
2010年北太平洋海区巴特柔鱼(Ommastrephebartrami)的生物学特征*
2012-01-05 11:27:48唐峰华伍玉梅
海洋科学进展 2012年2期
唐峰华,伍玉梅,樊 伟*
(1.中国水产科学研究院 东海水产研究所,上海200090;2.农业部东海与远洋渔业资源开发利用重点实验室,上海200090)
2010年北太平洋海区巴特柔鱼(Ommastrephebartrami)的生物学特征*
唐峰华1,2,伍玉梅1,2,樊 伟1,2*
(1.中国水产科学研究院 东海水产研究所,上海200090;2.农业部东海与远洋渔业资源开发利用重点实验室,上海200090)
根据2010-06—09北太平洋海区柔鱼(Ommastrephebartrami)的现场调查资料,对其群体组成、胴长与体重的关系、性腺成熟度、摄食等级和渔获产量变化作了初步研究。结果表明:2010-06东部海区(170°~178°W,38°~41°N)优势胴长组为280~300mm,优势体重组为800~1 000g,性成熟度以Ⅲ,Ⅳ和Ⅴ期为主,摄食等级5级均有分布;2010-07—09西部传统渔场海区(150°~157°E,38°~44°N)优势胴长组为200~260mm,体重组为200~600g,柔鱼各项生长指标呈逐日上升趋势,性成熟度以Ⅰ和Ⅱ期为主,摄食等级都以2级以上为主。柔鱼性成熟度群体百分比与高摄食强度呈显著的负相关,即性成熟度越高,摄食等级越低;性成熟度越低,摄食等级越高。
北太平洋;巴特柔鱼;性腺成熟度;摄食等级
(高 峻 编辑)
头足类是重要的海洋经济动物,广泛分布于太平洋、大西洋、印度洋和南极等海域。20世纪70年代初,由于北太平洋褶柔鱼(Todarodespacificus)在太平洋西侧的产量锐减,北太平洋的巴特柔鱼(Ommastrephe bartrami,简称北太平洋柔鱼)逐渐成为鱿钓渔业的新增对象。随着柔鱼加工需求增长,以及鱿钓生产向海外渔场拓展,其逐渐成为太平洋海区渔业的主要捕捞对象[1-2]。1993年以来,我国已有众多渔业公司进入北太平洋进行柔鱼鱿钓生产,收获颇丰。中国的鱿钓业起步较晚,许多渔业捕捞技术问题尚未解决,且对柔鱼生物学特征方面的基础研究并不完备。国内学者对北太平洋柔鱼资源生物学特征等方面进行了研究多集中于2000年之前,如早期郑元甲和严利平[3]、孙满昌等[4]、陈新军等[5]、黄洪亮和郑元甲[6]等等。目前急于研究探讨的问题是北太平洋海域柔鱼资源及渔业生物学特征现状如何?根据中国水产舟山海洋渔业公司北太平洋鱿钓生产船“1301”号,于2010-06—09在捕捞作业期间所采集的样本进行生物学特征现场测定,对柔鱼的生长、摄食、繁殖等生物学特征进行研究,为更好地开发北太平洋柔鱼资源提供参考和依据。
1 材料与方法
1.1 研究区域和调查站位
