
短命植物异果芥的生殖分配及结实特性
2011-12-10 10:34吴雪莲王文华谭敦炎
塔里木大学学报 2011年4期
吴雪莲 王文华 谭敦炎
(1 农业部农产品质量监督检验测试中心,西藏拉萨 850032)
(2 西藏自治区农牧科学院蔬菜研究所,西藏拉萨 850032)
(3 新疆农业大学林学院,乌鲁木齐 830052)
植物的生殖和结实特性是植物生活史中的一个重要特征[1,2],在种子植物的生活史中,结实特性同时受内部遗传因素和外部物理、生物环境的影响[3]。植物的生殖分配和结实结籽特性深刻地影响着植物资源的分配效率、后代品质,对种群的发展壮大起着至关的重要[4]。因此,对植物生殖分配和结实结籽特性的研究对于揭示种子植物的繁殖特性以及种群建成具有非常重要的价值。异果芥(Diptychocarpus stricus)属于十字花科异果芥属(Diptychocarpus Trautv.),是异果芥属中唯一的一个种。在我国,异果芥仅分布于新疆北部,是准噶尔荒漠中很常见的早春短命植物[5]。异果芥的生长发育特点为:具有两种不同花色的植株;在其同一花序上存在着两种形态和开裂方式不同的果实,是十字花科中为数不多的具有异形果实(heteromorphy fruit)的种之一。因此,对短命植物异果芥的生殖分配和结实格局等生物学特性的研究,对于探讨其在准噶尔荒漠中的生态适应对策具有非常重要的生态学意义,同时为该种群能够在恶劣的准噶尔荒漠中的发展壮大提供理论依据。从而使该物种在治理荒漠、保护生态环境方面起到十分重要的作用。
1 实验材料和方法
1.1 实验材料及观测地点
实验材料异果芥为一年生早春短命植物。该种植物具有如下特点:种子萌发具典型的春、秋季萌发现象,萌发的植株具有白花和紫红色花两种类型;春季萌发的植株生活周期仅为70天左右;两种花色植株的同一花序上均能形成两种不同形态的果实,其中花序上部的果实扁平,开裂,成熟时不脱落;下部果近圆柱形,不开裂,成熟时脱落。由于白色花下部果实萌生植株数量较少,不能进行样本统计分析,所以本试验只选取了紫红色花上、下部果实萌生植株和白色花上部果实萌生植株为实验材料。
实验观察在新疆农业大学实验农场的短命植物资源圃中进行。农场位于新疆乌鲁木齐市西郊,北纬 43°48',东经 87°34',属于天山前山带冲积扇平原,为典型的荒漠型气候。实验地环境、土壤均与野外条件相似。
1.2 实验方法
1.2.1 测定生殖分配
在果实成熟期,每种植株随机选取30株,连根挖出,将果实单独剪下,与植株其他部分分别装入纸袋烘干(80 ℃,72 h),用 BS210S(max210 g,d=0.1 mg)电子天平称取干重。生殖分配按果实的生物量占植物总生物量的比率计算。
