
华山松种子大小对啮齿动物贮藏行为的影响及其时空变化
2011-12-25 01:02陈帆,陈进
Zoological Research 2011年4期
陈 帆, 陈 进
(1. 中国科学院西双版纳热带植物园 热带森林生态学重点实验室, 云南 勐腊 666303; 2. 中国科学院研究生院, 北京 100049)
华山松种子大小对啮齿动物贮藏行为的影响及其时空变化
陈 帆1,2, 陈 进1,*
(1. 中国科学院西双版纳热带植物园 热带森林生态学重点实验室, 云南 勐腊 666303; 2. 中国科学院研究生院, 北京 100049)
华山松(Pinus armandii)是广泛分布在中国中西部海拔1 000~3 300 m的山地、种子较大(约300 mg)的松属植物。为探究华山松种子大小对啮齿动物贮藏行为的影响, 于2006年和2007年在滇西北3个不同地点进行种子标记和追踪实验。结果表明, 在所有年份和地点, 啮齿动物都倾向于贮藏更多的大种子和取食更多的小种子;啮齿动物贮藏大种子的数量, 以及平均距离和最大距离均显著高于小种子。3个地点具有不同的啮齿动物群落结构, 从而对种子命运产生显著影响。种子命运在两个年份间也存在显著差异。
Pinus armandi; 啮齿动物; 种子散布; 种子命运; 分散贮藏; 贮藏距离
啮齿动物对大种子和坚果类植物的散布和命运具有直接影响(Forget & Milleron, 1991; Forget, 1992; Theimer, 2001; Xiao & Zhang, 2004)。啮齿动物会采食相当比例的种子, 但同时也会散布种子,从而提高种子的分散度, 而分散贮藏可以降低种子被其他捕食者取食的几率, 从而提高幼苗建成的几率(Vander Wall, 1992, 2003), 啮齿动物也可能会将种子移动到更适合萌发和幼苗建成的微环境中(Wenny, 2000)。
同一种植物的种子大小存在显著差异的现象在植物的野生居群中很普遍, 并且对植物个体的繁殖成功很重要(Moegenburg, 1996; Brewer, 2001)。这种差异既对植物居群, 也对传播种子的动物产生影响。植物常常能从生产大种子中得到益处, 比如大种子建立幼苗的存活率通常高于小种子(Moegenburg, 1996; Khan, 2004; Moles & Westoby, 2004)。但是, 由于植物用来生产种子的资源是有限的, 相对于生产少量的大种子, 植物往往生产出数量更多的小种子(Smith & Fretwell, 1974), 因为对于许多植物来说, 小种子被散布的几率更高(Levey, 1987), 能被更多类型的种子散布者散布(Moermond & Denslow, 1985), 或被散布更远的距离(Pizo &Oliveira, 2001)。对于被啮齿动物散布的植物来说,情况却并不完全如此(Jansen et al, 2004)。啮齿动物往往更分散地贮藏大种子以保护其免遭其它竞争者的取食, 大种子的生存几率反而更大(Vander Wall, 1995)。大种子还有更大可能被啮齿动物重新挖掘出来, 被再次移动和贮藏到更远的地点(Vander Wall, 2003; Xiao et al, 2005), 由于大种子被更早地发现和移动更远的距离, 埋藏就更加分散,它们有更多建成幼苗的机会(Vander Wall, 2003)。同时, 在不同年份和群落结构条件下, 种子的命运往往具有显著的差异, 这种差异与生境内种子的相对和绝对可利用性、种子的能量价值以及群落内取食种子的动物的种类和多度等因素都随着年份和群落结构的变化而变化有关(Smallwood & Peters, 1986; Chambers & MacMahon, 1994)。