柔鱼样本为2010-06—09中国水产舟山海洋渔业公司生产船“1301”号在北太平洋公海海域进行的鱿钓生产所得。其中2010-06鱿钓生产集中在东部海区(170°~178°W,38°~41°N);2010-07—09集中在西部传统渔场海区(150°~157°E,38°~44°N),每月随机采样6~10个站位,各站位的随机取样为10~20个(符合统计学意义),样品全部进行现场生物学测定(站位见图1)。
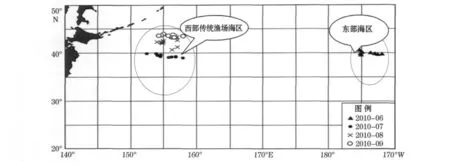
图1 北太平洋柔鱼渔场调查站位Fig.1 Sampling stations of the fishing grounds of neon flying squid in the North Pacific Ocean
1.2 生物学测定
1)海上现场调查和生物学测定依据《海洋监测规范》[7]进行,测量内容:胴长(L)、体重(W)、性别、性成熟度和摄食等级。其中胴长测定精确至1mm,重量精确至1g(海上渔船摇摆,采用手提电子称“KYPS08”定位读数)。
2)性腺成熟度等级划分参照Ehrhardt的方法[8],将性腺成熟度划分为Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ五期,将性腺成熟度达到Ⅰ和Ⅱ期的定为性未成熟群体,Ⅲ,Ⅳ和Ⅴ期的定为性成熟群体。
3)摄食强度根据相关参考文献[9],分为5级:0级为空胃;1级为胃内有少量食物,其体积不超过胃腔的1/2;2级为胃内食物较多,其体积超过胃腔的1/2;3级为胃内充满食物,但胃臂不膨胀;4级为胃内充满食物,胃臂膨胀变薄。
1.3 数据处理
1)调查数据用SPSS 12.0软件进行分析处理,用Excel 2003软件和SPSS软件运算并绘图。主要参数[10-12]的计算公式为
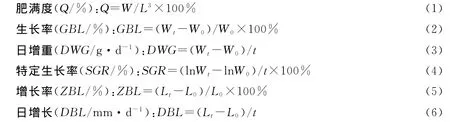
而柔鱼胴长与体重的关系式、体重的生长曲线分别为

式中,t为时间段(d);W为样品平均体重(g);Wt,W0分别为终末和初始平均体重(g);L为样品胴长(mm);Lt和L0分别为终末和初始平均胴长(mm);a,b,c,d为常数。
2)采用SPSS软件中的单因子方差分析法(ANOVA)分别对各月份柔鱼胴长与体重、胴长与性成熟度的差异显著性进行检验;并运用t检验方法对不同月份摄食等级的显著性差异进行检验。
2 结果与分析
2.1 结构特征
调查期间共测定北太平洋柔鱼438个,其中雌性263个,雄性175个。雌、雄个体的平均胴长分别为255和254mm;平均体重分别为587和582g。各月北太平洋柔鱼雌性个体的比例均大于雄性,性比在0.52~0.78波动。肥满度在东部海区显示较高水平;在西部传统渔场海区整体偏低,2010-07—09逐月上升(表1)。
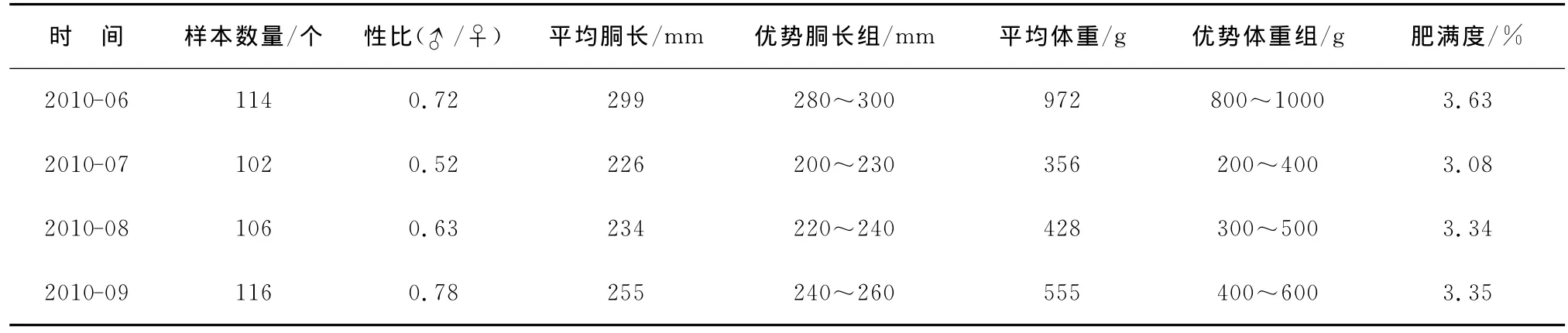
表1 柔鱼的样品Table 1 Sample situation of neon flying squid
2.2 胴长与体重的关系拟合
