
黄河口滨岸潮滩不同生境下翅碱蓬生物量空间分形特征与磷营养动态
2012-01-02 02:48牟晓杰孙志高刘兴土
草业学报 2012年3期
牟晓杰,孙志高,刘兴土
(1.中国科学院东北地理与农业生态研究所,吉林 长春130012;2.中国科学院海岸带环境过程重点实验室 山东省海岸带环境过程重点实验室 中国科学院烟台海岸带研究所,山东 烟台264003;3.中国科学院研究生院,北京100049)
*生物量空间分形特征是种群数量结构研究的基础和重要组成部分,对其进行研究不仅可深入了解种群的生物生态学特性,而且还可进一步揭示第一性生产力的形成过程。湿地营养元素循环是现代湿地生态学研究的热点[1,2]之一,磷(P)是湿地植物光合作用和初级生产过程中非常重要的营养元素,对其累积与分配特征进行研究是循环研究的重要基础,对深入了解湿地生态系统的生态过程和生态功能极为重要[3,4]。目前,国外学者已在湿地植物 P累积特征[5,6]、器官分布[5-8]、P养分利用率[6-9]以及 N、P交互作用的影响[8]等方面开展了大量研究。国内主要在湿地植物 P累积特征[10-12]、器官分布[10-14]、养分利用[14-16]以及 P 生物循环特征[17,18]等方面取得了较多成果。尽管国内外已对湿地植被P的累积与分配特征进行了较多研究,但这些研究大多针对同一生境条件下某种植被或不同生境条件下多种植被P累积与分配特征的相关研究,而关于不同生境条件下同种湿地植被P累积与分配特征的对比研究还不多见。
黄河三角洲由黄河携带大量泥沙填充渤海淤积而成,是中国暖温带保存最完整、最广阔和最年轻的湿地生态系统,也是东北亚内陆和环西太平洋鸟类迁徙的重要中转站、越冬栖息地和繁殖地。翅碱蓬(Suaedasalsa)是黄河三角洲重要的盐生植被之一,而作为该区湿地系统演替先锋阶段的翅碱蓬盐沼,是黄河三角洲丹顶鹤、白鹭、鹆形目鸟类等的主要栖息地,同时还起着维持湿地系统正常演替,防风固堤,调节气候等多种重要功能。翅碱蓬常分布于海陆交互作用的潮滩地带,大面积翅碱蓬在潮滩分布形成奇特壮丽的“红地毯”景观。潮汐作用是影响潮滩湿地变化的重要驱动力,在海陆相互作用下,潮滩地带的各种环境因素变化剧烈、频繁,湿地的稳定性、干扰程度及频率也不断变换,湿地面貌和结构始终处于变化之中。在翅碱蓬生长区内,由于潮滩不同位置环境条件特别是水盐条件的差异,分别在中潮滩和低潮滩形成了2种不同表现型的翅碱蓬群落,其对盐分变化极为敏感[19]。目前,国内关于不同水盐梯度带上不同翅碱蓬群落生物量空间分形特征与P营养动态的对比研究还未见报道。为此,本研究以位于中潮滩和低潮滩上的2种表现型翅碱蓬群落为对象,探讨其生物量空间分形特征、P累积与分配特征、N和P养分限制状况及其对盐分指示敏感程度的差异,以为2种翅碱蓬湿地营养物质循环和能量流动等研究提供基础数据。
1 材料与方法
1.1 研究区概况
试验于2008年5-11月在山东省黄河三角洲国家级自然保护区(东营)现黄河入海口滨岸潮滩进行。黄河三角洲国家级自然保护区(37°40′~38°10′N,118°41′~119°16′E)于1992年经国家林业局批准建立,是全国最大的河口三角洲自然保护区,在世界范围河口湿地生态系统中具有较强的代表性,主要保护黄河口新生湿地生态系统和珍稀濒危鸟类。保护区总面积15.3万hm2,其中核心区面积5.8万hm2,缓冲区面积1.3万hm2,试验区面积8.2万hm2。