强种群的竞争力[21],SAFI样地处较大的地下生物量可能跟潮水、淹水时间有关,该处风浪强度在一定程度上被SAP有所削弱,同时该样地互花米草定居时间较SAP处长,土壤营养丰富,且比其他样地有更好的水分条件,促进了地下生物量的增加。
3.2 防治建议
结合野外试验调查及近年国内外相关的防治结果,认为互花米草的繁殖与扩散主要是以克隆繁殖体萌蘖和根状茎快速生长为主[15]。这种营养繁殖特征可有效保障互花米草种群的维持和更新,种子繁殖受潮水冲刷及土壤水分影响较明显,但其对占据新生境的贡献较大,而在近海的SAFI处互花米草地下部分生物量是地上的2.39倍,因此,在互花米草的防治过程中,应对不同潮滩带实施不同的防治方式,就我们研究地新洋港滩涂而言,从生态和经济的综合角度出发,较高高程互花米草滩应以抑制其营养繁殖为主要方式来减少互花米草对当地种的入侵负面影响。有研究表明从米草滩的前缘开始沿高程梯度逐步向上控制可以节约44%的时间和经费[31]。如欲控制互花米草往海方向的扩散,以抑制其种子繁殖和营养繁殖结合的方式,采取拔除、挖掘、在目标米草丛周围挖环状沟等措施,提高滩涂生物多样性,发挥沿海滩涂的最大积极效应。
4 结 论
1)调查期间各样地茎和繁殖器官生物量均有所增加,繁殖前期SAFI处茎生物量增长最快,达18.97 g·m2·d-1。SAF03叶生物量始终都显著高于其他样地,其他样地互花米草叶生物量随时间的推移都有不同程度的减少;离海越近繁殖器官生物量增加越大。11月SAF89样地各器官生物量顺序为:叶>茎>繁殖器官,其他样地:茎>叶>繁殖器官。
2)各次调查中,随着时间的推移各样地叶生物量分配减少,而茎生物量与繁殖器官生物量分配增多,且表现出由陆向海各样地叶生物量分配减少,而茎生物量与繁殖器官生物量分配增加的趋势,繁殖器官生物量占地上生物量的比例不大。
3)SAFI处互花米草地下/地上生物量的比值为2.39,极显著高于其他样地。
(References):
[1]LU J J,HE W S,TONG C F,et al.Wetland Ecology[M].Beijing:Higher Education Press,2006.陆健健,何文珊,童春富,等.湿地生态学[M].北京:高等教育出版社,2006.
[2]BAZHIN N M.Influence of plants on the methane emission from sediments[J].Chemosphere,2004,54:209-215.
[3]XU Z Z,ZHOU G S.Effects of soil moisture on growth characteristics ofLeymus chinensisseedlings under different temperature conditions[J].Chinese Journal of Ecology,2005,24(3):256-260.许振柱,周广胜.不同温度条件下土壤水分对羊草幼苗生长的影响[J].生态学杂志,2005,24(3):256-260.
[4]ZHANG W H,LI H,LI J X,et al.Individual and modular biomass dynamics ofKingdonia uninflorapopulation in Qinling Mountain[J].Chinese Journal of Applied Ecology,2003,14(4):530-534.张文辉,李红,李景侠,等.秦岭独叶草种群个体和构件生物量动态研究[J].应用生态学报,2003,14(4):530-534.
[5]DAEHLER C C,STRONG D R.Status,prediction and prevention of introduced cordgrassSpartinaspp invasions in Pacific estuaries,USA[J].Biological Conservation,1996,78:51-58.
[6]AYRES D R,STRONG D R.TheSpartinainvasion of San Francisco Bay[J].Aquatic Nuisance Species Digest,2002,4:38-40.
[7]BLUM M J,SLOOP C M,AYRES D R,et al.Characterization of microsatellite loci inSpartinaspecies(Poaceae)[J].Molecular Ecology Notes,2004,4:39-42.
[8]LIU J E,ZHOU H X,QIN P,et al.Effects ofSpartina alterniflorasalt marshes on organic carbon acquisition in intertidal zones of Jiangsu Province,China[J].Ecological Engineering,2007,7:240-249.
[9]QIN P,MA L K,XIE M,et al.A study on dynamics of Fe,Cu,Mn,Zn in primary production ofspartina alterniflora[J].Acta Ecologica Sinica,1993,13(1):67-74.钦佩,马连琨,谢 民,等.Fe、Cu、Mn、Zn在互花米草(Spartina alterniflora)初级生产中的动态研究[J].生态学报,1993,13(1):67-74.
[10]YANG X M,QIN P,XIE M,et al.A correlation research between physiological compositions ofSpartina alternifloraand artificial seawater concentrations[J].Chinese Journal of Ecology,1997,16(3):7-10.杨晓梅,钦佩,谢民,等.人工海水环境中互花米草总黄酮等生理成分与盐浓度的相关性研究[J].生态学杂志,1997,16(3):7-10.