分别对东部海区和西部传统渔场的北太平洋柔鱼胴长和体重进行拟合,呈幂函数变化,且关系显著性明显。即东部海区(2010-06)W=1×10-5L3.1758,R2=0.984 0;西部传统渔场海区(2010-07—09)W=5×10-6L3.3433,R2=0.953 7。对调查期间柔鱼分雌、雄的胴长和体重拟合关系如图3,其中雌性:W=9×10-6L3.2372,R2=0.977 7;雄性:W=7×10-6L3.2731,R2=0.975 3。
2.3 生长参数
2010-07—09西部传统渔场海区的柔鱼生长情况分析表明。2010-07—08柔鱼的生长率为20.22%,特定生长率为0.59%;2010-08—09柔鱼的生长率为29.67%,特定生长率为0.87%。2010-07—09北太平洋柔鱼生长参数见表2,柔鱼体重随着生长时间的变化曲线(图4),呈显著的指数函数关系,即生长的体重W=25.182 3e0.0339t,该时间段柔鱼的各生长指标都是逐日上升的,尤其体重随着时间推移大幅上升,可推断这是柔鱼快速生长的旺盛期。
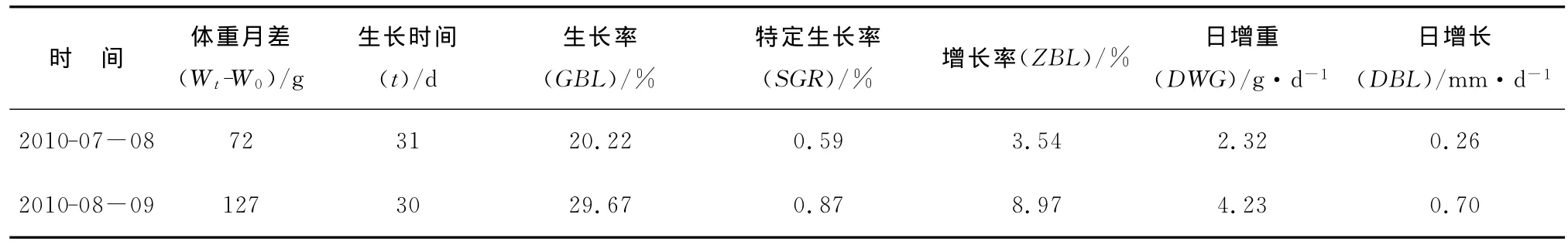
表2 西部传统渔场海区柔鱼的生长参数Table 2 Growth parameters of neon flying squid in western area

2.4 性成熟度
本次调查中2010-06东部海区统渔场成熟率(Ⅲ,Ⅳ和Ⅴ期)高达85.09%,2010-07—09西部传统渔场海区渔场成熟率逐月升高,依次分别为8.82%,12.26%,37.93%。调查期间各月柔鱼各性成熟度所占的比例如图5所示。而整个调查期间,性成熟度Ⅰ和Ⅱ期的个体比例最高,Ⅴ期比例最低。初步说明2010-06东部海区北太平洋柔鱼以生殖成熟群体为主,2010-07—09在西部海区柔鱼以未成熟群体为主。根据成熟率由12.26%大幅上升到37.93%,初步推断2010-08—09应是柔鱼的快速发育成长期。
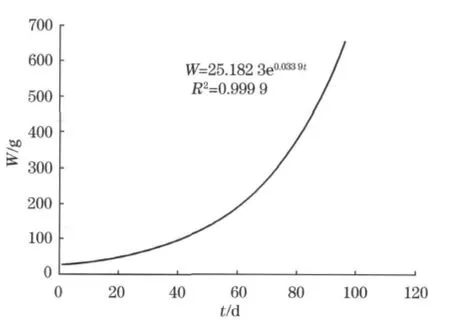
图4 2010-07—09西部传统渔场海区柔鱼体重和生长时间的关系Fig.4 The relationship between weight and growth time of neon flying squid in western area from July to September,2010
2.5 摄食等级
2010-06—09北太平洋柔鱼5个摄食等级均有分布,空胃(0级)和满胃(4级)的情况较少。2010-06东部海区摄食等级较均匀,1级较多,其次2级,接着依次为3,0,4级,分别为32.46%,29.82%,20.18%,14.04%,3.51%。2010-07—09摄食等级分布情况基本相同,2级比例最高,依次为2级>1级>0级>3级>4级,西部传统渔场海区柔鱼的摄食强度明显高于2010-06的东部海区,饵料生物优越。t检验方法显示除2010-06外,2010-07—09摄食等级差异均存在显著性(P<0.01),且最多的2级比例逐月降低(图6)。
