
三个芜菁品种源/库活性变化差异的比较
2011-12-10 10:34刘秀印郭玲杨伟高疆生
塔里木大学学报 2011年4期
刘秀印 郭玲 杨伟 高疆生,2*
(1 塔里木大学植物科学学院,新疆阿拉尔 843300)
(2 新疆生产建设兵团塔里木盆地生物资源保护与利用重点实验室,新疆阿拉尔 843300)
芜菁(Brassicarapa L)[1],大头菜,根如圆萝卜,华东一带通称香大头。十字花科(Cruciferae)芸薹属芸薹种芜菁亚种,能形成肉质根的二年生草本植物,肉质根肥大供食用(炒食、煮食或腌渍)。欧洲、亚洲和美洲均有栽培。在少数民族聚居的地方有着2000多年的栽培历史,是南疆许多地区少数民族每天必不可少的重要蔬菜。
对很多根菜类蔬菜来讲,源主要指的是叶源,库主要指的是肉质根。源是库的基础,同时库对源有反馈调节的作用,在产量形成中,源既参与库的建成,又参与库的充实,源-库具有相对性,作为源的地上部在一定时期为源,在另一时期就会转变成库,有的作物在一定时期收获的肉质根(库)又是生产源之一。
在1967年,Warren-Wilson(1967)提出了源/库活性计算的概念:源/库活性(source/sink activity)(ln w2—ln w1)/T[1],w1、w2 分别为植物始、末期的干重,T为连续两次测定的时间间隔。
源-库互作的机理[2]实际上是体内物质在源器官与库器官的转化、分配,促进体内物质向库器官中分配是源库关系协调、提高产量的关键。本试验旨在为完善芜菁的栽培技术,提高芜菁产量提供技术支持。
1 材料与方法
1.1 试验地概况
试验在新疆阿拉尔市塔里木大学园艺站进行。阿拉尔地处天山南麓、塔里木盆地边缘,该地区属温带大陆气候,光照时间长、昼夜温差大,年平均气温9.6~11.5 ℃[3],全年无霜期183 ~228 d,适宜多种农作物生长,试验地点土壤为壤土。

表1 供试土壤基本情况
1.2 试验材料
供试材料为3个芜菁品种:芜菁-1(柯坪地方品种)、芜菁-2(昌吉市亚华种苗有限责任公司培育品种)、芜菁-3(河南省邢阳种子有限责任公司培育品种)。
1.3 试验方法
试验于2011年8月16日至10月16日进行,3个品种同时播种,分别播于面积、土壤条件均相同的三个小区里,重复3次。各小区面积为60 m2,共180 m2,行距20 cm,定苗后株距15 cm,进行常规田间管理。从出苗后第7 d开始取样,对所取样株分别进行测定地上部和地下部鲜重、干重,肉质根直径(测量其最膨大部分的横茎)。每次每个品种分别取样株10株,每隔7 d取一次,直至收获,干物质质量测量方法为将相应材料先在105℃烘箱中杀青2 h[4],然后在 80 ℃烘至恒重。
1.4 数据处理
源活性(source activity)和库活性(sink acvtivity)=(ln w2—ln w1)/T[4]
采用Excel与DPS软件进行数据统计与分析。
2 结果与分析
2.1 不同品种肉质根直径和地下部鲜质量的变化
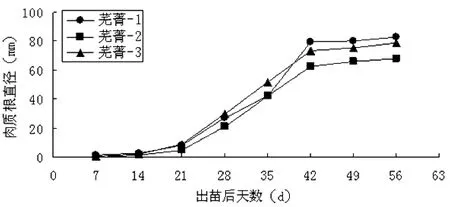
图1 不同品种肉质根直径的变化