松属植物的种子质量从 3.5到 909 mg不等(Benkman, 1995)。其中大约80种产生有翅的小种子, 可以利用风进行散布。另外, 30种具有无翅或近乎无翅的质量大于90 mg的大种子, 往往由鸦科动物(Tomback & Linhart, 1990)或者分散贮藏食物的啮齿动物(Vander Wall, 2003)散布。在某些相对隔绝的地区, 啮齿动物甚至是惟一种子散布者(Tomback et al, 2005)。华山松(Pinus armandiiFranch.)种子无翅或近无翅, 平均质量在300 mg左右, 主要由脊椎动物采食和散布(个人观察)。我们在不同年份和地点选择不同大小的华山松种子进行释放和跟踪实验, 试图回答以下问题:(1)种子大小对啮齿动物移动, 取食和贮藏种子的行为有何影响; (2)种子大小对啮齿动物贮藏种子的次数和平均距离有何影响; (3)不同年份和地点啮齿动物的贮藏和取食行为有何差异。
1 研究地点和方法
1.1 研究地点
野外实验在云南省迪庆藏族自治州进行。华山松在迪庆州主要沿澜沧江和金沙江河谷两侧的横断山脉分布, 分布区的海拔高度范围在 2 000~3 500 m, 在其分布的上限和下限。华山松和云南松(P. yunnanensis)以及高山松(P. densata)形成混交林。华山松纯林主要呈带状或岛状间断分布, 梅里雪山、白马雪山、石卡雪山等横断山脉的山脊以及金沙江、澜沧江的干热河谷形成了华山松分布的自然隔离带。我们选择香格里拉县尼西乡汤满村(E 099°32′, N 28°00′, 2 830 m)、德钦县云岭乡明永村梅里雪山国家级自然保护区(E 098°47′, N 28°28′, 2540 m)、维西县塔城乡及其落林业工作站(E 099°16′, N 27°22′, 3 060 m)等3个地点作为实验地点。3个地点的上层乔木都以华山松为主, 混生有少量云南松或高山松, 高度为20~30 m, 盖度为50%~75%。下层灌木主要为黄背栎(Quercus pannosa)。
1.2 种子释放实验
使用人工释放实验和标签法(Xiao & Zhang, 2006)来研究不同地点在地面活动的啮齿动物对不同大小的种子的散布行为和选择倾向。实验所用的种子为当地农民当年从树上采集的新鲜种子。其中大种子的质量全部大于 500 mg, 平均为(561.4 ± 5.84) mg (Mean ±SE,N= 1 200), 小种子的质量全部小于 250 mg, 平均为(211.6 ± 3.50) mg (N= 1 200)。使用电钻和直径为0.5 mm的钻头在每一个种子上钻一个贯穿整个种子的小洞, 用一根细不锈钢丝穿过小洞并系住, 钢丝的另外一头系一个1 cm×2 cm的塑料片, 塑料片标记有编号, 每个种子都有一个特定的编号。野外及围栏的研究表明,塑料片对啮齿动物移动及埋藏种子无显著影响(Xiao et al, 2006)。在尼西、明永和维西各设有20个释放点, 释放点间隔距离在100 m以上。将10粒大种子和 10粒小种子呈环形释放在地面上, 种子向内, 标签向外, 释放点设置在华山松林冠的下方。每个地点共释放200粒大种子和200粒小种子,释放实验于2006年和2007年的10至12月在3个地点分别进行, 总共释放并追踪了2 400粒种子。