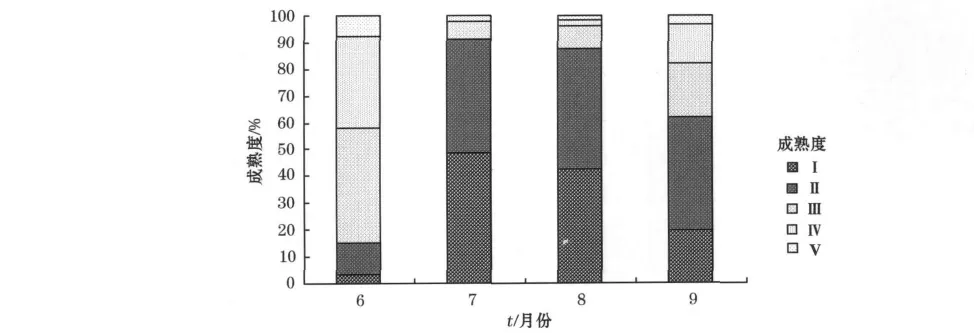
图5 柔鱼各性成熟度百分比Fig.5 Percentage of each sexual maturity degree of neon flying squid
通过对摄食等级较高的柔鱼胃含物成份的分析,发现柔鱼的饵料成份由头足类、鱼类和甲壳类组成。从饵料成份的出现机率来看,以头足类最高,在90%以上;鱼类次之,甲壳类最少。由于饵料在柔鱼胃内的消化程度较高,达2级和3级,因此对进一步分析饵料的种类有一定的困难。
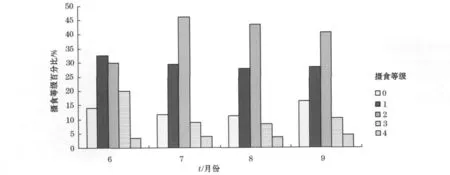
图6 北太平洋柔鱼摄食等级变化Fig.6 Change of feeding levels of neon flying squid
2.6 性成熟度与摄食强度的关系
东部海区(2010-06),柔鱼的性成熟群体比例较高,而西部传统渔场海区(2010-07—09),逐月的性成熟比例依次升高。所以根据2010-07,08,09,06柔鱼性成熟群体(Ⅲ,Ⅳ和Ⅴ期)百分比依次升高的顺序(假设2010-06在东部海区柔鱼视为“老年化”的稳定群体)和对应阶段的高摄食强度(2,3和4级)百分比关系进行拟合(图7),初步可以看出:性成熟度越低,摄食等级就高;性成熟度越高,摄食等级越低;二者呈明显的负相关。
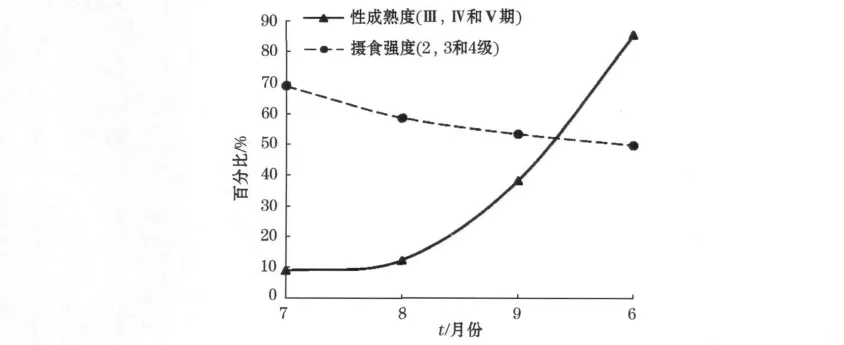
图7 性成熟度和摄食强度的关系Fig.7 Relationship between gonad maturities(Ⅲ,Ⅳ,Ⅴ)and feeding stages(2,3,4)
3 讨 论
3.1 不同海区柔鱼群体的分析
已往研究资料中,西部传统渔场海区优势胴长组最大为1999年的190~360mm,占88.44%,平均胴长为282.84mm;东部海区优势胴长组最大为1998年的380~400mm,占47.5%,平均胴长为396.80 mm[13]。本次调查中,2010-06东部海区(非传统渔场)优势胴长组为280~300mm,体重组为800~1 000 g;2010-07—09西部传统渔场海区优势胴长组为200~260mm,体重组为200~600g,而且3个月平均胴长和体重依次逐月升高,摄食等级都以2级为主,这说明处于北太平洋柔鱼的生长发育时期,该海域是其重要的索饵场;这与本次调查主要作业范围纬度逐渐升高(38°~44°N)可能有关(图1),纬度越高鱼体胴长有增加的趋势。与以往同期比较,总体柔鱼个体呈小型化趋势。本次调查中北太平洋柔鱼胴长和体重关系呈幂函数变化,分东部海区和西部传统渔场海区拟合,其关系式分别为W=1×10-5L3.1758和W=5×10-6L3.3433,关系显著性明显。
