
黄淮麦区小麦品种F2籽粒中HMW-GS的遗传表现
2011-12-23 04:05张建刚潘登奎
山西农业科学 2011年4期
张 丽,张建刚,潘登奎
(山西农业大学文理学院,山西太谷030801)
小麦是最重要的粮食作物之一,占世界人口主要粮食需求的35%以上[1]。高分子量谷蛋白亚基(HMW-GS)类型与小麦加工品质密切相关[2-7]。HMW-GS 是由小麦的同源染色体上的3 个复合位点控制的,分别位于染色体1A,1B,1D 长臂的近着丝点处,被命名为Glu-A1,Glu-B1,Glu-D1位点,每个位点由2 个紧密连锁的基因组成[8]。理论上,普通小麦的每一品种有6 个不同的HMW-GS,但由于某些位点的基因不表达或处于沉默状态,多数小麦品种只有3 ~5 个HMW-GS,其中Glu-A1 位点编码0~1 个亚基,Glu-B1 位点编码1~2 个亚基,Glu-D1 位点编码2 个亚基[9]。
为给小麦品质改良提供依据,本研究从3 个位点都有差异的HMW-GS 基因组合进行杂交配组,收获籽粒后利用SDS-PAGE 方法对其F2籽粒HMW-GS 进行分析,目的在于了解位点的差异对F2分离规律的影响是否相同,从而为杂种小麦的加工品质预测以及品质配组选亲提供依据。
1 材料和方法
1.1 材料
母本为安农98005,其HMW-GS 组成为Glu-A1c(无显带),Glu-B1h(14+15),Glu-D1d(5+10)。父本为运84-20,其HMW-GS 组成为Glu-A1a (1 号带),Glu-B1c(7+9),Glu-D1a(2+12)。亲本及其F2籽粒均由山西省农业科学院棉花研究所提供。所测材料均采用单粒传(SSD)育种技术培育。
1.2 方法
本研究按照小麦高分子量谷蛋白亚基的SDS-PAGE 方法[10]进行,采用北京六一仪器厂DYCZ-24D 型8 cm×10 cm 双垂直电泳槽,样品梳为10 槽或15 槽。取单颗籽粒,于无胚端切取约10 mg,钳碎置于1.5 mL 离心管中,加入150 μL提取液,振荡10 min,静置过夜。
分析采用垂直板聚丙烯酰胺凝胶电泳,分离胶10% ,浓缩胶3.5% ,电压为70 V,电泳3~4 h,染色,照相。本系统亚基编号按Payne 等[11]方法进行。
2 结果与分析
共测300 粒F2种子,采用PIP 法判读HMW-GS 带型,其中带型可辨的有291 粒,部分安农98005×运84-20 F2籽粒麦谷蛋白SDSPAGE(10%)图谱如图1 所示,其余9 粒带型不能肯定就不进行进一步研究。

2.1 F2 籽粒中出现33 种HMW-GS 带型组合
由表1 可知,共有33 种不同的HMW-GS 带型组合,其中回归母本带型(N,14+15,5+10)的占3.78%,回归父本带型(1,7+9,2+12)的占5.15%,其余91.07%为双亲杂合带型或缺失带型。
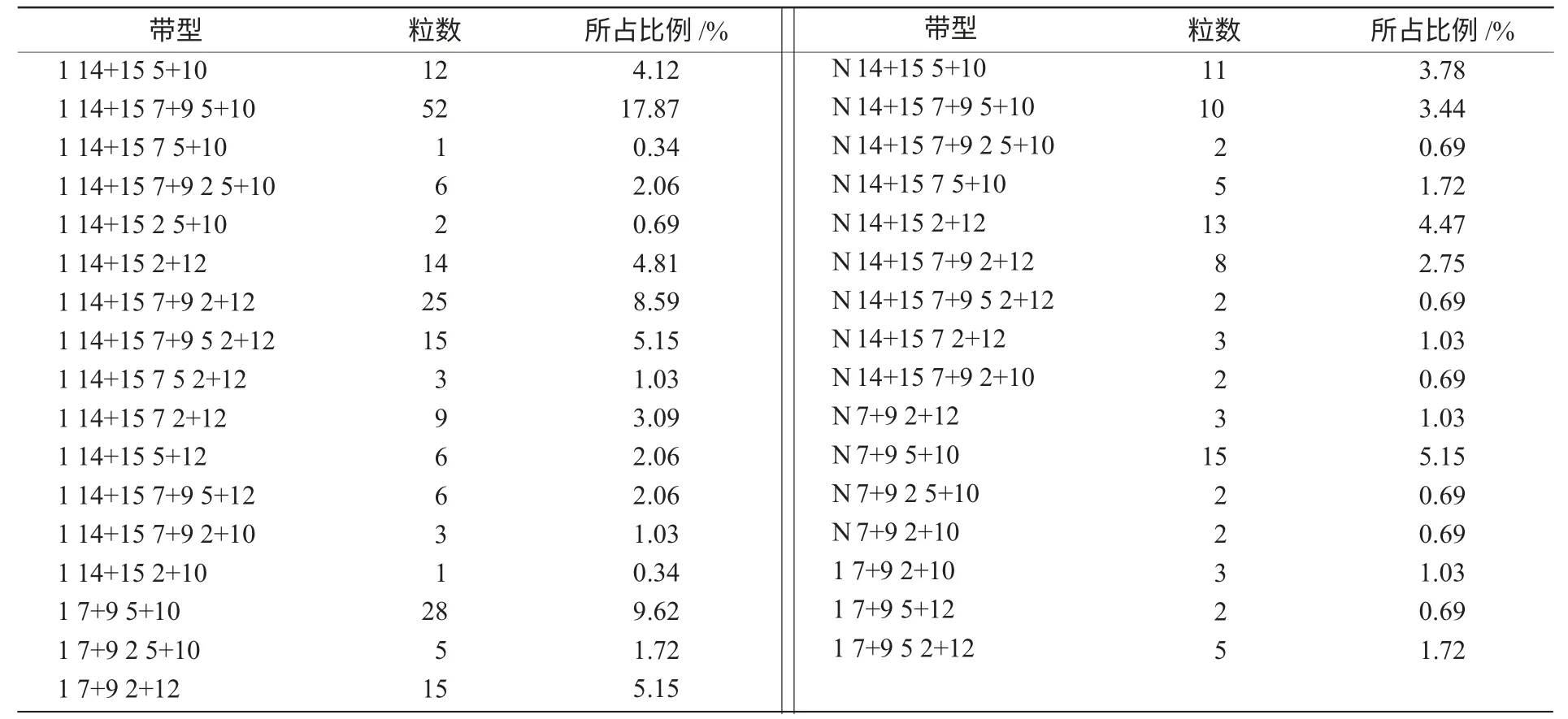
表1 安农98005×运84-20的F2 籽粒的HMW-GS带型统计
2.2 Glu-A1 位点上HMW-GS 的遗传表现及特点
