
行距对老芒麦种群构件组成、生物量结构及能量分配的影响
2012-01-02 02:48刘金平游明鸿白史且
草业学报 2012年3期
刘金平,游明鸿,白史且
(1.西华师范大学生命科学院,四川 南充637009;2.四川省草原科学研究院,四川 成都611731)
*老芒麦(Elymussibiricus)是禾本科披碱草属多年生疏丛型禾草[1],“川草2号”老芒麦(E.sibiricuscv.chuancao No.2)是目前我国青藏高原东缘广泛种植的一个优良牧草品种[2,3],为该地区生态保护、草地改良、退牧还草、种草养畜以及人草畜三配套等工程中的骨干牧草品种。目前,川西高原建有3 500hm2种子基地,近100 000hm2人工草地,由于高原特殊的自然条件与经营条件,生产中缺乏相关技术支撑,种子产量极低、鲜草产量极不稳定[4,5],远远不能满足高寒牧区对老芒麦种子与草产品的需要。开展适合高寒牧区优质牧草与种子生产的技术研究,提高生产的技术含量,是高寒牧区畜牧业发展急需解决的生产实践问题。
自20世纪70年代初Harper和White[6]提出植物种群构件结构理论以来,种群构件的研究方法在蕨类[7]、一年生草本[8,9],灌木和木本[10,11]及无性系植物[12,13]上进行了应用。构件数量与性状不仅决定种群同化作用的能力,对植株生物量分布、能量积累及抗逆性都有极其重要的作用。目前,老芒麦的研究多集中于资源评价、品种选育及栽培利用、草产量动态及适应性等研究[14-20],关于行距单因子对老芒麦种群构件组成、生物量结构及能量分配的影响未见报道。试验通过对不同行距,第3年盛花期老芒麦种群的根、茎、叶、花序等构件数量性状、生物量结构、能量累积及分配的比较分析,探讨适合川西北高原饲草与种子生产的合理行距,以及提高产量与质量的途径,以期为该区畜牧业发展、农业产业结构调整和生态环境治理等提供服务。
1 材料与方法
1.1 试验地概况
试验于阿坝州红原县邛溪镇二农场进行,为大陆性高原温带季风气候,东经102°32′、北纬32°46′、海拔3 497 m,年均温1.1℃,极端高温23.5℃,极端低温-33.8℃,年降水量738mm,相对湿度71%,≥10℃年积温仅865℃。土壤为草甸土,0~20cm 土壤的有效氮、有效磷、有效钾含量分别为276mg/kg,10.2mg/kg,131 mg/kg,有机质含量5.87%,pH 值5.91[4]。