保护区属暖温带季风气候区,具有明显的大陆性季风气候特点,雨热同期,四季分明,冷热干湿界限极为明显。春季干旱多风,常有春旱,夏季炎热多雨,高温高湿,时有台风侵袭。该区年平均气温12.1℃,无霜期196d,≥10℃的年积温约4 300℃,年均蒸发量1 962mm,年均降水量为551.6mm,70%的降水集中于7和8月。保护区的土壤类型主要为隐域性潮土和盐土,主要植被类型为芦苇(Phragmitesaustralis)群落、芦苇-荻(Triarrhenasacchariflora)群落、穗状狐尾藻(Myriophyllumspicatum)群落、柽柳(Tamarixchinensis)群落、翅碱蓬群落及补血草(Limoniumsinense)群落等,其中芦苇群落、柽柳群落及翅碱蓬群落分布较广。
1.2 研究方法
1.2.1 植物样品采集与测定 翅碱蓬种子一般于4月末萌发,7月初开花,10月末成熟,11月末死亡。本试验于2008年5-11月,采用定位研究方法,按照“典型性、代表性、一致性”的原则,在现黄河入海口滨岸潮滩翅碱蓬分布区内,选择中潮滩翅碱蓬群落(37°45′57.0″N,119°09′40.7″E,JP1)和低潮滩翅碱蓬群落(37°46′38.9″N,119°09′41.4″E,JP2)为研究对象,分别设置30m×30m的样地,测定不同表现型翅碱蓬的地上、地下生物量。地上生物量采用收获法,即在典型样地按植物生长特点每20d左右采样1次。采样时,随机选取4~5个50cm×50cm的样方,用剪刀沿地面剪下植物地上部分,带回实验室,并将其分离为茎、叶。由于成熟期翅碱蓬的果实与小叶紧密相连,难以准确区分,所以实际为叶+果实。地下生物量采用挖掘法,并在地上生物量测定小区内进行测定。采样时,将样方内0~40cm的根全部挖出,放在细纱网袋中将泥土冲洗干净至无。生物量测定的同时,随机在每个样地内设置2~3个50cm×50cm的样方,用剪刀沿地面将植物的地上部分逐株剪下,带回实验室,在分别测定植物单株株高后,将其装入信封中。将采集的所有植物样品置于烘箱中先于105℃下杀青,后于80℃下烘干至恒重。称量后,计算2种翅碱蓬的地上、地下生物量,并将每个测定时期所得到的单株株高与株重数据建立数据库。同时,将植物样品粉碎,过0.25mm筛后备用。全磷(TP)含量测定采用H2SO4-H2O2消煮,钼锑抗比色法,全氮(TN)含量测定采用半微量凯氏法[20]。
1.2.2 环境因子测定 试验进行的同时,采用常规方法定期测定生长季内不同翅碱蓬群落的环境因子,如不同深度地温、电导率、pH和土壤含水量等。另外,采集不同层次土壤样品,将采集的土样及时带回实验室自然风干后,捡去残根等杂物,用球磨机磨碎,过0.149mm筛后测定其有机质、全氮、铵态氮和硝态氮含量。其中,全氮含量采用凯氏法测定,有机质含量采用重铬酸钾容量法测定[20]。铵态氮和硝态氮含量在用2mol/L的KCl浸提后通过连续流动分析仪(Bran-Lubee AAA3)测定。
1.2.3 计算方法 1)空间分形维数[21]:

式中,B为单株株重,H为单株株高,D为斜率,C为常数。计算时,分别从每次测定所建立的数据库中随机选取45~55株翅碱蓬个体地上生物量与株高数据,在双对数坐标下对B和H的一系列对数值进行线性回归,所得拟合直线斜率即为翅碱蓬地上生物量空间结构静态分形维数。再分别从每次测定的数据库中随机选取15株植物个体地上生物量与株高数据,计算整个生长期翅碱蓬种群地上生物量空间结构动态分形关系的D值。