图2 不同品种地下部鲜重的变化
试验结果发现,上述3个不同品种,其肉质开始膨大(破白[5])的时间开始于出苗后14~21 d,而在“破白”后至42 d,肉质根直径增长速度几乎都呈直线上升的趋势,42 d之后3个品种的变化都比较缓慢(如图1)。从图1看出,芜菁-2在整个生长季中其肉质根直径大小较其它两个品种偏小。
由图2可以看出,三个芜菁品种地下部鲜质量在出苗后21 d内几乎没变化,在21~35 d才开始缓慢增加,35 d后均开始快速增加,而在出苗后的49 d,地下部的根鲜质量增加又比较平缓。在35~49 d,芜菁-1的地下部鲜重呈直线增加;芜菁-3和芜菁-2在35~42 d均增长较快,在42~49 d芜菁-3增长速度有所减慢,而芜菁-2增长速度大幅减小,地下部鲜重变化不大。在49~56 d芜菁-3与芜菁-1地下鲜重几乎不变,而芜菁-2有所增加但其最终地下鲜重在三品种中为最小。
综合图1和图2可以看出,3个不同类型的芜菁品种,随着肉质根直径的增加,肉质根鲜质量增加,但肉质根鲜质量的增加迟于肉质根直径。3个品种地下鲜重增加与肉质根直径的增大变化趋势在整个生长季大体一致均呈单“S”形曲线。芜菁-2的肉质根直径是供试三个品种中最小,其地下部鲜重也为最小,芜菁-1与芜菁-3的肉质根直径较大,其地下鲜重也较大,这一点表明三个供试品种的地下部鲜重与肉质根直径之间存在很大相关性。
2.2 不同品种根冠比的变化

图3 不同品种根冠比变化图
图3可以明显看出,三个供试品种的根冠比(鲜质量)在出苗后21 d内变化较小,之后则呈现增加的趋势。另外,从出苗至35 d三个供试品种的根冠比变化趋势大体保持一致,第35 d后出现分歧。芜菁-3与芜菁-2均在42 d后开始快速增长,而芜菁-1稍早,在35 d后快速增长且增长趋势呈直线。
在3个芜菁品种从出苗到收获,随着地上部与地下部鲜重不断的相对变化,根冠比逐渐接近1。从图3可明显看出芜菁-2最终的根冠比稍大于1,芜菁-3的接近0.9,芜菁-1的相比要小些接近0.7。
综合图2与图3可看出,三个供试品种的地下部鲜重变化趋势与根冠比变化趋势在出苗至第35 d明显一致,第35 d后三个供试品种这两方面的变化趋势呈相反状态。芜菁-1在第35 d~第49 d地下部鲜重呈直线增加,而其根冠比在第35 d~42 d较缓增加,之后在第42 d~49 d以较快速度增加,这说明其地上部鲜重在第35 d~42 d增加速度较快而后在第42 d~49 d有所减缓;在整个生长过程中,芜菁-2地下鲜重整体上为三者中最小但根冠比整体上却为三者中最大,这表明芜菁-2的地上鲜重在整株中所占的比例较其它两个品种的要小,该品种不仅地下部鲜重小,地上部鲜重相对更小;芜菁-3在第35 d~49 d地下部鲜重的变化趋势与根冠比变化趋势呈相反的关系,在第35 d~42 d其地下部鲜重增长速度较快而后在第42 d~49 d增长速度有所减慢,但其根冠比在这两个时期的变化分别为增长速度相比较缓而后加速增加,这表明在其地下部鲜重以较快速度增加时其地上部鲜重以相对更快的速度增加,当其地下鲜重以较缓速度增加时,地上部鲜重增加速度相对更缓,即其地上部鲜重变化较地下部鲜重变化更剧烈。对于芜菁而言,地上部是主要的源,地下部为主要的库。由此可知,芜菁-3这一品种地下部鲜重的变化与地上部鲜重变化有很大相关性,尤其是在第35 d~49 d这一时期内。
2.3 不同品种地下部与地上部干质量的变化
从图4可以看出,3个芜菁品种其地下部干质量的变化基本呈现一致的趋势。在出苗后35 d内,3个品种的地下部干质量增加很少,其增加主要发生在出苗35 d以后,并且出现了快速增长的趋势,同时,与地上部干质量存在较大的品种间差异不同的是,在出苗后49 d内,3个芜菁品种地下部干质量没有太大的差别,即使是出苗后56 d测定时,芜菁-3和芜菁-2的肉质根干质量也比较接近,芜菁-1的稍大于前者。
从图5可以看出,三个品种芜菁叶片干质量自出苗后7d~21 d变化均不大,自第21 d均开始以较快速度增加。第35 d后芜菁-3与芜菁-1增长速度大幅增长而后有所减小,而芜菁-2在35~42 d之间变化不大,但是在42 d之后增长速度陡然加快。3个供试品种地上部干重在第49 d~56 d增加速度变缓,最终地上部干重三个品种相差无几。