1.3 种子的命运和贮藏距离
在释放后的第1、3、5、7、9、11和13 d检查每个释放点, 统计还留在原地并且未被移动或取食的种子数量, 通过标签在释放点周围 30 m内寻找已经从释放点消失的种子。记录所有种子的编号和当时的状态。种子的状态分为留在原地和被移动 2种。被移动的种子又可分为4种:被移动并贮藏、被移动并取食(由于没有绝对的“原地”取食, 所有的取食都被视为被移动后的取食)、被移动但留在地面上未被取食和未能寻获。如果种子被移动后未被取食, 则用木棒标记种子所在的位置和编号, 在后面的调查中继续追踪这些种子的状态。我们无法确定种子的最终命运, 但在经过前7~9 d的频繁移动后, 种子被移动, 贮藏和取食的速率在 9~13 d时大大降低, 短期状态趋于稳定(个人观察), 所以,我们统计了每粒种子在13 d的实验结束后的状态。在实验过程中, 如果种子第一次被移动并被贮藏, 用皮尺测量贮藏点距离原始释放点的距离。如果第一次被贮藏的种子又第二次被贮藏, 测量二次贮藏点距离释放点的距离。
1.4 啮齿动物种类和数量的调查
用花生米作诱饵, 使用鼠笼对样地中啮齿动物的种类和数量进行取样调查。在每个样地中每隔10 m放置鼠笼2个(笼口分别朝向相反的方向), 共50个, 每天观测一次, 连续观测15 d, 共计750个笼日(一个鼠笼放置一昼夜的捕鼠单位)。记录捕获啮齿动物的种类和数量, 将其浸泡在70%乙醇溶液中带回实验室进行种类鉴定。在地面上堆砌陷阱, 用核桃仁作诱饵诱捕松鼠。由于松鼠比较敏感, 不易捕获, 同时走访当地猎人来了解样地中松鼠的种类并获取标本。
1.5 数据处理
使用 Wilcoxon检验比较各个地点和年份间大小种子命运的差异; 使用单因素分析(one-way ANOVA)比较大小种子贮藏距离之间的差异。所有数据都在SPSS for Windows 13.0软件上进行分析。
2 结 果
2.1 种子的移动速率
啮齿动物取食和移动种子的速率很快。移动主要集中在前7 d。7 d后, 移动速率大大降低(图1)。在尼西和维西, 大种子的移动速率略快于小种子,而在明永, 大种子和小种子的移动速率相差很小。从不同年份上看, 2007年种子被移动的速率快于2006年(P< 0.001, 图1)。2006年5 d内啮齿动物大约移动了40%的种子, 2007年则达到80%。

图1 大种子和小种子从释放点被移动的速率Fig. 1 Seeds removal rate at 3 study sites in 2006 and 2007
2.2 种子命运