2)植物P累积速率(VP,g/m2·d)[12]:
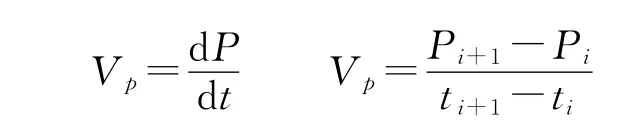
式中,Pi、Pi+1分别为ti、ti+1时刻的P累积量(g/m2)。
1.3 数据处理
运用Origin 8.0软件进行作图、计算和数学模拟,运用SPSS 16.0软件进行统计分析。
2 结果与分析
2.1 生物量空间分形特征
2.1.1 空间结构静态分形 JP1、JP2的lnB和lnH均具有良好线性关系(表1),二者相关系数分别为0.546~0.811和0.612~0.865,且均达到1‰的极显著水平。相应的,不同测定时期的D值分别为1.692~3.839和2.260~3.955,总体呈先增加后降低趋势。从不同时期地上生物量与株高的良好关系可知,二者植物体均具有自相似性,其空间结构具有明显分形特征,不同时期的地上生物量均是以株高的幂函数形式积累。一般而言,D值越大,表明其生物量空间积累越多,占据空间越大,反之则表明生物量空间积累越少,占据空间也越小。对比研究发现,除7和9月中旬JP2的D值低于JP1外,其他时期均明显高于JP1(表1),说明JP2单株生物量的空间积累及空间占据能力整体大于JP1。方差分析表明,二者D值在生长季内尽管差异较大,但并未达到显著水平(P>0.05)。
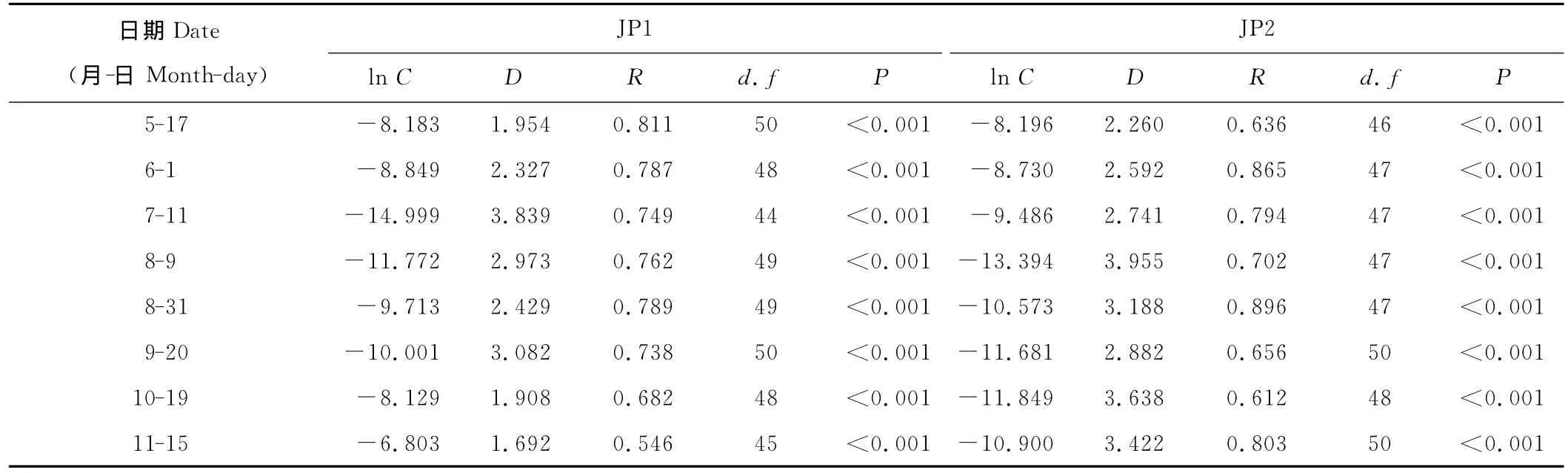
表1 地上生物量空间结构分形模型参数Table 1 Fractal model parameters of spatial structure of aboveground biomass
2.1.2 空间结构动态分形 JP1、JP2地上生物量与株高的对数值均存在显著线性关系(P<0.01)(图1),相关系数分别为0.834和0.876,D值分别为2.012和2.366,表明生长季内JP1、JP2地上生物量增长与株高的关系具有自相似性,分别遵从D=2.012和D=2.366的幂函数规律增长。另外,二者动态条件下地上生物量与株高自相似特征的存在还表明,较高植株是较矮植株地上生物量积累的放大形式。从统计意义上讲,二者的较大植株均可以看作是由其较小植株经过生长过程放大得到,而表征其放大过程的度量值即为D值,反映出二者均是一种分形生长过程。

图1 植物株重与株高的动态分形关系Fig.1 Dynamics of fractal relationships between plant biomass(B)and height(H)
2.2 磷营养结构动态
JP1、JP2不同器官因生长阶段和自身组织结构的不同,其TP含量均具有明显季节变化(图2)。二者根、茎的TP含量变化模式较为一致,其在生长初期均较高,之后整体呈下降趋势,并于11月中旬达到最低值。与之相比,二者叶的TP含量波动明显且变化模式差异较大。具体来说,JP1叶的TP含量在8月末之前呈较缓波动变化,9-11月则呈倒“V”型变化。而JP2叶的TP含量在生长阶段整体呈“V”型变化,并分别于7月中旬和11月中旬达到最低值和最高值。二者叶中TP含量在8月末前的变化可能主要与此间叶生物量增加产生的“稀释效应”差异有关,之后则与成熟期果实发育期间根、茎中的P营养向叶和果实的转移机制与转移程度有关,而这也是导致茎和根中TP含量降低的一个重要原因。就P分布而言,二者不同器官的TP含量在大多时期均表现为叶>茎>根,说明叶是P的主要累积器官。对比研究发现,JP1不同器官(根、茎、叶)的P含量一般均明显高于JP2的相应器官,前者分别为后者的(1.98±0.64),(1.41±0.46),(1.98±0.93)倍(n=8),说明JP2生长所受P养分的限制程度可能大于JP1。经方差分析,二者根中TP含量变化差异显著(P<0.05),而茎和叶中TP含量变化的差异不显著(P>0.05)。