1.2 试验材料及设计
以川草2号老芒麦原种为材料。以3m×5m=15m2,间距1m建立小区。按随机区组3次重复排列,以30 cm(18行)、45cm(11行)、60cm(8行)、75cm(6行)、90cm(5行)为行距,按100粒/m播种量播种。播后同样水肥、除杂等相同水平管理。播后第3年进行测定。
1.3 测定指标及方法
构件组成:盛花期,在小区中间随机取50cm长的样段,齐地刈割地上部分,称鲜重后,摊薄均匀选取1kg鲜草,3次重复。每份鲜草中随机选取50个生殖枝,测定顶端数第3片成熟叶片长度与叶中部宽度为叶长、叶宽,单枝叶片数为单蘖叶数;测定枝条长度、节间长、节直径、茎直径、单枝节数;测定花序长度、花序宽度、花序厚度、小穗数、小花数;测定生殖枝数及所占比例。而后把1kg鲜草的叶、茎、花序分离,装布袋中。按50cm(长)×30 cm(宽)×30cm(高)挖取茎基与根系,测定根系深度、分布范围后冲洗泥土后装袋。
生物量结构:分离装袋的叶、茎、花序在105℃下烘至恒重后称重,计算每hm2构件生物量,其和为地上生物量总量;把茎基与根系分开后,在105℃下烘至恒重后称重,计算每hm2构件生物量,其和为地下生物量总量。
能量分配:把烘干叶、茎(含茎基)、花序、根系粉碎后,用parr1281氧弹量热仪检测单位重量构件热能值,计算每hm2构件能量值。
1.4 数据处理
采用SAS 9.1进行多重比较、方差分析等数据处理。
2 结果与分析
2.1 行距对老芒麦构件组成的影响
2.1.1 行距对叶数量性状的影响 叶是光合作用的主要器官,其性状与数量对植物同化作用大小、构件形成、生物量积累与分配起决定性作用。试验中尽管叶片性状有一定的差异,其变异系数叶长>叶宽>单蘖叶数(表1),但行距对叶长、叶宽、单蘖叶片数影响均不显著。
2.1.2 行距对根数量性状的影响 根系深度及分布范围决定植株吸收范围与吸收能力,也决定植物的适应性与抗逆性,行距对根系深度有显著影响(P<0.05)(表1),随着30~75cm行距增加根系深度增加,但差异不显著,90cm行距根系显著深于其他行距。行距极显著影响根系分布范围(P<0.01),不同行距间有明显的差异,行距越大,根系分布越广。