东部海区柔鱼胴长明显大于西部传统渔场海区的。从本次柔鱼群体组成看出,北太平洋东部和西部可能存在2个明显不同的群体。柔鱼为一年生,并分为中部秋生群、东部秋生群、西部冬春生群和中东部冬春生群共4个群体,其中西部冬春生群为中国鱿钓船的传统捕捞对象,主要捕捞区域为38°~46°N,165°E以西海域,即2010-07—09调查的西部传统渔场海区;170°E以东为东部海区该海域的柔鱼为东部秋生群[14-16]。东部海区体重与胴长的关系式b参数(3.175 8)明显小于西部海区体重与胴长的关系式b参数(3.343 3)的值(图2),根据指数的函数关系说明东部海区柔鱼体重随胴长的变化更趋向于平稳,而西部传统渔场海区柔鱼体重随着胴长增加而上升得更明显;同时根据2海区的柔鱼的性成熟度比较(图5)表明,东部海区北太平洋柔鱼以生殖成熟群体为主,西部传统渔场海区柔鱼以未成熟群体为主;东部海区和西部传统渔场柔鱼的摄食等级(图6)显示,西部传统渔场海区柔鱼的摄食强度明显高于东部海区的。根据这些比较,初步判断东、西部海区柔鱼从生态特征上有一定的区别和差异。但是东、西部海区的柔鱼是否属于不同群体,光从形态学[17]上区分是不够的,还需要进一步分析其成熟度随时间的变化及利用耳石信息分析技术[18-19]来确定,最好使用微观分子生物学测定其DNA[20]进行判定。
3.2 不同时间的差异分析
2010-07—09和对应的西部传统渔场海区(150°~157°E)是北太平洋柔鱼摄食的高峰期和重要的索饵场,该时期的空胃率仅为11%~16%,胃饱和度主要以2级为主。根据对各月性成熟度(Ⅲ、Ⅳ和Ⅴ期)百分比和对应阶段的摄食强度(2,3和4级)百分比呈显著的负相关(图7),初步推断摄食等级的组成有可能与渔汛期的盛衰有关,随着渔汛期的开始到旺盛到后期的衰落,柔鱼群体的性成熟比例逐渐升高,从生长发育到成熟稳定,柔鱼的胃口也越来越小,最后趋于低摄食等级的稳定状态。柔鱼的摄食强度与性成熟度的呈负相关关系,尚不能完全下定论,最好以2010-09—2011-06连续性的调查,获取柔鱼性成熟度与摄食等级完整的相关数据以此来证明。
2010-07—09西部传统渔场海区连续性的调查结果显示,该时期柔鱼的各项生长参数值逐渐上升,尤其是体重随着生长时间的推移呈指数增长,显著性非常明显。本次调查时间有限,但初步判定2010-06—09柔鱼的月平均产量逐渐上升趋势,而且西部传统渔场海区的产量显然高于东部海区的。2010-06东部海区170°~178°W水表温较低,钓线一般放至200m以下作业,深水捕捞使鱿钓效率大大降低。该海域低产量原因:1)海域广阔,柔鱼资源分布不集中;2)水温较低,柔鱼活动层处于200~400m,水位较深;3)该海域的柔鱼性成熟度以Ⅲ期以后的为主,活动能力不强。2010-07—09在西部传统渔场海区150°~157°E范围内,水表温逐月上升,到了2010-07中下旬基本在18℃以上,钓线放至30mm即可作业,捕捞效率提高;2010-08—09是柔鱼的盛渔汛期,该海域海表温急剧上升,稳定在20℃左右,为柔鱼重要的生长索饵场和快速生长的一个阶段,所以随着鱼群聚集,2010-07—09平均产量上升逐渐。