图2 不同器官全磷含量变化Fig.2 Changes of total phosphorus content in different organs
2.3 磷营养累积动态
2.3.1 磷累积量动态 生长季内JP1、JP2地上器官的P累积量分别为0.062~1.671和0.035~0.201g/m2,地下部分分别为0.023~0.145和0.002~0.012g/m2(表2)。除11月中旬外,JP1地上器官(茎、叶)的P累积量均明显高于JP2,分别为JP2的1.87~15.18和1.61~9.10倍,而根在生长季内均表现为JP1>JP2,为JP2的3.32~20.07倍。以上结果说明,JP2不同器官较低的P累积量可能主要与其生长所受P养分限制状况有关。具体而言,JP1、JP2地上器官的P累积量自5月中旬开始逐渐增加,并于生长旺期或成熟期达到最大值,之后随地上器官枯萎而不断降低。与之相比,JP1根的P累积量整体呈“增加-降低”变化,而JP2则呈明显波动变化。比较而言,JP1不同器官P累积量在成熟期以前表现为茎>叶>根,之后为叶>茎>根,而JP2不同器官P累积量生长季整体表现为叶>茎>根。比较而言,JP1、JP2各器官P储量在不同时期所占比例差异较大,但整体以叶所占比例为优,茎次之,而根最低(表3)。另外,二者植物体的P累积总量也差异较大,除11月中旬外,JP1植物P累积总量均明显高于JP2,为JP2的1.95~11.90倍。
2.3.2 磷累积速率动态 JP1地上器官的VP整体呈“增加-降低”变化(表2),除6月初和11月中旬为负值外,其他时期均为正值。7-8月,由于植物处于生长旺期,对P的需求量很大,而叶又是P的重要储库,由此导致此间叶的VP为正值且相对较高。9-10月,植物处于果实成熟期,P在果实中大量累积,由此导致此间叶+果实的VP出现较大峰值。11月之后,植物不断枯萎,此间的P也开始不断转移,由此导致叶+果实的VP呈较大负值。JP1茎的VP在8月上旬达到最大正值,而于10月中旬取得最大负值,且其在此间的变化与叶大致呈相反变化,反映了叶与茎在P养分累积方面的密切联系。与之相比,JP2地上及不同器官VP的波动变化更为明显,其中叶+果实的VP除7月中旬和11月中旬为负值外,其他时期均为正值,原因可能主要与JP1、JP2的植物生态学特性及所处水盐状况的差异有关。茎的VP分别在6月初和8月末出现2次峰值,而于9月中旬达到最大负值,原因可能与8月末之后茎逐渐衰老,其中的P开始向叶+果实的转移机制和转移程度有关。比较而言,JP1地上部分VP的变化在8月末前与茎较为相似,之后则与叶较为一致,而JP2地上部分VP的变化在生长季内整体均与叶+果实基本一致。与地上及不同器官的VP变化相比,二者地下部分VP的变化一直较为平缓。

表2 不同器官P累积量(A)与累积速率(VP)Table 2 Changes of phosphorus accumulation amount(A)and accumulation rate(VP)in different organs