在两年的研究中, 3个实验地点大小种子的命运有很大不同, 从整体来看, 大种子的贮藏比例高于小种子, 而取食比例低于小种子(图2)。具体来看, 2006年在尼西, 啮齿动物取食小种子的比例显著高于大种子(Z= -2.101,P= 0.036), 贮藏的比例则没有显著差别。在明永, 啮齿动物取食小种子的比例显著高于大种子(Z= -2.433,P= 0.015), 贮藏的比例则达不到显著水平(Z= -1.725,P= 0.084)。在维西, 啮齿动物贮藏大种子的比例显著高于小种子(Z= -2.844,P= 0.004), 取食比例没有显著差异。2007年的情况则有所不同, 除了在维西, 啮齿动物取食小种子的比例显著高于大种子(Z= -2.349,P= 0.019), 其它两个地点的取食比例, 以及所有地点的贮藏比例均无显著差异。
2.3 贮藏种子的距离
在所有年份和地点, 大种子被啮齿动物贮藏的平均距离和次数均大于小种子(表1)。大种子被贮藏的距离分布更广, 并且更为分散(图3)。2006年在尼西, 大种子被啮齿动物贮藏的距离显著高于小种子(F1,32= 4.57,P= 0.040), 而在明永和维西的差别不显著。在2007年, 维西的差异达到显著(F1,59= 4.31,P= 0.042), 而尼西和明永的差异不显著。
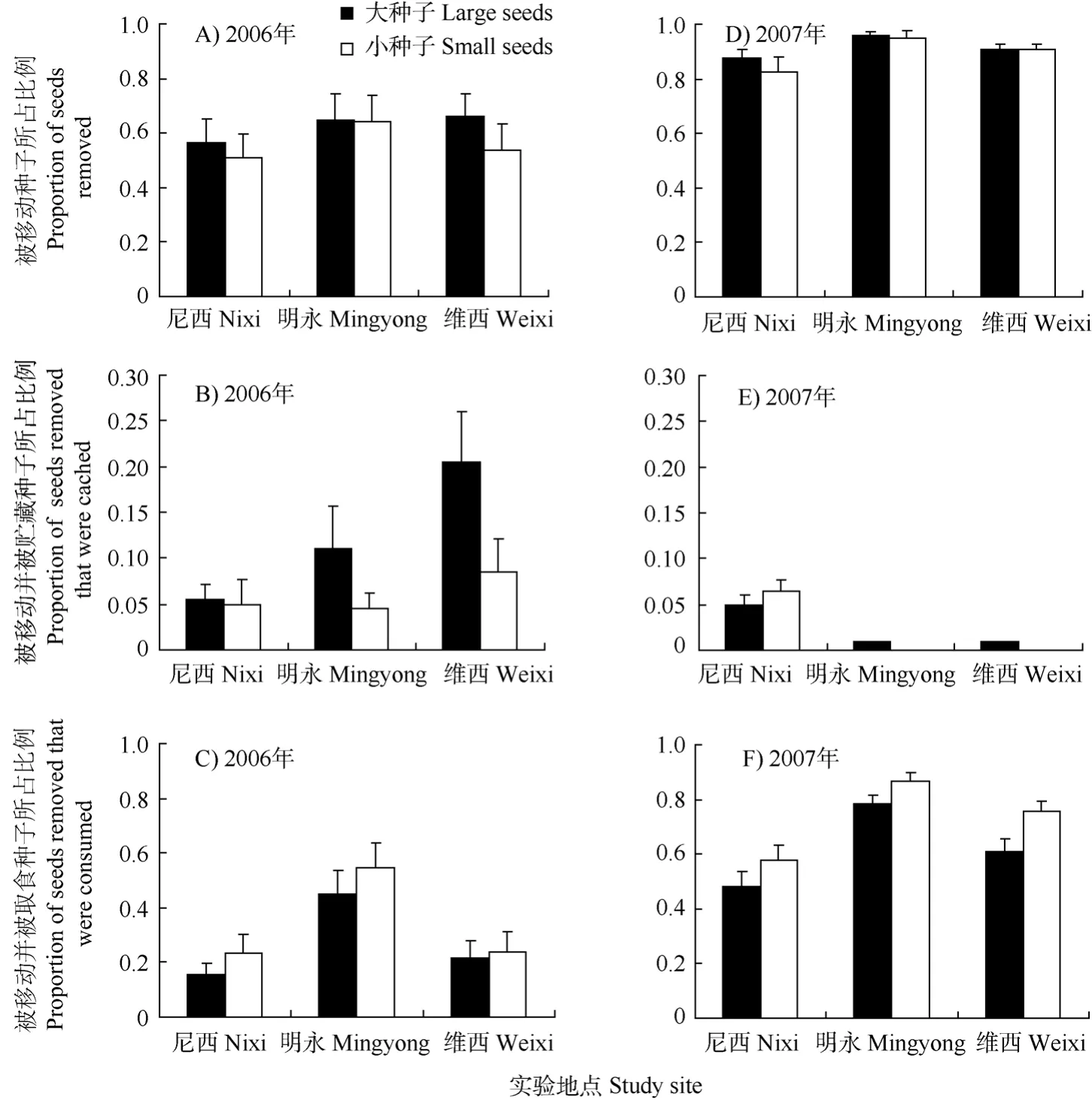
图2 被移动、被移动并被贮藏、被移动并被取食的大种子和小种子占所有释放的种子的比例(Mean ± SE, N = 20)Fig. 2 Proportion of seeds removed, removed and cached, removed and consumed at 3 study sites in 2006 and 2007 (Mean ± SE, N = 20)

表1 释放实验中第一次和第二次测量的啮齿动物贮藏种子的平均距离(m, Mean ± SE)和次数Tab. 1 Measurement of mean distance (m, Mean ± SE) and frequency of seed caching by rodents for the first and second time in seed-tracking experiment
2.4 啮齿动物的群落组成
两年中在3个地点一共捕获4种啮齿动物 (表2), 其中姬鼠属(Apodemus)的两个种在 3个点最为常见, 数量也最多。另外, 还发现有少量的西南绒鼠(Eothenomys custos)和社鼠(Niviventer confucianus)。维西的啮齿动物数量在两年中都是最高的。在尼西和维西于2007年的啮齿动物数量大大低于2006年,而在明永于 2007年的啮齿动物数量略有提高。实验过程中还发现3种采食种子的松鼠, 其中最常见的是珀氏长吻松鼠(Dremomys pernyi), 在尼西和明永都有分布, 而在尼西还发现有少量的赤腹松鼠(Callosciurus erythraeus), 在明永有一定数量的隐纹花松鼠(Tamiops swinhoei), 在维西没有发现松鼠活动。

图3 释放实验中大种子和小种子第一次被贮藏距离的分布Fig. 3 Distribution of seed caching distance for the first time in experiment at 3 study sites in 2006 and 2007
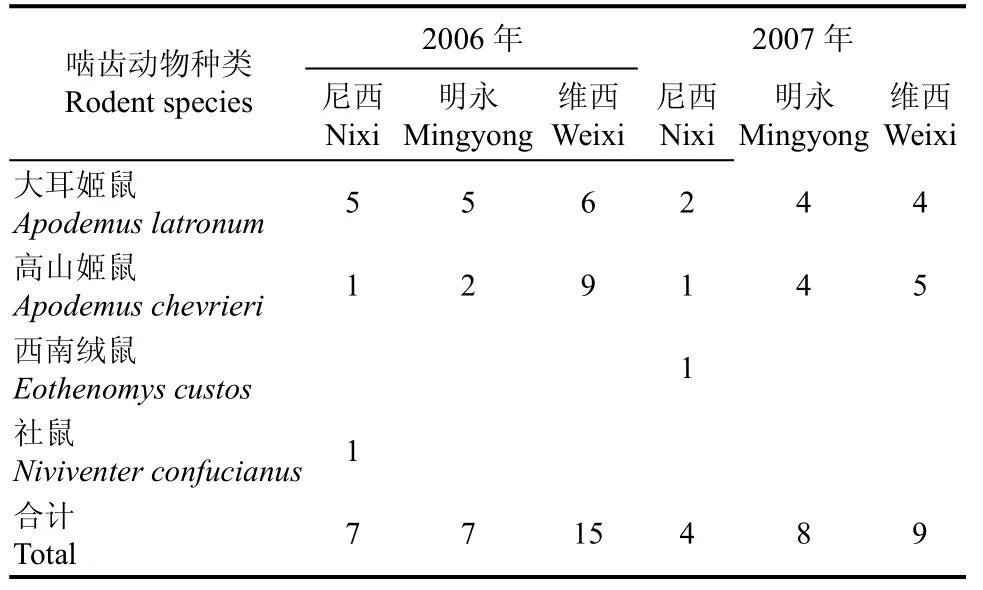