4 结 语
利用2010-06—09的渔场调查数据,对东部海区(170°~178°W,38°~41°N)和西部传统渔场海区(150°~157°E,38°~44°N)的柔鱼群体的结构特征及资源状况进行研究。4个月的调查结果表明,东部海区柔鱼胴长、体重明显大于西部海区的;西部传统渔场海区柔鱼群体的资源量在2010-06—09是逐月升高的;2010-07—09是西部渔场海区北太平洋柔鱼的一个主要生长时期。因调查时间所限,未能对渔汛后期10和11月北太平洋柔鱼的生殖群体进行研究。后续的工作,还需要结合各群体的分布特点及产卵、洄游等习性,进行长时间、大范围的连续取样,以对柔鱼繁殖习性方面进行分析研究。另外,考虑到北太平洋渔场作业的外部环境因子,柔鱼资源状况受海洋环境因子及捕捞作业强度等因素的影响,还需要对北太平洋柔鱼进行后续的取样调查和结合海洋环境数据的分析,才能对北太平洋柔鱼的群系组成资源量在整个渔汛期的变化幅度和变化趋势做更深入的探讨。
(References):
[1]DONG Z Z.Biology of the economic species of cephalopods in the world oceans[M].Ji’nan:Shangdong Science and Technology Press,1991:82-118.董正之.世界大洋经济头足类生物学[M].济南:山东科学技术出版社,1991:82-118.
[2]WANG R G,CHEN X J.Economy soft fish resources and fishing in the world oceans[M].Beijing:China Ocean Press,2005:80-152.王尧耕,陈新军.世界大洋性经济柔鱼类资源及其渔业[M].北京:海洋出版社,2005:80-152.
[3]ZHENG Y J,YAN L P.Composition characteristics of soft fish group in North Pacific Sea in 1999[J].Pelagic Fisheries,2000,3:7-14.郑元甲,严利平.1999年北太平洋海区柔鱼群体组成特征[J].远洋渔业,2000,3:7-14.
[4]SUN M C,ZHANG S H,QIAN W G.Approach on biological characteristics ofOmmastrephesbartramiion the central and eastern squid fishing grounds in the North Pacific[J].Marine Fisheries,2001,23(1):21-24.孙满昌,张圣海,钱卫国.北太平洋中东部海域的柔鱼生物学特征研究[J].海洋渔业,2001,23(1):21-24.