表3 不同器官P储量比例Table 3 Proportions of phosphorus storage in different organs %
2.4 湿地植物N/P动态
N、P是生态系统中非常重要的元素,其供给状况对于植物的初级生产具有重要影响。N/P可作为氮饱和的诊断指标,对植物生长限制元素有着重要指示意义[22,23]。已有研究表明[24],当湿地植物N/P<14时,植物生长受N限制;N/P为14~16,则同时受N、P限制;N/P>16,则受P限制。利用该关系来探讨黄河口滨岸潮滩湿地植物的N、P养分限制状况可知,JP1植物的N/P为9.87±3.47<14(表4),说明其生长受N限制,这与章文龙等[15]对闽江河口湿地植物、肖蓉等[25]对二百方子湿地植物、Pezeshki和Delaune[26]对盐滩植物限制元素的结论一致。与之相比,JP2的N/P为14<15.73±5.00<16(表4),说明其生长同时受N、P限制,但更多受P限制。这一结果正好与前述P是JP2植物生长重要限制元素的推测相一致。另外,通过对不同时期、不同器官N/P的研究发现,JP1、JP2不同器官N/P高低及其变化具有明显的时间性,而这种时间性又主要与不同时期各器官对N、P的吸收利用状况以及土壤中有效N、P养分的供给状况有关。

表4 不同器官的N/P动态Table 4 Dynamics of N/P ratio of different organs
3 讨论
3.1 生物量空间结构差异
生物量空间结构是反映湿地植物群落特征的重要参数。本研究表明,黄河口滨岸潮滩JP1、JP2的生物量均具有明显空间结构分形特征,其对空间的占据能力总体表现为JP2>JP1。水盐梯度是黄河口滨岸潮滩湿地最具典型的环境因子,是决定植被分布与演替的关键因素[27]。已有研究表明,水分、盐分条件可直接影响植物生物量空间分配的变化。王丽等[28]的研究表明,水分梯度及水分梯度下土壤环境条件是影响三江平原典型小叶章(Calamagrostisangustifolia)生物量分异的主要原因。贺海波和李彦[29]探讨了盐分胁迫对猪毛菜(Salsolanitraria)、盐生草(Halogetonglomeratus)2种盐生植物生长和生物量分配的影响。结果发现,盐分胁迫显著影响了2种盐生植物的生长和生物量空间分配。Rawat和Banerjee[30]的研究也发现,盐分对印度黄檀(Dalbergiasissoo)幼苗的生物量空间分配具有重要影响。本研究中,JP1、JP2主要受盐分胁迫和水分梯度2种环境因子的影响。相对于JP1,JP2由于经常受到海流、潮汐等海洋作用力的影响,不仅面临盐分胁迫,而且还同时受到海水渍涝等带来的综合逆境胁迫[31]。假定JP2与JP1一样具有较高的株高,则生态系统很易受到逆境胁迫的破坏,进而使得系统将处于不稳定状态。在这种环境下,JP2可能形成了一种区别于JP1的特殊适应对策,即通过降低株高、扩大植株茎和叶空间占据能力来适应综合逆境胁迫。另外,由于N、P是生态系统中非常重要的元素,其供给状况对植物的初级生产具有重要影响,所以JP1、JP2不同器官生物量空间分布及其差异亦受到其所处生境养分状况、地形和地貌等因素不同程度的影响。本研究表明,N是JP1湿地植物净初级生产力的限制养分,而N、P(主要是P)同时为JP2植物初级生产力的限制养分。养分元素的受限及其限制程度必然影响植物生物量分配[15]。由于N是蛋白质的主要成分,而蛋白质是植物细胞的主要组成部分(约占50%以上)。P在植物体内主要与其他有机物结合形成磷脂、核酸和辅酶等,其植物生理意义与N差异较大[15]。而可能正是由于N、P在植物生理意义上的差异以及JP1、JP2湿地养分限制状况的差异使得二者的生物量空间分布在一定程度上产生较大分异。