表2 3个实验地点捕获啮齿动物的种类和捕获率Tab. 2 Species and number of captured rodents at 3 study sites in 2006 and 2007
3 讨 论
3.1 种子大小对种子命运的影响
啮齿动物对种子贮藏的过程是在种子的隐藏度, 移动过程中能量消耗, 记忆和遗忘之间的权衡(Clarkson et al, 1986)。很多证据表明, 啮齿动物为了平衡移动, 以及贮藏过程中的能量消耗和获得(Stapanian & Smith, 1978; Clarkson et al, 1986), 更倾向于贮藏能量价值高的大种子和取食能量价值低的小种子(Vander Wall, 1995, 2003; Jansen et al, 2004; Zhang et al, 2008; Chang et al, 2009)。我们的研究也表明, 啮齿动物贮藏大种子的比例高于小种子, 而取食大种子的比例小于小种子(图 2)。惟一的例外是在 2007年的尼西, 啮齿动物贮藏了更多的小种子, 这是因为大种子虽然一次贮藏数量大于小种子(表1), 但更多的大种子又被取出并食用, 从而导致实验结束时, 大种子的贮藏比例反而小于小种子。
3.2 种子大小对贮藏距离的影响
在所有年份和地点的释放实验中, 大种子被贮藏的距离都大于小种子(表1)。这说明啮齿动物更倾向于将能量更丰富的大种子贮藏得更远, 从而增加了大种子散布的成功率。许多研究都表明了同一物种的大种子被移动的距离大于小种子被移动的距离(Brewer, 2001; Theimer, 2003; Jansen et al, 2004)。Xiao et al (2005)研究发现, 种子被贮藏的平均距离和最大距离会随着种子的增大而显著提高, 这和我们的研究结果类似(表1, 图3)。
3.3 不同地点啮齿动物群落结构对种子命运的影响
不同地点对啮齿动物的贮藏和取食行为也有影响。在两年中, 在明永的种子取食比例总是最高,而在尼西的取食比例总是最低(图 2)。这可能与 3个地点啮齿动物群落结构的组成有关。姬鼠是这个地区主要的分散贮藏种子的啮齿动物类群(Wang & Yang, 2007; Wang & Chen, 2009)。在维西, 姬鼠数量多, 活动频繁, 而很少有松鼠活动, 所以在 2006年在维西的分散贮藏作用最显著。在明永, 取食比例高, 这可能与隐纹花松鼠的取食活动有关。而在尼西, 几种啮齿动物同时发生作用, 取食和贮藏都占一定的比例。3个地点种子贮藏的情况则更为复杂。相对于2006年, 2007年在明永和维西贮藏的比例极低, 而尼西的贮藏比例却没有显著差异(图2)。啮齿动物贮藏种子的比例往往取决于生境内食物的丰富度, 在种子产量越高的生境, 贮藏的比例越高(Forget & Milleron, 1991; Forget, 1992)。在尼西,可能因为啮齿动物数量较少(表 2), 对食物的相对需求较低, 年份变化对食物相对丰富度的影响不大,从而导致在2007年贮藏比例相对于2006年没有显著变化。而2007年在其它两个地点, 虽然在观察过程中, 有一定数量的种子被一次和多次贮藏(表 1),但在实验结束时, 绝大部分的贮藏的种子又被取出后食用, 可能是由于这两个点的啮齿动物数量多,食物相对匮乏所致。
3.4 不同年份种子命运的比较