1.2.2 测定结实特性
在异果芥果实成熟期,每种植株随机选取30株,对一、二级分枝数/株、上、下部果实数/株、果实总数/株、上、下部果实败育率/株等进行统计。
1.3 数据处理
利用Excel软件对有关数据进行处理。应用SigmaPlot软件进行绘图。
2 实验结果
2.1 生殖分配及其相关性分析研究
成熟期春萌植株单株生物量及生殖分配的方差分析结果见表1。

表1 异果芥成熟期春萌株单株生物量与生殖分配间比较
由表1可知,紫红色花植株上萌生植株与下部果实萌生植株的单株生物量存在极显著差异(P<0.01),但两种植株的生殖分配差异不显著(P >0.05)。同时紫红色花和白色花植株上部果实萌生植株之间的单株生物量与生殖分配间均无显著差异(P >0.05)。
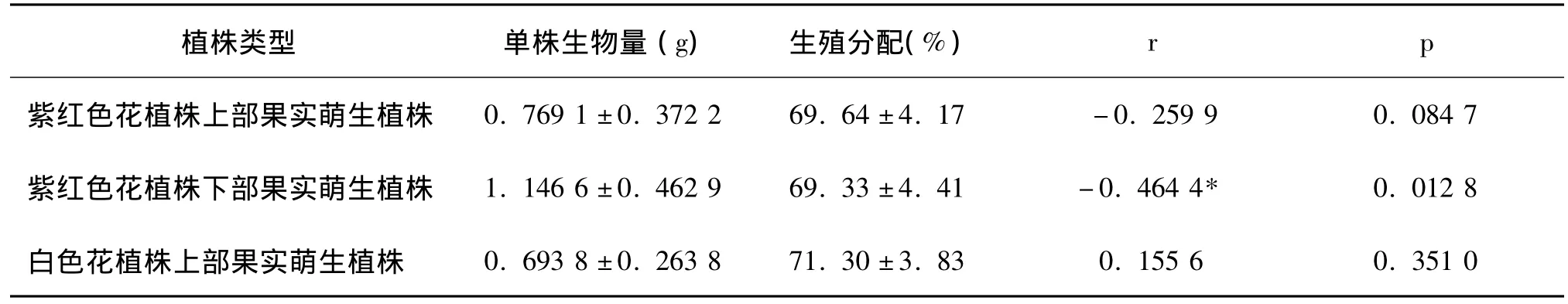
表2 异果芥成熟期春萌株单株生物量与生殖分配间的相关性
同时表2的相关性分析结果表明:紫红色花和白色花植株上部果实萌生植株的单株生物量与生殖分配间均无显著相关性(p>0.05)。紫红色花植株下部果实萌生植株的单株生物量与生殖分配间存在显著相关性(0.01<p <0.05)。
2.2 结实特性与相关分析
对三种植株繁殖构件与结实特性测定及统计分析结果如表2。

表3 异果芥春萌株繁殖构件特征与结实特性
表3结果表明:三种植株一、二级分枝数/株、上、下部果实数/株、果实总数/株等均存在极显著差异(P<0.01),为紫红色花下部果实萌生植株的分枝数最多、果实总数最多,紫红色花下部果实萌生植株的次之,而白色花上部果实萌生植株的最少。上、下部果实败育率/株差异不显著(P>0.05)。
三种植株单株生物量与单株结果数之间的关系如图1、图2、图3。
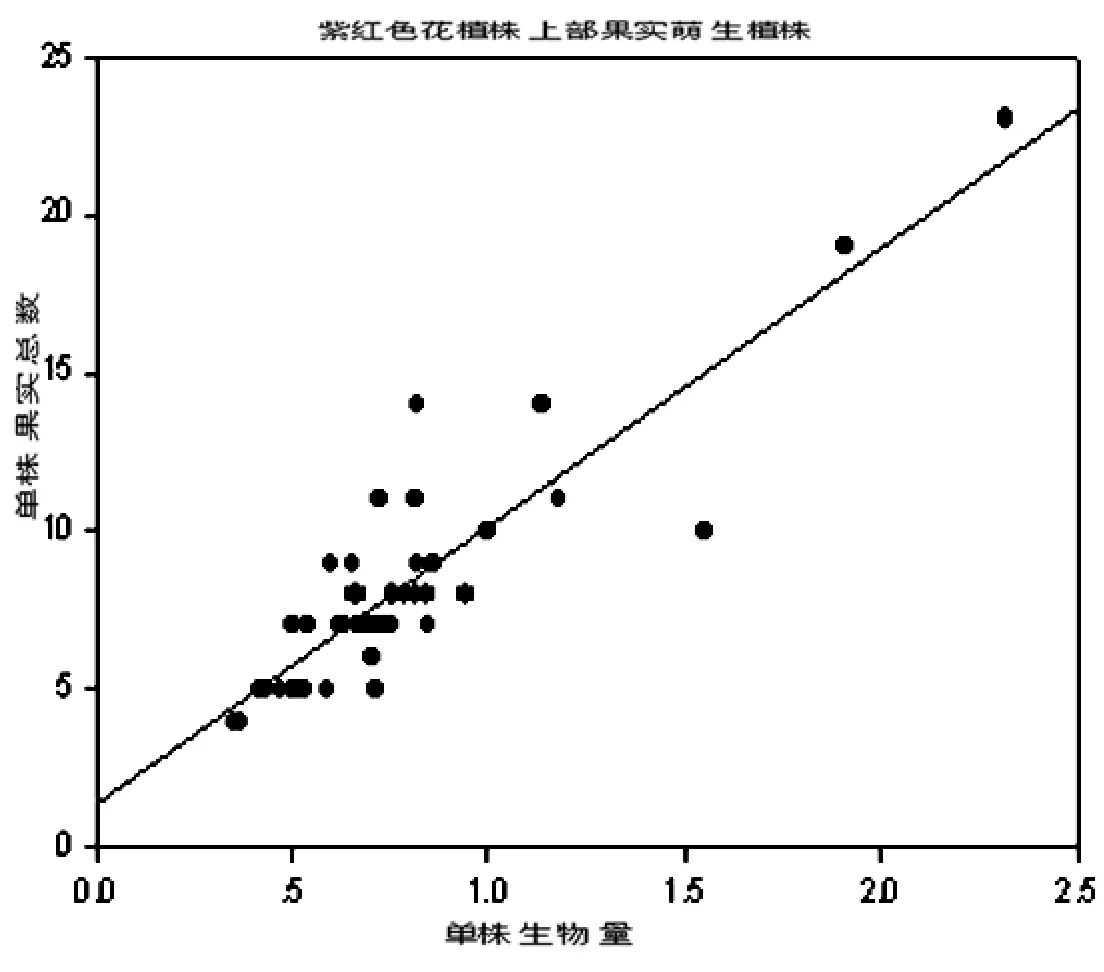
图1 异果芥紫红色花植株上部果实萌生植株结实数与单株生物量的相关关系(r=0.983,t< 0.001,在0.01水平显著相关。)
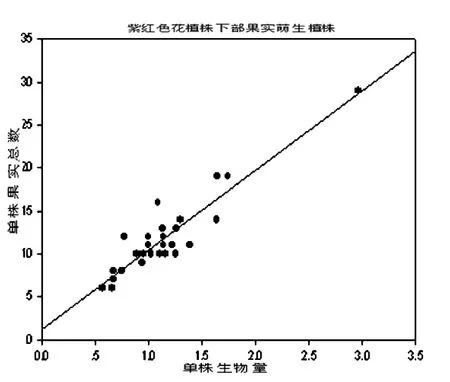
图2 异果芥紫红色花植株下部果实萌生植株结实数与单株生物量的相关关系(r=0.990,t<0.001,在0.01水平显著相关。)

图3 异果芥白花植株上部果实萌生植株结实数与单株生物量的相关关系(r=0.981,t< 0.001,在0.01水平显著相关。)