2.1.3 行距对分蘖数的影响 在单位长度播量一定下,不同行距提供植株的空间、养分等不同,使单位面积营养枝数、生殖枝数、分蘖数间存在极显著不同(P<0.01)(表2)。小行距分蘖数显著高于大行距,30cm行距分蘖数高达2 628枝/m2,随着行距增加单位面积分蘖数减少,但行距大于60cm减少不显著。但单位面积营养枝数、生殖枝数与行距没有显著的线性关系。生殖枝比例显著受行距的影响(P<0.01),60cm行距单位面积生殖枝数所占比例最大,显著高于其他行距比例。
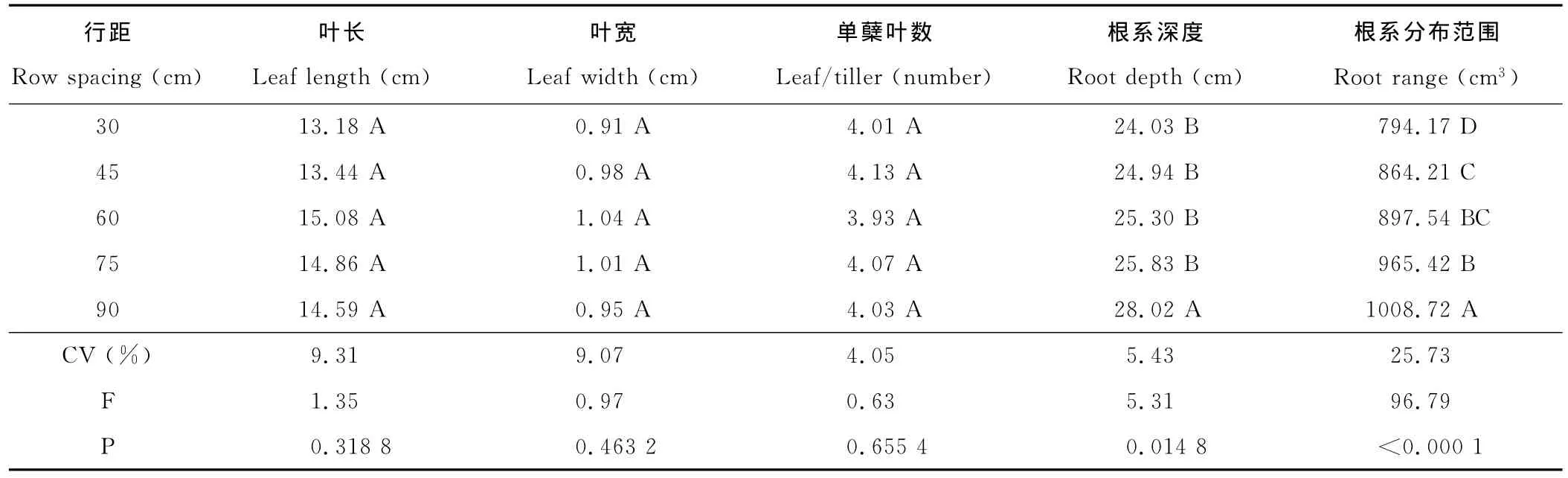
表1 行距对叶和根系数量性状的影响Table 1 Effect of different row spacing on quantitative traits of leaves and roots
2.1.4 行距对茎数量性状的影响 行距对决定株高的单蘖节数、节间长有显著影响(P<0.05),致使不同行距下老芒麦自然高度存在极显著差异(P<0.01)(表2)。30和45cm行距单位面积分蘖数高,枝条间对水肥与空间竞争使自然高度显著低于大行距处理,而行距大于60cm后,高度不再增加。行距对茎直径无明显影响,但小行距下对光照及空间竞争使枝条节直径明显低于大行距,行距越大节直径越粗,为营养物质贮藏与累积及抗逆性增加奠定基础。
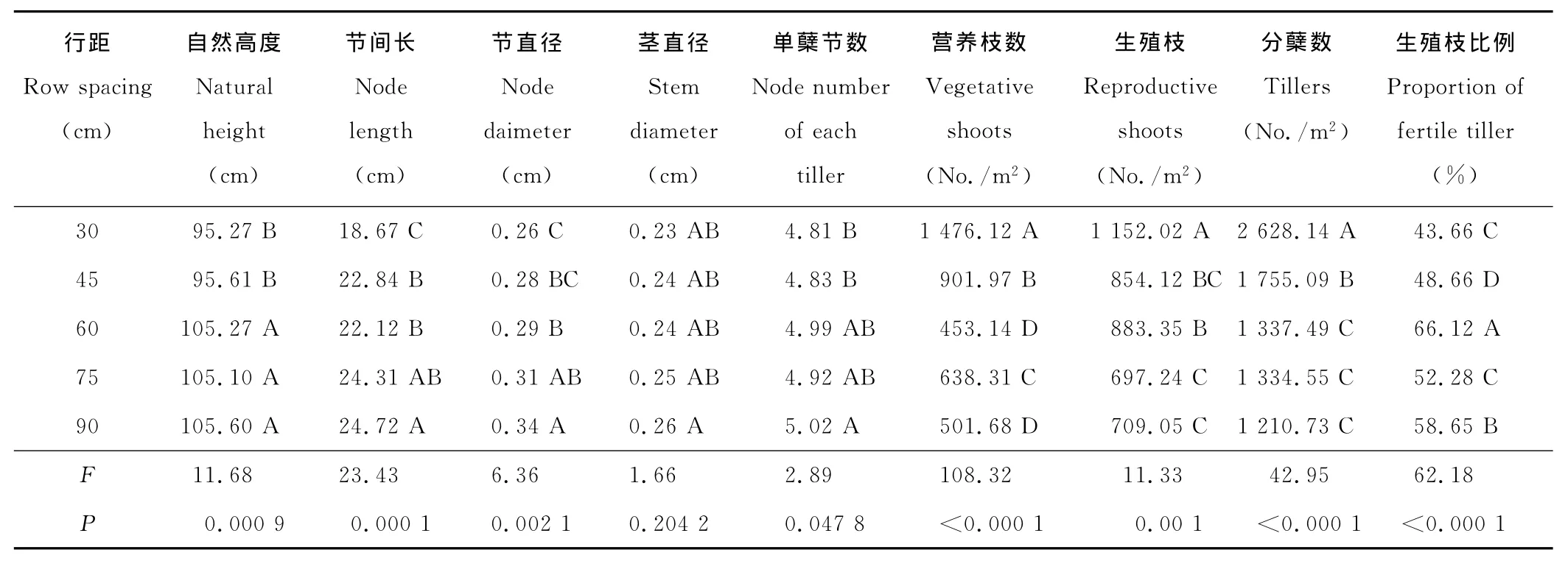
表2 行距对分蘖数及茎数量性状的影响Table 2 Effect of different row spacing on tillers and quantitative traits of stem
2.1.5 行距对花序数量性状的影响 行距对花序柄和花序轴有极显著影响(P<0.01),对花序柄长影响大于对花序柄直径与花序轴长的影响(表3)。柄长随行距增大而增长,柄直径随着柄长增长而增大。但行距超过60 cm,柄性状增加不明显,行距大于45cm后花序轴长不再增加。小穗数/生殖枝、小花数/生殖枝显著受行距的影响(P<0.05)。小穗数/生殖枝除30cm行距明显低、60cm明显高外,其他处理间差异不大。行距大于45cm后,对小花数/生殖枝影响不显著。