不同年份种子被移动的速率以及种子命运存在差异, 2007年种子被移动速率更高(图1), 被移动后取食的种子比例更高, 被贮藏的比例更低(图2)。但是, 年份本身并不是影响种子命运的因素, 真正起作用的是种子产量、啮齿动物组成及其相对密度等因素。研究表明, 啮齿动物常常在种子产量小的年份取食更多的种子, 和种子表现为对抗关系, 而在种子产量大的年份贮藏更多的种子, 而表现为和植物的互惠关系(Vander Wall, 2002; Jansen et al, 2004)。在两年的研究中我们发现, 相对于2006年,当年结实的球果数量在 2007年显著的降低, 种子的减产可能造成啮齿动物食物短缺, 从而导致啮齿动物更多的取食而不是贮藏种子。种子减产可能的原因有两个:第一, 这可能和松树的一种繁殖策略有关, 即每隔几年有种子产量的丰年和歉年的变化,如中国东北地区的红松就有 3~5年的丰歉年的周期(Lu, 2002); 第二, 根据调查, 当年秋季迪庆地区雨水偏多, 妨碍了华山松的传粉和结实, 导致了华山松种子的减产。
致谢:思茅师范高等专科学校石青鹏、徐昌德、谭玉金帮助进行野外实验; 香格里拉高山植物园方震东、马宗灵、李红、和光、肖茂荣提供技术和交通支持; 香格里拉县汤满村王志刚夫妇、维西县及其落林工站李氏父子、德钦县明永村嘎太永宗和扎西夫妇提供后勤支持; 版纳植物园吴德林鉴定啮齿动物标本; 审稿专家对本文提供宝贵修改意见。在此一并致谢。
Benkman CW. 1995. Wind dispersal capacity of pine seeds and the evolution of different seed dispersal modes in pines [J].Oikos, 73: 221-224.
Brewer SW. 2001. Predation and dispersal of large and small seeds of a tropical palm [J].Oikos, 92: 245-255.
Chambers JC, MacMahon JA. 1994. A day in the life of a seed: movements and fates of seeds and their implications for natural and managed systems [J].Annu Rev Ecol Syst, 25: 263-292.
Chang G, Xiao ZS, Zhang ZB. 2009. Hoarding decisions by Edward’s long-tailed rats (Leopoldamys edwardsi) and South China field mice (Apodemus draco): the responses to seed size and germination schedule in acorns [J].Behav Proc, 82: 7-11.
Clarkson K, Eden SF, Sutherland WJ, Houston AI. 1986. Density dependence and magpie food hoarding [J].J Anim Ecol, 55: 11-121.