结果表明,异果芥三种植株的结实数与单株生物量呈线性正相关。说明单株生物量越大,单株果实数越多。即植物植株的个体越大,产生的果实数量就越多(r值分别为0.983,0.990,0.981;t<0.001)。
3 讨论
3.1 生殖分配与生态适应对策
繁殖分配是一种自然选择。一次结实草本植物的生殖分配一般占其总生物量的20% ~40%,多次结实植物则占每年总生物量的20%以下[6,7]。在本研究中,异果芥三种春萌株的单株总生物量之间虽然存在显著差异,但其生殖分配差异不显著(表1),均可达65%以上,明显高于其他已报道的一次结实的草本植物。单株生物量与生殖分配之间相关性不显著(表2),表明它们的生殖分配比较稳定,不会随着植株生物量的改变而改变。从它们的生存环境来看,新疆准噶尔荒漠生境条件非常恶劣,多风、干旱、温度变化大。异果芥在严酷、恶劣环境条件的影响下,倾向于高的生殖分配。本文所研究的这种短命植物具有生殖分配高、生长周期短的特征,为较典型的r-生活史对策,即表现为快速发育、小型成体、数量多而个体小的后代,高的繁殖能量分配和短的世代时间。因此,较高的生殖分配是这种短命植物与新疆准噶尔荒漠环境长期适应的结果。
3.2 结实特性与果实多态性
植株的结实率、种子产量与植物的繁殖能力的关系很大[8]。从结实特性看,异果芥这种短命植物结实数量较多,为该物种的生存发展提供更多机会。同时,三种植株在单株生物量、结实数上存在差异,其单株生物量都与结实数显著正相关(图1,图2,图3),即植株个体越大,产生的果实数量越多。说明这种植物在繁殖过程中,能很好的利用资源优势,尽可能多的产生果实。此外,异果芥不同形态的果实(种子)萌生植株所表现出的单株生物量、结实特性上的差异,使之形成独特的适应机制—果实多态性[9]。许多植物的多态果实在大小、果皮厚度等都存在明显不同,使其在散布、休眠及萌发等生态学行为也上存在差异,是一种混合生殖对策,可以减小环境变化对植株生殖成功的影响[9,10]。面临环境变异,植物会演化出最佳的生殖对策,以达到对局部环境条件的最佳适应,这种适应常常表现在生活史特征或生殖方式的变异上[11]。植物的生殖过程受环境影响,反之,在变化的环境中,植物会通过某些生态反应来保证生殖成功。异果芥的果实多态性正是一种生态适应策略。
[1]方炎明.植物生殖生态学[M].济南:山东大学出版社,1996,18 -21,132 -147.
[2]孙凡,钟章成.四川大头茶繁殖分配及其与环境适应性的关联度研究[J].植物生态学报,1997,21(1):44-52.