表3 行距对花序数量性状的影响Table 3 Effect of different row spacing on quantitative traits of inflorescence
2.2 行距对老芒麦生物量结构的影响
行距对老芒麦第3年鲜草产量有极显著影响(P<0.01)(表4),行距30~90cm下,单位面积植株行数相差近4倍,而鲜草产量变异范围为11 447~19 225.6kg/hm2,变异系数仅为0.69%。行距极显著影响老芒麦地上生物量与地下生物量总量(P<0.01),行距越小,地上与地下生物量累积均越大,地上生物量受行距影响大于地下生物量。
叶、茎、花序的生物量均显著受行距的影响(P<0.01),受影响程度是茎>叶>花序,三者共同作用使地上生物量间存在极显著差异。但90cm行距叶、茎生物量及地上生物量总和均显著高于75cm,这与老芒麦栽培年限、落粒性及自播能力强有很大关系。
行距对地下根系有显著影响(P<0.01),行距越大,根系生物量越小。行距越大越有利于单株根系的延伸与扩展,但老芒麦生物学特性与植株密度的差异,决定其根系分布能力有一定极限,致使大行距根系的生物量较低。行距对地下茎基影响极小(P>0.05),说明大行距植株密度虽小,但通过根茎的延伸使植株分蘖能力强于小行距,使单位面积地下茎基生物量间无显著差异。
2.3 行距对老芒麦能量累积的影响
2.3.1 行距对构件单位生物量能量值的影响 行距对叶、茎、根、花序的单位生物量能量值无显著影响(P>0.05)(表5),影响由大到小的顺序为叶>根>茎>花序,尤其30cm行距叶的能量值显著低于45~60cm行距下的值。单位生物量能量值不同构件间有差异,根>叶>花序>茎,表明老芒麦不同构件间营养价值的差异。
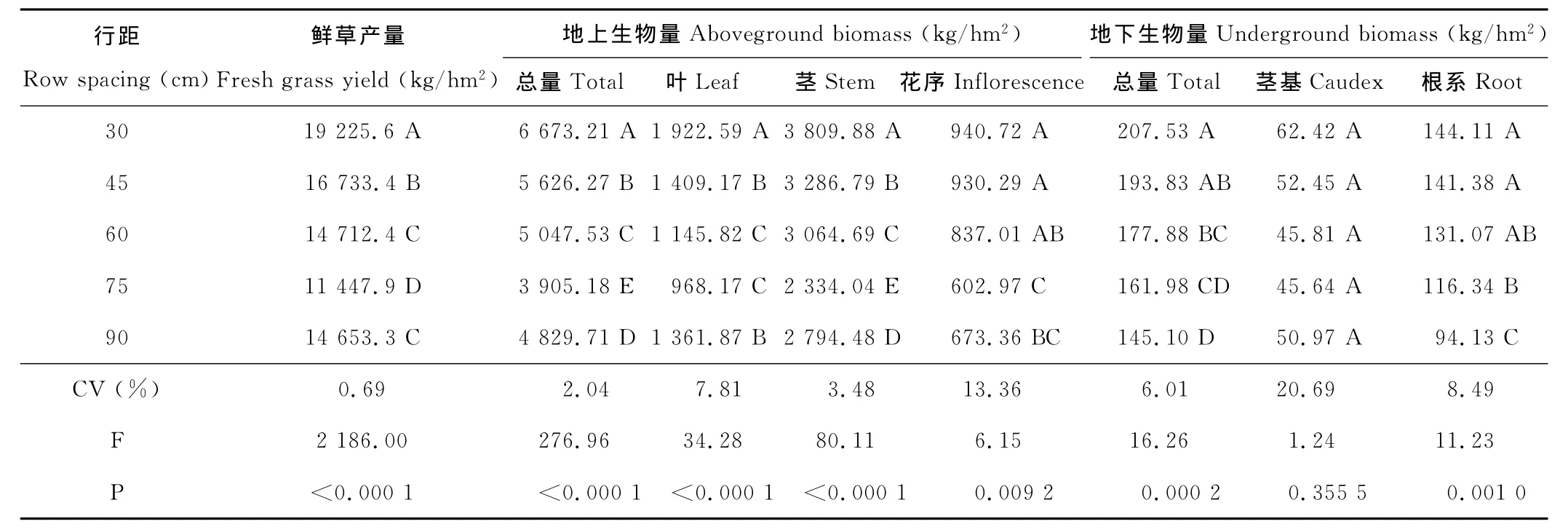
表4 行距对老芒麦生物量结构的影响Table 4 Effect of different row spacing on biomass structure of E.sibiricus