Forget PM, Milleron T. 1991. Evidence for secondary seed dispersal by rodents in Panama [J].Oecologia, 87: 596-599.
Forget PM. 1992. Seed removal and seed fate inGustavia superba(Lecythidaceae) [J].Biotropica, 24: 408-414.
Jansen PA, Hemerik L, Bongers F. 2004. Seed mass and mast seeding enhance dispersal by a neotropical scatter-hoarding rodent [J].Ecol Monogr, 74: 369-389.
Khan ML. 2004. Effects of seed size and seedling success inArtocarpus heterophyllusL., a tropical tree species of northeast India [J].Acta Oecol, 25: 103-110.
Levey DJ. 1987. Seed size and fruit-handling techniques of avian frugivores [J].Am Nat, 129: 471-485.
Lu CH. 2002. Hoarding behavior of Eurasian Nutcracker (Nucifraga caryocatact) and its role in seed dispersal of Korean pine (Pinus koraiensis) [J].Acta Zool Sin, 48(3): 317-321. [鲁长虎. 2002.星鸦的贮食行为及其对红松种子的传播作用. 动物学报, 48(3): 317-321.]
Moegenburg SM. 1996.Sabal palmettoseed size: causes of variation, choice of predators, and consequences for seedlings [J].Oecologia, 106: 539-543.
Moermond TC, Denslow J. 1985. Neotropical avian frugivores: patterns of behavior, morphology and nutrition, with consequences for fruit selection [J].Ornithol Monogr, 36: 865-897.
Moles AT, Westoby M. 2004. Seedling survival and seed size: a synthesis of the literature [J].J Ecol, 92: 372-383.
Pizo MA, Oliveira PS. 2001. Size and lipid content of nonmyrmechorous diaspores: effects on the interaction with litter-foraging ants in the Atlantic rain forest of Brazil [J].Plant Ecol, 157: 37-52.
Smallwood PD, Peters WD. 1986. Grey squirrel food preferences: The effects of tannin and fat concentration [J].Ecology, 67:168-174
Smith CC, Fretwell SD. 1974. The optimal balance between size and number of offspring [J].Am Nat, 108: 499-506.
Stapanian MA, Smith CC. 1978. A model for seed scatter-hoarding: coevolution of fox squirrels and black walnuts [J].Ecology, 59: 884-898. Theimer TC. 2001. Seed scatter-hoarding by white-tailed rats: consequences for seedling recruitment by an Australian rain forest tree [J].J Trop Ecol, 17: 177-189.
Theimer TC. 2003. Intraspecific variation in seed size affects scatter-hoarding behavior of an Australian tropical rain-forest rodent [J].J Trop Ecol, 19: 95-98.
Tomback DF, Linhart YB. 1990. The evolution of bird-dispersed pines [J].Evol Ecol, 4: 185-219.
Tomback DF, Schoettle AS, Chevalier KE, Jones CA. 2005. Life on the edge for limber pine: seed dispersal within a peripheral population [J].Ecoscience, 12: 519-529.
vander Wall SB. 1992. The role of animals in dispersing a ‘wind-dispersed’pine [J].Ecology, 73: 614-621.
Vander Wall SB. 1995. The effects of seed value on the caching behavior of yellow pine chipmunks [J].Oikos, 74: 533-537.
Vander Wall SB. 2002. Masting in pines alters the use of cached seeds by rodents and causes increased seed survival [J].Ecology, 83 :3508-3516.
Vander Wall SB. 2003. Effects of seed size of wind-dispersed pines.Pinus. on secondary seed dispersal and the caching behavior of rodents [J].Oikos, 100: 25-34.
Wang B, Chen J. 2009. Seed size, more than nutrient or tannin content, affects seed caching behavior of a common genus of old world rodents [J].Ecology, 90: 3023-3032.
Wang B, Yang XD. 2007. Seed predation ofApodemus latronumon 18 plant species in Northwest Yunnan [J].Zool Res, 28(4): 389-394. [王博, 杨效东. 2007. 大耳姬鼠对滇西北18种植物种子的捕食. 动物学研究, 28(4): 389-394.]