3.2 植物P含量、累积量及分配比的变化及差异
本研究表明,黄河口滨岸潮滩2种表现型翅碱蓬不同器官的P含量、累积量、累积速率及分配比均具有明显季节变化,且其之间均存在不同程度差异。原因可能主要有两方面:1)与2种表现型翅碱蓬自身生态学特性的差异有关。由于JP1、JP2分别处在不同的水盐梯度带上,其所处生境的差异导致翅碱蓬群落形成了绿色和紫红色2种具有不同特性的表现型,而相关研究发现,二者叶片肉质化程度[32]、叶片色素积累及光合特性[31]、叶片抗氧化系统[33-35]等均明显不同,内部组织结构的差异必然影响植物对营养元素的吸收状况,进而导致二者P累积与分配特征的差异。2)与二者所处环境条件的差异有关。生长季内黄河口滨岸低潮滩与中潮滩湿地的土壤理化性质、土壤温度、水分条件及养分条件等均存在明显差异[19],而这些环境因素的差异均会对2种表现型翅碱蓬P的吸收、利用状况及其差异产生重要影响。一些相关研究也表明,湿地环境条件(如养分状况)对植物P的累积与分配具有重要影响。Xie等[14]的研究表明,环境养分供给状况(不同N、P养分处理)可明显影响P在水葫芦不同器官中的累积与分配率,且当养分含量增加后,植物体自身会适时调整生物量分配以达到对获得资源的最佳利用。Romero等[8]的研究则表明,环境养分供给状况虽然对芦苇的生长和N、P分配具有一定影响,但P对其产生的影响并不显著。黄河口滨岸潮滩湿地土壤的养分含量非常低[19],说明2种表现型翅碱蓬对贫养分条件均具有较强的适应能力。另外,黄河口滨岸低潮滩湿地相对于中潮滩湿地经常受到海流、潮汐等海洋作用力的影响,盐度相对较高,盐碱化现象较为严重。而相关研究表明,盐分胁迫会对植物P的累积分配特征产生重要影响,Mashhday等[36]研究了水分和盐分胁迫对小麦(Triticumaestivum)和黑小麦(Triticalerimpau)N、P和氯化物累积特征的影响,结果发现,盐分和水分显著影响了植物的生长以及对N、P的吸收,随盐分增加,植物养分累积量逐渐减少,且高的水分条件可以增加植物对盐分的耐受限度。侯振安等[37]的研究则表明,盐分胁迫对羊草(Leymuschinensis)N、P、K的吸收具有很强的抑制作用。崔保山等[38]的研究还发现,翅碱蓬的盐分生态阈值区间为5.17~20.25g/kg,盐分过高或过低都不利于翅碱蓬的生长及养分吸收。可见,盐分条件是导致2种表现型翅碱蓬P累积与分配特征差异的另一个主要原因。同时,潮滩湿地的热量条件、水分条件等大多直接影响湿地植物的生长节律和生态学特性而影响其对P的累积与分配。总之,黄河口滨岸潮滩湿地是一个复杂的生态系统,2种表现型翅碱蓬对P的吸收、累积与分配特征的变化及差异在很大程度上取决于土壤、水体、潮汐、气候和植物自身生理等因素的综合影响。
[1]Mistch W J,Gosselin J G.Wetlands[M].New York:Van Nostrand Reinhold Company Inc.,2000:89-125.