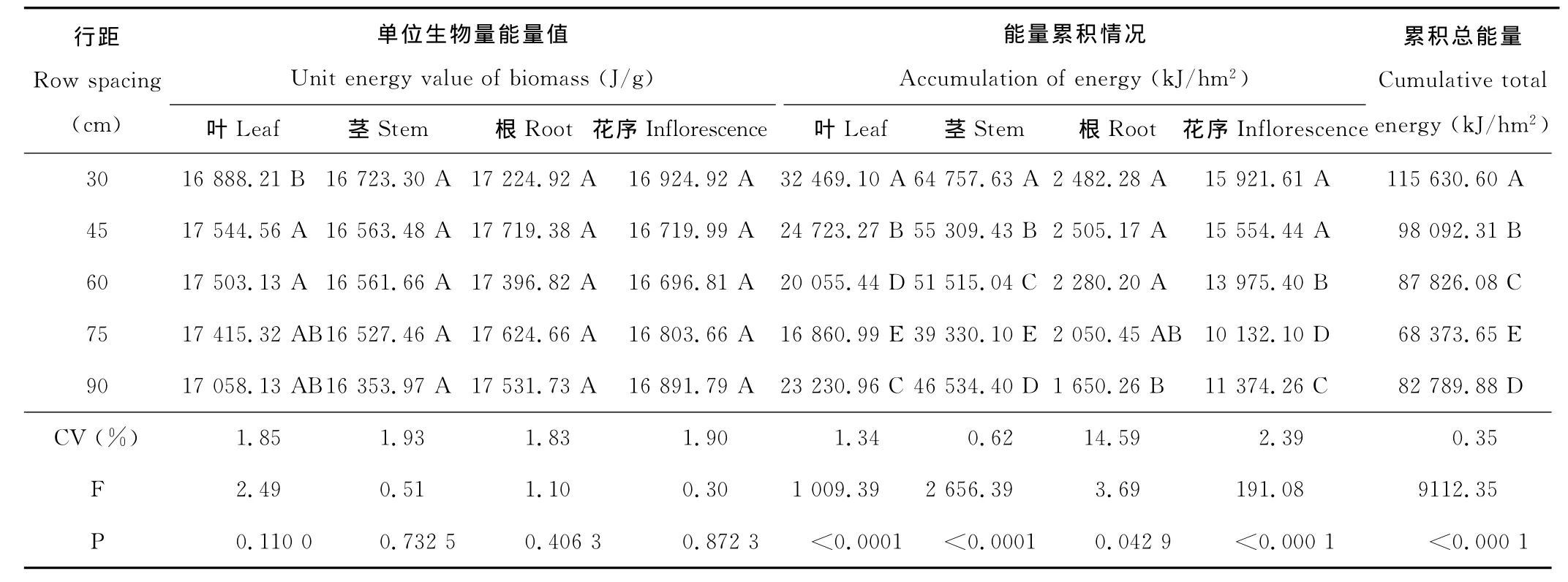
表5 行距对老芒麦能量累积的影响Table 5 Effect of row spacing on energy cumulation of E.sibiricus
2.3.2 行距对构件能量累积的影响 虽然行距对叶、茎、根、花序的单位生物量能量值影响不大,但由于行距对生物量结构的显著影响,致使行距对叶、茎、根、花序的能量累积有显著影响,影响由大到小的顺序为茎>叶>花序>根。行距对单位面积累积总能量也有极显著影响(P<0.01),行距30cm累积能量最大达115 630.6 kJ/hm2,随行距增大,叶、茎、根、花序累积能量逐渐减少,75cm 行距累积能量仅为68 373.65kJ/hm2,为30cm的59.13%。行距90cm由于空间大,容易受自播影响,使其能量累积大于75cm。
2.3.3 行距对构件能量分配的影响 行距极显著影响老芒麦能量在构件间的分配,使叶、茎、根、花序累积能量在不同行距下所占比例呈显著差异(表6)。构件能量分配受行距影响由大到小为叶>茎>花序>根。30cm行距的叶分配能量最大达28.08%;60cm行距的茎与花序分配能量最大分别达58.66%与15.91%;75cm行距的根分配能量最大达2.99%;90cm行距受自播小苗的影响使其叶的能量偏大于正常水平。
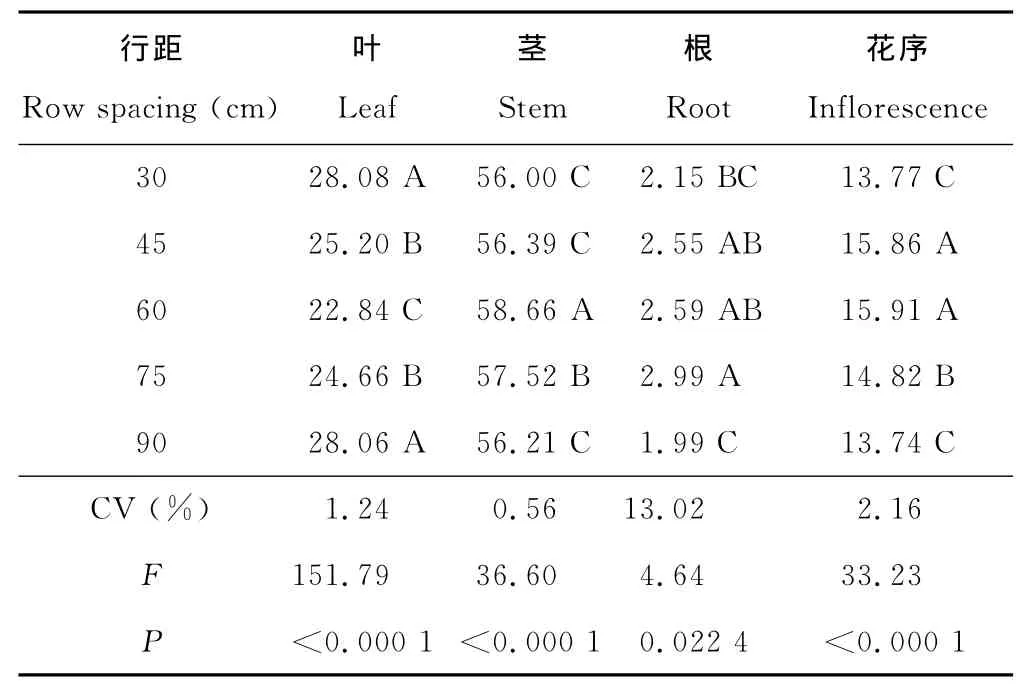
表6 行距对老芒麦能量分配的影响Table 6 Effect of row spacing on energy distribution of E.sibiricus %
3 结论与讨论
行距对老芒麦不同构件数量性状的影响不同。通过对叶、茎性状的影响,尤其对单位面积分蘖数、单蘖节数、节间长的显著影响,使枝条密度与高度表现为极显著差异。而密度与高度是决定禾本科牧草鲜草产量的第一、第二主要因子[2,3],导致行距对鲜草产量的显著影响。根系深度与分布范围直接影响植物的抗旱、抗寒及抗倒伏等抗逆能力,大行距有利于提高老芒麦的抗逆性能。生殖投入比例及花序数量性状的差异,使不同行距下老芒麦生殖投入量、种子潜在产量存在差异,造成开花授粉、种子发育、种子活力及落粒性等相关性状的物质基础不同。