[5]CHEN X J,TIAN S Q,YE X C.Study on population structure of flying squid in Northwestern Pacific based on gray system theory[J].Journal of Shanghai Fisheries University,2002,11(4):335-341.陈新军,田思泉,叶旭昌.西北太平洋柔鱼种群的聚类分析[J].上海水产大学学报,2002,11(4):335-341.
[6]HUANG H L,ZHENG Y J.Analyses on the biological characteristics of squid in the North Pacific[J].Marine Fisheries,1998,20(3):106-110.黄洪亮,郑元甲.北太平洋海区柔针生物学特征分析[J].海洋渔业,1998,20(3):106-110.
[7]GB17378-2007Specification for marine monitoring[S].Beijing:Chinese Standard Press,2008.GB17378-2007海洋监测规范[S].北京:中国标准出版社,2008.
[8]EHRHARDT N M,JACQUENMIN P,GARCIA F,et a1.On the fishery and biology of the giant squidDosidicusgigasin the Gulf of California,Mexico[R].FAO.Advances in assessment of world cephalopod resources.FAO Fisheries technical paper NO.231.Rome:[s.l.]1983:306-320.
[9]GB12763.6-2007Specifications of oceanographic surrey:Part 6Marine Biologica Survey[S].Beijing:China Standard Press,2008.GB 12763.6-2007海洋调查规范:第6部分海洋生物调查[S].北京:中国标准出版社,2008.
[10]WANG Y Y,HUO Y Z,CAO J C,et al.Influence of low temperature and low light intensity on growth ofUlvacompessa[J].Journal of Fishery Sciences of China,2010,17(3):593-599.王阳阳,霍元子,曹佳春,等.低温、低光照强度对扁浒苔生长的影响[J].中国水产科学,2010,17(3):593-599.
[11]ZHANG Y Q,YIN J S,WANG B Q,et al.Effects of stocking density on growth,survival and behavior of juvenile Siberian charrSalvelinusleucomaenis[J].Journal of Dalian Fisheries University,2009,24(6):520-524.张永泉,尹家胜,王丙乾,等.养殖密度对白点鲑幼鱼生长、存活以及行为的影响[J].大连水产学院学报,2009,24(6):520-524.
[12]CHEN X J.The fishery resources and fishery ground[M].Beijing:China Ocean press,2004:46-83.陈新军.渔业资源与渔场学[M].北京:海洋出版社,2004:46-83.
[13]HUANG H L,ZHENG Y J,CHEN J H.Study on the biological characteristics ofOmmastrephebartramiin the North Pacific[J].Marine Fisheries,2003,25(3):126-129.黄洪亮,郑元甲,程家骅.北太平洋海区柔鱼生物学特征研究[J].海洋渔业,2003,25(3):126-129.
[14]YATSU A,MIDORIKAWA S,SHIMADA T,et al.Age and growth of the neon flying squid,Ommastrephesbartrami,in the North Pacific Ocean[J].Fish.Res.,1997,29:257-270.
[15]YATSU A,TANAKA H,MORI J.Population structure of the neon flying squid,Ommastrephesbartramii,in the North Pacific[C]∥OKUTANI T.Contributed papers to International Symposium on Large Pelagic Squids.Tokyo:Japan Marine Fishery Resources,1998:31-48.
[16]NAGASAWA K,MORI J,OKAMURA H.Parasites as biological tags of stocks of neon flying squid(Ommastrephesbartramii)in the North Pacific Ocean[C]∥OKUTANI T.Contributed Papers to International Symposium on Large Pelagic Squids.Tokyo:Japan Marine Fishery Resources,1998:49-64.