[2]章家恩,徐琪.现代生态学研究的几大热点问题透视[J].地理科学进展,1997,16(3):29-37.
[3]Woodmansee R G,Duncan D A.Nitrogen and phosphorus dynamics and budgets in annual grasslands[J].Ecology,1980,6:893-904.
[4]郭彦军,倪郁,韩建国.农牧交错带人工种草对土壤磷素有效性的影响[J].草业学报,2010,19(2):169-174.
[5]Greenway M,Woolley A.Constructed wetlands in Queensland:Performance efficiency and nutrient bioaccumulation[J].Ecological Engineering,1999,12:39-55.
[6]Davis S M.Growth,decomposition,and nutrient retention ofCladiumjamaicenseCrantz andTyphadomingensisPers.in the Florida Everglades[J].Aquatic Botany,1991,40(3):203-224.
[7]Garver E G,Dubbe D R,Pratt D C.Seasonal patterns in accumulation and partitioning of biomass and macronutrients inTyphaspp.[J].Aquatic Botany,1988,32(1-2):115-127.
[8]Romero J A,Brix H,Comín F A.Interactive effects of N and P on growth,nutrient allocation and NH4uptake kinetics byPhragmitesaustralis[J].Aquatic Botany,1999,64:369-380.
[9]Lorenzen B,Brix H,Mendelssohn I A.Growth,biomass allocation and nutrient use efficiency inCladiumjamaicenseandTyphadomingensisas affected by phosphorus and oxygen availability[J].Aquatic Botany,2001,70:117-133.
[10]郭长城,胡洪营,李锋民,等.湿地植物香蒲体内氮、磷含量的季节变化及适宜收割期[J].生态环境学报,2009,18(3):1020-1025.
[11]刘长娥,杨永兴,杨杨.九段沙中沙湿地植物磷元素的分布累积与动态[J].同济大学学报(自然科学版),2008,36(11):1537-1541.
[12]秦胜金,刘景双,孙志高.三江平原湿地小叶章群落磷素积累动态与生物量动态分析[J].生态学杂志,2006,25(6):646-651.
[13]孙志高,刘景双,王金达,等.三江平原不同群落小叶章种群生物量及氮、磷营养结构动态[J].应用生态学报,2006,17(2):221-228.
[14]Xie Y H,Wen M Z,Yu D.Growth and resource allocation of water hyacinth as affected by gradually increasing nutrient concentrations[J].Aquatic Botany,2004,79:257-266.
[15]章文龙,曾从盛,张林海,等.闽江河口湿地植物氮、磷吸收效率的季节变化[J].应用生态学报,2009,20(6):1317-1322.
[16]徐德福,李映雪,赵晓莉,等.重金属锌胁迫下湿地植物对磷的吸收能力变化研究[J].环境工程学报,2009,3(6):1033-1036.
[17]刘长娥,杨永兴.九段沙芦苇湿地生态系统 N、P、K的循环特征[J].生态学杂志,2008,27(3):418-424.
[18]向万胜,童成立,吴金水,等.湿地农田土壤磷素的分布、形态与有效性及磷素循环[J].生态学报,2001,21(12):2067-2073.