Wenny DG. 2000. Seed dispersal, seed predation, and seedling recruitment of a neotropical montane tree [J].Ecol Monogr, 70: 331-351.
Xiao ZS, Jansen PA, Zhang ZB. 2006. Using seed-tagging methods for assessing post-dispersal seed fate in rodent-dispersed trees [J].For Ecol Manage, 223: 18-23.
Xiao ZS, Zhang ZB,Wang YS. 2005. Effect of seed size on dispersal distance in five rodents-dispersed fagaceous species [J].Acta Oecol, 28: 221-229.
Xiao ZS, Zhang ZB. 2004. Hoarding behavior of rodents and plant seed dispersal [J].Acta Theriol Sin, 24(1): 61-70. [肖治术, 张知彬. 2004.啮齿动物的贮藏行为与植物种子的扩散. 兽类学报, 24(1): 61-70.]
Xiao ZS, Zhang ZB. 2006. Tin-tagging method, a powerful method for tracking seeds dispersed by rodents [J].Chn J Ecol, 25(10): 1292-1295. [肖治术, 张知彬. 2006. 金属片标签法—一种有效追踪鼠类扩散种子的方法. 生态学杂志, 25(10): 1292-1295.]
Zhang HM, Chen Y, Zhang ZB. 2008. Differences of dispersal fitness of large and small acorns of Liaodong oak (Quercus liaotungensis) before and after seed caching by small rodents in a warm temperate forest, China [J].For Ecol Manage, 255: 1243-1250.
Effects ofPinus armandiiseed size on rodents caching behavior and it’s spatio-temporal variations
CHEN Fan1,2, CHEN Jin1,*
(1.Key Laboratory of Tropical Forest Ecology,Xishuangbanna Tropical Botanical Garden,the Chinese Academy of Sciences,Mengla666303,China; 2.Graduate School of the Chinese Academy of Sciences,Beijing100049,China)
Pinus armandii, a native pine species, has large (about 300 mg), wingless seeds, and distributes from central to western China at an altitude of 1 000 - 3 300 m. To determine how the seed size affects rodents caching behavior, tagged seed releasing and tracking experiments were conducted at 3 sites in Northwest Yunnan province in 2006 and 2007. Our data indicated that for all sites and both years, compared with the smaller seeds, the proportions of cached large seeds were significantly higher, whereas the consumed ones were significantly lower. Meanwhile, the mean and maximum values of caching distances were also significantly increased in large seeds. Seed fate was different between the two years and within the three sites as there have different rodent community compositions.
Pinus armandi; Rodents; Seed dispersal; Seed fate; Scatter-hoarding; Caching distance
Q949.665; Q959.837;Q958..122.5
A
0254-5853-(2011)04-0435-07
10.3724/SP.J.1141.2011.04435
2010-11-12;接受日期:2011-05-12
“973”项目 (2007CB411603)
∗通讯作者(Corresponding author),E-mail: cj@xtbg.org.cn
猜你喜欢
中国-东盟博览(政经版)(2021年11期)2021-12-09
海外星云(2021年6期)2021-10-14
动漫星空(兴趣百科)(2020年5期)2020-06-10
娃娃乐园·绘本(2019年4期)2019-05-07
戏剧之家(2018年25期)2018-09-21
花火A(2017年11期)2017-12-16
消费导刊(2016年7期)2017-08-12
大观(2016年6期)2016-07-05
兵团工运(2016年1期)2016-01-31
读者(乡土人文版)(2015年7期)2015-12-13

- Zoological Research的其它文章
- 悬尾应激对小鼠空间记忆及其反转学习的损伤效应
- Visual modeling reveals cryptic aspect in egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth’s Leaf Warbler (Phylloscopus reguloides)
- Afferent and efferent pathways in the visual system of the freshwater snail Planorbarius corneus
- Notch signaling dependent differentiation of cholangiocyte-like cells from rhesus monkey embryonic stem cells
- Metabolism and thermoregulation between Mrs Hume’s Pheasant (Syrmaticus humiae) and Elliot’s Pheasant (S. ellioti)
- Localization of stationary pronuclei during conjugation of Paramecium as indicated by immunofluorescence staining