[3]Sutherland S.Patterns of fruit- set:what controls fruitflower ratios in plants?[J].Evolution,1986,40(1):117-128.
[4]熊志斌,冉景丞,谭成江,等.濒危植物掌叶木种子生态特征[J].生态学报,2003,23(4):820-825.
[5]毛祖美,张佃民.新疆北部早春短命植物区系纲要[J].干旱区研究,1994,11(3):1-26.
[6]祖元刚,张文辉,阎秀峰,等.濒危植物裂叶沙参的保护生物学[M].北京:科学出版社,1999.
[7]姚红,谭敦炎.胡卢巴属四种短命植物个体大小依赖的繁殖输出与生活史对策[J].植物生态学报,2005,29(6):954-960.
[8]Susko D J,Lovett-Doust L.Plant- size and fruit- positioneffectsonreproductiveallocationinAlliaria petiolata(Brassicaceae)[J].Canadian Journal of Botany,2000,78:1398-1407.
[9]Imbert E.Ecological consequences and ontogeny of seed heteromorphism[J].Perspectives in Plant Ecology,Evolution and systematics,2002,5(1):13 -36.
[10]Venable D L.The evolueionary ecology of seed heteromorphism[J].The American Naturalist,1985,126(5):577-595.
[11]Harper J L.The population biology of plants[M].London:Academic Press,1977.
猜你喜欢
中国生殖健康(2020年8期)2021-01-18
中国生殖健康(2020年2期)2021-01-18
阅读(快乐英语高年级)(2019年8期)2019-09-10
中国生殖健康(2018年1期)2018-11-06
中国生殖健康(2018年2期)2018-11-06
丝绸(2018年9期)2018-09-10
小火炬·智漫悦读(2018年12期)2018-03-13
今古传奇·故事版(2017年24期)2018-02-07
莫愁(2017年36期)2017-12-25
海外英语(2013年10期)2013-12-10