[11]XIAO Q,ZHENG H L,YE W J,et al.Effects of waterlogging on growth and physiology ofSpartina alternifloar[J].Chinese Journal of Ecology,2005,24(9):1025-1028.肖强,郑海雷,叶文景,等.水淹对互花米草生长及生理的影响[J].生态学杂志,2005,24(9):1025-1028.
[12]SUN S C,ZHU X B,LV C Q.Function assessment and ecological controls on alien species cordgrass[J].Chinese Journal of Ecology,2004,23(3):93-98.孙书存,朱旭斌,吕超群.外来种米草的生态功能评价与控制[J].生态学杂志,2004,23(3):93-98.
[13]XIE Z F,HE W S,LIU W L,et al.Influence ofSpartina alterniflorasaltmarsh at its different development stages on macrobenthos[J].Chinese Journal of Ecology,2008,27(1):63-67.谢志发,何文珊,刘文亮,等.不同发育时间的互花米草盐沼对大型底栖动物群落的影响[J].生态学杂志,2008,27(1):63-67.
[14]ZHANG D,YANG M M,LI J X,et al.Vegetative Dispersal Ability ofSpartina alterniflorain Eastern End of Chongming Island[J].Journal of East China Normal University:Natural Science,2006,5(2):130-135.张东,杨明明,李俊祥,等.崇明东滩互花米草的无性扩散能力[J].华东师范大学学报:自然科学版,2006,5(2):130-135.
[15]DENG Z F,AN S Q,ZHI Y B,et al.Preliminary studies on invasive model and outbreak mechanism of exotic species,Spartina alternifloraLoisel[J].Acta Ecologica Sinica,2006,26(8):2678-2686.邓自发,安树青,智颖飙,等.外来种互花米草入侵模式与爆发机制[J].生态学报,2006,26(8):2678-2686.
[16]CHEN Z Y,LI B,CHEN J K.Ecological consequences and management ofSpartinaspp.invasions in coastal ecosystems[J].Biodiversity Science,2004,12(2):280-289.陈中义,李博,陈家宽.米草属植物入侵的生态后果及管理对策[J].生物多样性,2004,12(2):280-289.
[17]ZHANG X L.Analysis on the advantages and disadvantages of introducingSpartinato China coastal zones[J].Chinese Journal of Ecology,2007,26(11):1878-1883.张秀玲.米草属引入中国海岸带的利弊分析[J].生态学杂志,2007,26(11):1878-1883.
[18]HE J,ZHAO C J,QING H,et al.Effect of soil-water condition on morphological plasticity of clonal plantSpartina alterniflora[J].Acta Ecologica Sinica,2009,29(7):3518-3524.何军,赵聪蛟,清华,等.土壤水分条件对克隆植物互花米草表型可塑性的影响[J].生态学报.2009,29(7):3518-3524.
[19]LIU G S.Soil physical and chemical analysis and description of soil profiles[M].Beijing:Standard Press of China,1996.刘光菘.土壤理化分析与剖面描述[M].北京:中国标准出版社,1996.
[20]QIN Y Y,JIANG X X,LI F,et al.Biomass dynamics ofSpartina alternifloraat its sexual propagation stage in Shankou mangrove area of Guangxi China[J].Chinese Journal of Ecology,2008,27(12):2083-2086.覃盈盈,蒋潇潇,李峰,等.山口红树林区互花米草有性繁殖期的生物量动态[J].生态学杂志,2008,27(12):2083-2086.
[21]MOKANY K,RAISON R J,PROKUSHKIN A S.Critical analysis of root:shoot ratios in terrestrial biomes[J].Global Change Biology,2006,12:84-96.
[22]MAO Z G,WANG G X,LIU J E,et al.Characteristics of grain-size distributions in different vegetation zones of coastal wetland in north Jiangsu[J].Advances in Marine Science,2008,31(4):454-463.毛志刚,王国祥,刘金娥,等.苏北滨海湿地不同植被带沉积物粒度特征分析[J].海洋科学进展,2008,31(4):454-463.
[23]DONG M.Clonal growth in plants in relation to resource heterogeneity:foraging behavior[J].Acta Bontanica Sinica,1996,38(10),828-835.董鸣.克隆生长的特性——觅食性[J].生态学报,1996,38(10):828-835.
[24]WANG Y H,WANG K,XING F.Advances of studies on the morphological plasticity,integration and foraging behavior of stoloniferous herbaceous plants[J].Chinese Journal of Ecology,2005,24(1):70-74.王艳红,王珂,邢福.匍匐茎草本植物形态可塑性、整合作用与觅食行为研究进展[J].生态学杂志,2005,24(1):70-74.