图4 不同品种地下部干重的变化

图5 不同品种地上部干重的变化

图6 不同品种源活性的变化

图7 不同品种库活性的变化
综合图4与图5可以看出,3个供试芜菁品种即芜菁-1、芜菁-2、芜菁-3,其地上部干质量从出苗至35 d增加速度较缓,但是在第35 d~49 d时增加迅速,而其地下部干重在最初35 d内几乎没什么变化,增加量很小,35 d后快速增加,这说明在最初35 d里以地上部干重积累为主。而后在第35 d~42 d地上部与地下部干重同时以较快速度增加,此期也是三个供试芜菁产量形成的重要时期。结合图2与图4可看出,三个供试品种虽地下鲜重存在较大差异但最终干重相差不大。芜菁-2的地下鲜重为三者中最小但其干重与其它两者相差不大,尤其是与芜菁-3,表明其干物质含量较其它两者高;芜菁-1与芜菁-3两者地下鲜重相当但地下干重芜菁-1高于芜菁-3,说明前者干物质含量高于后者。2.4 不同品种源活性与库活性的变化
2.4 芜菁源活性和库活性
源/库活性(source/sink activity)(ln w2一 ln w1)/T,其中w1、w2分别为始、末期的干重。(源/库活性没有具体单位,是个数值)。
从图6可以看出,不同芜菁品种,其生长初期(出苗后28 d内),源活性差别不大,且芜菁-1与芜菁-2在最初28内变化趋势基本一致,但自第28 d~35 d芜菁-1与芜菁-3的源活性变化相似,都以较快速度增加,而芜菁-2却以较快速度减小,在第35 d~42 d,三者的变化情况又与前一时期相反。在第42 d~49 d时,芜菁-2源活性快速减小,芜菁-1的减速较缓,但芜菁-3缓速增加。自出苗后21 d开始,肉质根开始膨大,源活性的品种间差异突现,其中芜菁-2从出苗后从第42 d开始呈现下降趋势,而芜菁-1的源活性与芜菁-3的源活性都是在出苗后35 d快速下降。图6明显反映出芜菁-1与芜菁-3的源活性高峰出现在第35 d,而芜菁-2的要迟些出现在第42 d。
在库活性方面,由图7可看出在出苗后28 d内,随着肉质根的缓慢生长,3个品种的库活性变化也不大,自第28 d三个品种的库活性开始增强但变化情况差异很大。芜菁-3和芜菁-1其库活性在出苗后42 d达到最大值而之后则呈现快速下降态势,但芜菁-2则在35 d达到最大值后出现快速下降趋势,这表明芜菁-2的源活性高峰出现在第35 d,早于芜菁-1与芜菁-3,后两者达到高峰时间出现在第42 d。
综合图6、图7可以看出,在3个供试芜菁品种中芜菁-1与芜菁-3的源活性与库活性呈现一致的趋势,源活性高峰均出现在第35 d,库活性高峰均出现在第42 d,这表明芜菁-1与芜菁-3在最初35 d以地上部物质积累为主,即以增强源为主,随着肉质根的生长,库活性逐扩大增强并在第42 d出现库活性高峰。而‘芜菁-2’的源活性高峰晚于前两者出现在第42 d,库活性高峰却早于前两者出现在第35 d,由此可以看出肉质根作为芜菁的贮藏器官,其生长落后于作为同化器官叶片的生长。
3 小结与讨论
芜菁肉质根的形成依赖于健壮的莲座叶的形成,叶片长势良好,同化作用旺盛,运输到肉质根的营养也就会越来越多,从而促进了肉质根的膨大。同时,芜菁肉质根的生长也逐渐改变了原来的源一库关系(Hole,1996)[6],并在一定程度上刺激了叶片器官的功能,提高同化产物合成和转运能力,进一步促进肉质根的膨大,提高芜菁的产量。