行距不仅可以提高土地的利用率、调控草地的播种量与植株密度,同时也调节了单位植株可利用的水、肥、气、热、光照、空间等生态因子,从而影响根、茎、叶、花序等构件的数量与性状,导致植物种群的生物量结构发生显著的改变。试验表明,行距对构件生物量影响为茎>叶>花序>根,小行距有利于地上与地下生物量累积。打草地建设时,适当采用小行距,可以提高单位面积的鲜草产量。同时不适合种子生产,虽然其花序分配的生物量大,但其生殖枝发育受高密度、强竞争的影响较大,致使小穗数、小花数、授粉率、结实率远低于60cm行距。可见,生产中可以通过选择合理的行距,影响老芒麦的构件数量及性状,改变人工草地的构件结构与环境因子,为提高老芒麦鲜草产量、种子产量及抗逆性奠定基础。
不同行距下,老芒麦种群自然因子与生物因子都存在不同,引起构件数量性状的差异,必然影响老芒麦的同化作用速度及合成物质的能力。试验发现不同构件单位生物量能量值有差异,能量值由高到低为根>叶>花序>茎,但行距对叶、茎、根、花序的单位生物量能量值均无显著影响。说明行距虽然影响叶片面积与数量、茎的高度与粗细、根系的深度与分布、花序的数量及大小,必将影响植株对地下水肥的吸收能力、光合作用能力,对同化作用的产物种类与数量、物质转移及贮存能力也有影响,但各构件单位生物量能量值趋于稳定,说明同一构件不论糖、脂肪、蛋白质含量及存在形态有无差异,其单位生物量合成、贮藏或氧化释放的ATP几乎相等。随行距增大,单位面积的叶、茎、根、花序累积能量逐渐减少,使草地截获与贮藏的总能量降低。所以加强对环境因子以及限制性因子的分析,优化与改善生态因子为植物提供适宜的生长环境条件,是挖掘老芒麦种子生产潜力、提高产量的有效途径。
试验从高寒牧区产业化生产的实际技术需求出发,通过调控行距,实质是改变了水、肥、气、热、光照及生物等生态因子综合形成的生境条件,生境条件往往引起植株细胞分裂、组织分化、器官形成等微观过程的差异,致使同化作用、新陈代谢、能量分配不同,最终影响到植株形态、生物量结构及生产性能等外在指标,在其他植物上研究表明,每个生态因子对构件结构及生产性能都有影响[24],有待于在老芒麦上进行深入系统的研究。
[1]陈默君,贾慎修.中国饲用植物[M].北京:中国农业出版社,2002:121-122.