[25]ZHAO L,ZHI Y B,LI H L,et al.Effects of initial clone number on morphological plasticity and biomass allocation of the invasivespartina anglica[J].Journal of Plant Ecology,2007,31(4):607-612.赵磊,智颖飙,李红丽,等.初始克隆分株数对大米草表型可塑性及生物量分配的影响[J].植物生态学报,2007,31(4):607-612.
[26]MARKS T C,TRUSCOTT A J.Variation in seed production and germination ofSpartinaanglica with in a zoned salt marsh[J].The Journal of Ecology,1985,73:695-705.
[27]THOMPSON J D,MCNEILLY T,GRAY A J.Population variation inSpartinaanglica C.E.Hubbard,1.Evidence from a common garden experiment[J].New Phytologist,1991,117:115-128.
[28]HARPER J L.The Population Biology of Plants[M].London:Academic Press,1977.
[29]DONG K H,MI J.Quantitative characters of vegetative multiplication ofBothriochloa ischaemumPopulations[J].Acta Agrestia Sinica,2006,14(3):210-213.董宽虎,米佳.白羊草种群繁殖的数量特征[J].草地学报,2006,14(3):210-213.
[30]GAO Q,GUO Z J,WANG P C,et al.Study on biomass reproductive allocation in four kinds of rhizomatous grass populations in Daqingshan Mountain[J].Chinese Agricultural Science Bulletin,2008,24(5):22-25.高强,国振杰,王普昶,等.大青山4种根茎禾草种群生物量生殖分配研究[J].中国农学通报,2008,24(5):22-25.
[31]GREVSTAD F S.Simulating control strategies for a spatially structured weed invasion:Spartina alterniflora(Loisel)in Pacific Coast estuaries[J].Biological Invasions,2005,7:665-677.
Above-ground Biomass Dynamics ofSpartina Alterniflorain the Coastal Wetland of North Jiangsu
XU Wei-wei1,2,WANG Guo-xiang1,2,LIU Jin-e1,2,CHEN Zheng-yong1,2,SONG Kang1,2,WU Xiao-dong1,2
(1.School of Geography Science,Nanjing Normal University,Nanjing 210046,China;2.Jiangsu Key Laboratory of Environmental Change and Ecological Construction,Nanjing 210046,China)
Sample plots includingSpartina alterniflorapatch (SAP),S.alternifloraflat infra-edge(SAFI),S.alternifloraflat 2003(SAF03)andS.alternifloraflat 1989(SAF89)were established landward on the intertidal flat of Xinyanggang in Yancheng of North Jiangsu,in which the above-ground biomass dynamics ofS.alterniflorawas studied.The results show that:1)The leaf biomass ofS.alterniflorahas,in different degrees,reduced with time except for SAF03,where the leaf biomass is always significantly higher(P<0.05)than other plots.The biomasses of the stem and the reproductive organs increase to some extent at each of the plots,of which the stem biomass increases fast before the propagation stage,reaching to 18.97g·m2·d-1.The biomass of the reproductive organs increases in a large amount in the near-sea area.In November,the order of the biomass of different organs is,from higher to lower,leaf>stem>reproductive organs in the SAF89plot and stem>leaf>reproductive organs in other plots;2)Both with the course of time and from the land toward the sea,the leaf biomass decreases in its proportion,while the biomasses of both the stem and the reproductive organs increase in proportion;3)The ratio of the Under-/above-ground biomasses ofS.alterniflorais 2.39in the SAFI plot,being extremely higher than those(P<0.01)in other plots.
exotic species;biomass proportion;growth characteristics;coastal wetland
October 9,2010
P737.17
A
1671-6647(2012)01-0132-09
2010-10-09
国家自然科学基金——互花米草对苏北盐沼沉积物有机碳库功能格局影响机理(40973053);江苏省高校自然科学重大基础研究项目——江苏省海滨湿地外来种互花米草扩张策略研究(08KJA180002)
徐伟伟(1986-),男,安徽芜湖人,硕士研究生,主要从事水环境生态学方面研究.E-mail:lovelock_312@hotmail.com
*通讯作者,女,副教授,主要从事海滨系统生态学方面研究.E-mail:liujine@njnu.edu.cn
(陈 靖 编辑)
致谢:陈正勇、宋康、吴晓东和张小飞等人协助采集样品和实验室工作。
猜你喜欢
海洋通报(2022年5期)2022-11-30 12:03:58
海洋通报(2022年5期)2022-11-30 12:03:48
安徽农业科学(2022年14期)2022-08-04 02:46:28
海洋通报(2021年5期)2021-12-21 06:20:02
现代园艺(2021年23期)2021-12-01 07:47:44
海洋通报(2021年1期)2021-07-23 01:55:40
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
海洋通报(2020年3期)2020-10-20 09:26:46
福建林业(2015年3期)2015-09-03 10:49:47