在Glu-A1 位点上,母本安农98005 基因型为Glu-A1c(不显带),父本运84-20 基因型为Glu-A1a,编码1 号亚基。由表1 还可知,显1 带的有213 粒,无1 带的有78 粒,经χ2检验,二者分离比符合3∶1。Glu-A1a 与Glu-A1c 为1 对相对基因,由表性比3∶1 可知,其遗传规律符合1 对相对基因的分离规律,即Glu-A1a 与Glu-A1c 的分离比为1∶1。
2.3 Glu-B1 位点上HMW-GS 的遗传表现及特点
在Glu-B1 位点上,母本安农98005 基因型为Glu-B1h,编码14+15 号亚基;父本运84-20基因型为Glu-B1c,编码7+9 号亚基。从表1 可以看出,显7+9 带的有211 粒,显14+15 带的有211 粒,经χ2检验,二者分离比符合1∶1。另外发现,在本组合中出现了21 粒只显7 号带而无9 号带亚基的籽粒,占所测籽粒的7.22%,Glu-B1c 的表达空位率为7.22%。同时还出现基因杂合,即14+15,7+9 号亚基同时存在,在本组合中出现了131 粒,占所测籽粒的45.02%。
2.4 Glu-D1 位点上HMW-GS 的遗传表现及特点
在Glu-D1 位点上,母本安农98005 基因型为Glu-D1d,编码5+10 号亚基,父本运84-20 基因型为Glu-D1a,编码2+12 号亚基。从表1 可以看出,显2+12 带的有115 粒,显5+10 带的有151 粒,经χ2检验二者分离比符合9∶7。但在本组合中出现了25 粒2+10 和5+12 号亚基的籽粒,这种基因重组占所测籽粒的8.59%。
3 讨论
在本研究材料中有一部分在Glu-B1 位点上存在基因杂合状态,即14+15,7+9 号亚基同时存在,这种现象说明这些位点可能在遗传上还不稳定,存在着潜在的遗传分化。利用基因杂合可以人工培育出多种优质高分子量麦谷蛋白组合在一起的材料,这既能为小麦的品质改良及育种提供一定理论基础,同时也可为培育小麦优质品种增加新的材料。
小麦在遗传上有它自己的特点和复杂性,很少受环境因素的影响,每个染色体上2 个连锁基因重组率非常低,在Glu-D1 上,出现了5+12,2+10 号亚基的重组类型。目前,5+12 被认为是比5+10 更优质的亚基。而且在本试验中有三亚基组合的形式出现,即2+5+10,2+5+12 等亚基出现,所以可利用连锁基因重组和三亚基人工合成来改良小麦品质,为培育小麦新品种增加新材料。
还有一部分只显7 号带而无其他HMW-GS的籽粒,即应该同时显现的带却未显现。HMW-GS 表达空位尽管几率很小、原因不明,但其优点也较为明显,对消除所有的HMW-GS 带来说可能是非常有用的。
[1]李硕碧,高翔.小麦高分子量谷蛋白亚基与加工品质[M].北京:中国农业出版社,2001.
[2]Payne P I.Genetics of wheat storage protein and the effect of allelic variation in bread making quality [J]. Plant Physiol,1987,38:141-153.