试验结果发现,三个品种即芜菁-1(柯坪品种)、芜菁-2(昌吉品种)、芜菁-3(河南品种)的生长状况基本相似,出苗后28 d内,地上部的生长较快,地下部生长缓慢,且根冠比小于0.2,出苗28 d后进入肉质根膨大期,根冠比增加,地下部干重明显增加。在源活性方面,3个芜菁品种在28 d之前基本处于缓慢增长的趋势,其后均有不同程度的降低和增长。在库活性方面,不同品种具有较为一致的变化态势。在出苗后28 d内,随着肉质根的生长,库活性快速增强,芜菁-1(柯坪品种)、芜菁-3(河南品种)库活性在45 d时最强,芜菁-2(昌吉品种)则35 d库活性最强,提前了10天。经测产3个品种的产量依次为:芜菁-3(柯坪品种)>芜菁-1(河南品种)>芜菁-2(昌吉品种)。
那么,在生产实践中,当芜菁出现“破白”时,要及时间苗、培土,少浇水,使直根深入土层[7]。在出苗后21 d~40 d时,源库活性非常旺盛,在这期间,芜菁地上部的生长量比苗期大大增加,此期需要较多的水肥供应,但要适量灌溉,防止叶片徒长。芜菁快要收货时,叶子继续生长,但生长速度逐渐减慢,这时应适当浇水,防止空心[8]。总之在生产上可利用库源关系采取相应措施,调节地上部与地下部的平衡生长,在不同的阶段采取不同的措施,从而达到提高产量,改善品质的目的。
[1]张振贤,梁书华.根菜类蔬菜肉质根形成生理研究进展[J].山东农业大学学报,1994,25(2):249 -25.
[2]满为群,杜维广,张桂茹.大豆高光效种质与高产品种源库平衡的研究[J].中国油料,1995,17(2):8 -11.
[3]周竹青,周广生.作物库、源、流生理机理研究进展[J].中国农学通报,2001,17(6):45 -48.
[4]刘克礼,高聚林,孙会忠,等.马铃薯源的供应能力与库容量的关系[J].中国马铃薯,2004,18(l):4 -8.
[5]刘建国,帕尼古丽,董志新,等.单位叶面积负荷量对大豆源库调节效应的研究[J].石河子大学学报,2003,7(4):259 -262.
[6]魏建军,刘建国,董志新,等.源库调节对大豆光合速率及同化产物运转分配影响的研究[J].新疆农业科学,2004,41(2):65 -68.
[7]刘晓冰,张秋英.试论作物的源库系统[J].农业系统科学与综台研究,1992,8(2):131 -136.
[8]卢钢,庄晓英,叶纵芝.萝卜肉质根膨大过程库活性与蛋白质变化研究[J].浙江大学学报(农业与生命科学版),2004,30(l):39 -43.
猜你喜欢
今日农业(2022年2期)2022-11-16
美食(2022年4期)2022-04-16
山西中医药大学学报(2020年2期)2020-06-06
美食(2020年5期)2020-06-01
中成药(2018年12期)2018-12-29
中国科技纵横(2018年14期)2018-08-30
浙江农林大学学报(2018年4期)2018-08-22
现代农业科技(2017年16期)2017-09-22
Asian Journal of Urology(2015年3期)2015-12-16
江苏农业科学(2014年1期)2014-07-18