[17]LI S L,CHEN X J,LIU B L,et al.Morphological analysis on population structure of neon flying squid(Ommastrephebartrami)in the Northwest Pacific Ocean[J].Periodical of Ocean University of China:Natural Science,2010,40(3):43-48.李思亮,陈新军,刘必林,等.基于形态法的西北太平洋柔鱼种群结构研究[J].中国海洋大学学报:自然科学版,2010,40(3):43-48.
[18]MA J,LIU B L,CHEN X J,et al.Advances on using statolith to study age and growth of cephalopod[J].Marine Fisheries,2009,31(3):316-324.马 金,刘必林,陈新军,等.利用耳石鉴定头足类年龄与生长研究进展[J].海洋渔业,2009,31(3):316-324.
[19]YANG L L,CHEN J H.Research progress on information analysis of statoliths in cephalopods[J].South China Fisheries Science,2009,5(3):74-80.杨林林,程家骅.头足类耳石信息分析的研究进展[J].南方水产,2009,5(3):74-80.
[20]FENG J B,WU C L,DING H Y,et al.Microsatellite analysis of genetic diversity in 9wild stocks of oriental river prawnMacrobrachiumnipponensein Hongze Lake[J].Journal of Fishery Sciences of China,2010,17(2):218-227.冯建彬,吴春林,丁怀宇,等.洪泽湖日本沼虾9个野生群体遗传多样性微卫星分析[J].中国水产科学,2010,17(2):218-227.
Biological Characters of Neon Flying Squid(Ommastrephebartrami)in the North Pacific Ocean
TANG Feng-hua1,2,WU Yu-mei1,2,FAN Wei1,2
(1.EastChinaSeaFisheriesResearchInstitute,CAFS,Shanghai 200090,China;2.KeyLaboratoryof EastChinaSeaandOceanicFisheryResourcesExploitation,MinistryofAgriculture,Shanghai 200090,China)
Based on the survey data of neon flying squid(Ommastrephebartrami)fishery in the North Pacific Ocean from May to September in 2010,the population composition,the relationship between mantle length and body weight,gonad maturity,feeding stages and amount of harvest were investigated.Results show,in June in the eastern sea area(170°~178°W,38°~41°N),280~300mm was the dominant mantle length,800~1 000g was the dominant body weight,gonad maturity was at stageⅢ,Ⅳ,Ⅴ,feeding stages were distributed from 0to 4class;from July to September in the western sea area(150°~157°E,38°~44°N),200~260mm was the dominant mantle length,200~600g was the dominant body weight.During this period,the growth parameters were raising day by day.Gonad maturity was mainly at stageⅠandⅡ,feeding stages were primarily above classes 2.There was a significant negative correlation between sexual maturity degree of squid and high intensity of feeding.The higher degree of maturity,the lower feeding level was and vice versa.
North Pacific Ocean;Ommastrephebartrami;gonad maturity;feeding stages
November 11,2011
S931.5
A
1671-6647(2012)02-0257-09
2010-12-30
国家高技术研究发展计划——大洋渔场渔情信息应用技术系统开发(2007AA092202)
唐峰华(1982-),男,浙江湖州人,助理研究员,主要从事海洋生态学和渔业资源方面研究.E-mail:f-h-tang@163.com
*通讯作者,研究员,E-mail:fanwee@126.com
致谢:中国水产舟山海洋渔业公司远海渔业部门陆良峰经理和朱金鑫。
猜你喜欢
海洋信息技术与应用(2021年3期)2022-01-17 02:37:26
中国水产(2021年12期)2021-12-06 11:14:00
当代水产(2021年1期)2021-03-19 05:17:14
大自然探索(2021年12期)2021-02-07 01:37:04
当代水产(2019年9期)2019-10-08 08:02:44
当代水产(2019年4期)2019-05-16 03:04:20
甘肃畜牧兽医(2019年2期)2019-03-20 01:49:42
建筑热能通风空调(2018年5期)2018-07-09 03:16:44
渔业致富指南(2016年12期)2016-11-11 07:07:24
海洋渔业(2016年6期)2016-04-16 03:00:23