[3]Kanenori Takata,Hiroaki Yamauchi,Zenta Nishio,et al.Effect of high-molecular-weight glutenin subunits with different protein contents on bread-making quality [J]. Food Sci Technol Res,2002,8(2):178-182.
[4]Takata K,Yamauchi H,Nishio Z,et al. Effect of high molecular weight glutenin subunits on bread-making quality using near-isogenic lines[J].Breed Sci,2000,50:303-308.
[5]张建华,张定一,姬虎太,等.山西不同时期小麦品种的高分子量麦谷蛋白亚基组成分析[J]. 山西农业科学,2008,36(4):39-41.
[6]谢飒英,谢三刚,宋昱,等.31 份国外小麦品种的HMW-GS组成分析[J].河南农业科学,2010(6):16-17,21.
[7]张之为,张立平,赵昌平,等.二系杂交小麦HMW-GS 组成与品质关系的初步研究 [J]. 华北农学报,2009,24(1):149-153.
[8]Harberd N P,Bartels D,Thompson R D. DNA restriction fragment variation in gene family encoding HMW-GS of wheat[J].Biochem Genet,1986,24:579-596.
[9]Ford J,Nalpiea J M,Halford N G. The nucleotide sequence of a HMW-GS gene located on chromosome 1A of wheat(Triticum aestivum L.)[J].Nucleic AcidsRes,1985,13:6817-6832.
[10]张丽,张建刚,潘登奎,等.小麦高分子量麦谷蛋白亚基的SDS-PAGE 方法[J].生物技术通报,2006(2):78-80.
[11]Payne P I,Lawrence G J. Catalogue of alleles for the complex gene loci,Glu-A1,Glu-B1 and Glu-D1 which code for high-molecular-wheat subunits of gulutenin in hexaploid wheat [J]. Cereal Research Communications,1983,11(1):29-35.
猜你喜欢
现代畜牧科技(2021年4期)2021-12-05
现代畜牧科技(2021年10期)2021-11-19
世界最新医学信息文摘(2020年68期)2020-12-25
延安大学学报(医学科学版)(2019年1期)2019-03-29
河北农业科学(2018年2期)2018-07-26
中国粮油学报(2016年5期)2016-01-23
现代检验医学杂志(2015年6期)2015-02-06
实验动物与比较医学(2014年5期)2014-02-28
中国医学科学院学报(2013年6期)2013-03-11
中国糖料(2013年1期)2013-01-22