[19]牟晓杰,孙志高,王玲玲,等.黄河口滨岸潮滩不同表现型翅碱蓬 K、Ca、Mg含量与累积的季节变化[J].草业学报,2010,19(3):177-190.
[20]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999.
[21]马克明,祖元刚.羊草种群地上部生物量与株高的分形关系[J].应用生态学报,1997,8(4):417-420.
[22]Gusewell S,Koerselman W,Verhoeven J T A.Biomass N-P ratios as indicators of nutrient limitation for plant populations in wetlands[J].Ecological Applications,2003,13:372-384.
[23]Tessier J T,Raynal D J.Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation[J].Journal of Applied Ecology,2003,40:523-534.
[24]Koerselman W,Meuleman A F M.Vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation[J].Journal of Applied Ecology,1996,33:1441-1450.
[25]肖蓉,白军红,高海峰,等.封闭性和开放性沼泽湿地土壤全磷的季节变化特征[J].草业学报,2010,19(3):88-93.
[26]Pezeshki S R,Delaune R D.Carbon assimilation in contrasting streamside and inlandSpartinaalterniflorasalt marsh[J].Vegetation,1988,76:55-61.
[27]王海梅,李政海,宋国宝,等.黄河三角洲植被分布、土地利用类型与土壤理化性状关系的初步研究[J].内蒙古大学学报(自然科学版),2006,37(1):69-75.
[28]王丽,胡金明,宋长春,等.水分梯度对三江平原典型湿地植物小叶章地上生物量的影响[J].草业学报,2008,17(4):19-25.
[29]贺海波,李彦.干旱、盐胁迫条件下两种盐生植物生物量分配对策的研究[J].干旱区研究,2008,25(2):242-247.
[30]Rawat J S,Banerjee S P.The influence of salinity on growth,biomass production and photosynthesis ofEucalyptuscamaldulensisDehnh.andDalbergiasissooRoxb.seedlings[J].Plant and Soil,1998,205:163-169.
[31]阮圆,刘彧,王宝山.不同自然盐渍生境下盐地碱蓬叶片色素积累及光合特性的研究[J].山东师范大学学报(自然科学版),2008,23(1):115-117.
[32]刘彧,丁同楼,王宝山.不同自然盐渍生境下盐地碱蓬叶片肉质化研究[J].山东师范大学学报(自然科学版),2006,21(2):102-104.
[33]王琳,岳晓翔,王宝山.不同生境下两种表型盐地碱蓬叶片POD比较研究[J].山东师范大学学报(自然科学版),2008,23(4):103-105.
[34]岳晓翔,陈敏,段迪,等.绿色和紫红色表型盐地碱蓬叶片抗氧化系统比较研究[J].山东师范大学学报(自然科学版),2008,23(1):121-124.
[35]闫留华,彭建云,陈敏,等.潮间带生境下两种表型盐地碱蓬的抗氧化系统比较[J].植物生理学通讯,2008,44(1):109-111.
[36]Mashhday A S,Sayed H I,Heakal M S.Effect of soil salinity and water stresses on growth and content of nitrogen,chloride and phosphate of wheat and triticale[J].Plant and Soil,1982,68:207-216.
[37]侯振安,李品芳,龚元石.盐渍土壤环境对羊草生长与养分吸收的效应[J].石河子大学学报(自然科学版),2001,5(1):6-10.
[38]崔保山,贺强,赵欣胜.水盐环境梯度下翅碱蓬 (Suaedasalsa)的生态阈值[J].生态学报,2008,28(4):1408-1418.
猜你喜欢
海洋通报(2022年5期)2022-11-30
河海大学学报(哲学社会科学版)(2022年4期)2022-11-21
流行色(2021年8期)2021-11-09
海洋通报(2021年1期)2021-07-23
动漫星空(兴趣百科)(2020年11期)2020-11-09
海洋通报(2020年3期)2020-10-20
汉语世界(The World of Chinese)(2019年5期)2019-11-11
趣味(数学)(2019年12期)2019-04-13
现代装饰(2018年11期)2018-11-22