[2]鄢家俊,白史且,张新全,等.青藏高原东南缘老芒麦自然居群遗传多样性的SRAP和SSR分析[J].草业学报,2010,19(4):122-134.
[3]游明鸿,张昌兵,刘金平,等.行距和肥力对川草2号老芒麦生产性能的作用[J].草业科学,2008,25(2):69-72.
[4]游明鸿,刘金平,白史且,等.肥料和除草剂混施对老芒麦生产性能的影响[J].草业学报,2010,19(5):283-286.
[5]刘金平,游明鸿.肥料和除草剂混施对老芒麦构件组成及生物量结构的影响[J].中国草地学报,2010,32(4):42-48.
[6]Harper J L,White J.The demography of plants[J].Annual Review of Ecology and Systematics,1974,5:419-463.
[7]Abulfaith H A,Bazzaz F A.The biology ofAmbrosiatrifidaL.Ⅳ.Demography of plants and leaves[J].New Phytologist,1980,84:107-112.
[8]Bazzaz F A,Harper J L.Demographic analysis of the growth ofLinumusitassimum[J].New Phytologist,1977,78:29-52.
[9]何维明,董鸣.分蘖型克隆植物黍分株和基株对异质养分环境的等级反应[J].生态学报,2002,22(2):169-175.
[10]何丙辉,钟章成.同养分条件对银杏枝种群构件生长影响研究[J].西南农业大学学报,2003,25(6):475-478.
[11]Lehtila K,Tuomi J,Sulkinoja M.Bud demography of mountain birch Betula pubescens spp tortuosa near tree line[J].Ecology,1994,75(4):945-955.
[12]董鸣,张淑敏,陈玉福.匍匐茎草本蛇莓对基质养分条件的克隆可塑性[J].植物学报,2000,42(5):518-522.
[13]刘金平,张新全,游明鸿.野生扁穗牛鞭草无性系种群构件组成及生物量结构多样性[J].生态学报,2006,26(11):3656-3660.
[14]盘朝邦.老芒麦品系几种经济性状的遗传参数分析[J].中国草地,1989,(3):47-50.
[15]毛培胜,韩建国,王颖,等.施肥处理对老芒麦种子质量和产量的影响[J].草业科学,2001,(4):7-13.
[16]徐智明,周青平,刘云芬,等.平衡施肥对老龄多叶老芒麦种子产量的影响[J].甘肃农业大学学报,2004,(6):639-643.
[17]刘文辉,周青平,颜红波,等.高寒地区施肥对短芒老芒麦草产量和种子产量的影响[J].草地学报,2007,(2):193-195.
[18]贺晓,李青丰.4种微量元素对老芒麦种子质量的影响[J].草业学报,2007,16(3):88-92.
[19]张锦华,李青丰,李显利.氮、磷肥对旱作老芒麦种子生产性能作用的研究[J].中国草地,2001,(2):38-41.
[20]毛培胜,韩建国,吴喜才.收获时间对老芒麦种子产量的影响[J].草地学报,2003,11(1):33-37.
猜你喜欢
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14
新农民(2021年15期)2021-09-16
草业科学(2021年5期)2021-06-18
耕作与栽培(2021年2期)2021-05-19
中国畜禽种业(2019年7期)2019-01-06
西藏农业科技(2018年4期)2018-04-25
南风(2017年34期)2018-01-16
科学种养(2017年6期)2017-06-13
科学种养(2017年5期)2017-05-17
中国草食动物科学(2016年1期)2016-01-27
