
PAEs污染下2种基因型菜心根际土壤微生物特征及差异性
2011-10-20 02:04:52曾巧云莫测辉蔡全英华南农业大学资源环境学院农业部生态农业重点开放实验室广东广州50642暨南大学环境工程系广东广州50632
中国环境科学 2011年3期
曾巧云,莫测辉,蔡全英(.华南农业大学资源环境学院,农业部生态农业重点开放实验室,广东 广州50642;2.暨南大学环境工程系,广东 广州 50632)
PAEs污染下2种基因型菜心根际土壤微生物特征及差异性
曾巧云1,莫测辉2*,蔡全英1(1.华南农业大学资源环境学院,农业部生态农业重点开放实验室,广东 广州510642;2.暨南大学环境工程系,广东 广州 510632)
设置邻苯二甲酸酯(PAEs)3个污染水平(T1,T2,T3)和对照处理(T0),盆栽2种基因型菜心(油青60天菜心和特青60天菜心),研究2种菜心不同生长期根际土壤微生物生物量碳氮(Cmic、Nmic)和总体代谢活性(AWCD)的动态变化特征及差异性,初步揭示2种菜心吸收累积PAEs差异的根际微生态机理.结果表明,2种菜心根际土壤 Cmic分别介于 84.45~213.30mg/kg和 98.33~229.85mg/kg,Nmic分别介于9.00~25.86mg/kg和12.69~34.36mg/kg,显著高于未种植物的处理. AWCD值分别介于0.744~1.075和0.847~1.318,特青60天菜心显著高于未种植物的处理. 2种基因型菜心相比,特青60天菜心根际土壤Cmic, Nmic以及AWCD值均大于油青60天菜心(T3处理开花期除外),部分处理间差异显著. 2种基因型菜心茎叶和根系中PAEs含量与Cmic、Nmic和AWCD之间的相关性均未达到显著程度,说明2种基因型菜心茎叶和根系中PAEs含量可能与其他因素有关.
邻苯二甲酸酯(PAEs);菜心(Brassica parachinensis);土壤;微生物生物量;微生物功能多样性
我国一些地区农业土壤中邻苯二甲酸酯(PAEs)浓度达到几个至十几个 mg/kg,超过了美国有关土壤控制标准[1-2],对土壤微生态和植物生长产生不利影响[3-5],而且还可能累积在植物体内[6-9],危及农产品安全和人类健康.微生物降解是自然环境中 PAEs完全矿化的主要途径[10].根系分泌物能为根际微生物的生长提供碳源,因此植物对根际微生物的生长具有一定的促进作用[11].但不同植物类型或同一植物不同基因型根系分泌物存在着差异,直接影响根际微生物数量、群落结构和代谢功能多样性[12],以及根际环境中污染物的生物有效性[13],从而导致污染物被植物吸收累积及被微生物降解的差异性.因此,研究污染条件下不同植物或同一植物不同基因型根际土壤的微生物特性,对明确污染物在土壤-植物系统中的环境行为具有重要意义.目前关于PAEs污染下植物对土壤中微生物影响的研究鲜见报道.因此,本研究应用已筛选出的PAEs高/低吸收累积基因型菜心(油青 60天菜心和特青 60天菜心)[8-9],以环境中普遍检出的2种PAEs化合物(邻苯二甲酸正二丁酯(DBP)和邻苯二甲酸二(2-乙基己基)酯(DEHP))为研究对象,研究 PAEs污染水平下 2种基因型菜心不同生长期根际土壤微生物生物量和代谢功能多样性的动态变化及其差异性,初步揭示 2种基因型菜心吸收累积PAEs差异的根际微生态机理.
1 材料与方法
1.1 供试材料
供试植物为前期试验所筛选出的对 PAEs高吸收累积基因型菜心(油青60天菜心,简称油青 60)和对 PAEs低吸收累积基因型菜心(特青60天菜心,简称特青 60)[8-9],种子购自广东省农业科学院.
供试化合物DBP和DEHP,为分析纯,均购自广州化学试剂厂.
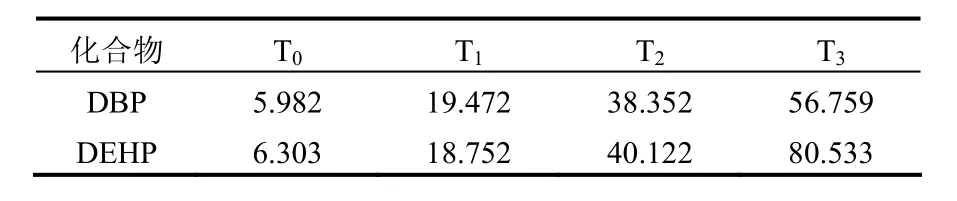
表1 盆栽前土壤中PAEs化合物的初始含量(mg/kg)Table 1 Initial concentrations of DBP and DEHP in soil(mg/kg)
供试土壤为水稻土,采自华南农业大学农场,其pH 5.86,有机质含量为13.5g/kg,全氮、全磷和全钾含量分别为1.00g/kg、1.38g/kg和32.16g/kg.土壤经自然风干过 5mm 筛后备用.设置 3个PAEs人工污染水平(即T1,T2,T3,DBP和DEHP的浓度同时分别为20,50,100mg/kg)和对照处理(即T0,土壤中不添加 PAEs),污染土壤的配制参考文献[8-9].盆栽前土壤中DBP和DEHP的初始含量见表1.
1.2 盆栽与采样
盆栽试验在华南农业大学资源环境学院网室中进行.试验设置未种植物处理和种植 2种基因型菜心处理,用根垫法(60目不锈钢丝网)区分根际、非根际土壤,其中根际土壤为 2kg,非根际土为3kg.每个处理重复6次,随机区组排列.盆栽期间用蒸馏水浇灌(以不渗漏为准),人工防治虫害,不喷施农药.
分别于菜心的幼苗期(播种后第 15d)、叶片生长期(播种后第 25d)、菜苔形成期(播种后第39d)和开花期(播种后第52d)采集根际土壤样品.根际土壤采集的具体操作为采集不锈钢丝网以上,菜心根基周围2cm的土壤,去除根系及其它杂物,置于0~4℃的冰箱备用.
1.3 测定项目与方法
土壤微生物生物量碳(Cmic)和微生物生物量氮(Nmic)的测定采用氯仿熏蒸-K2SO4浸提法[14].土壤微生物功能多样性采用Biolog ECO板进行测定,按照文献[15]报道的方法:称取相当于 10g烘干土壤的根际土壤,加入到装有100mL灭菌生理盐水(0.85%)的 250mL三角瓶中,在旋涡振荡器上振荡1min后置于冰水浴中1min.如此重复3次.静置 5min,吸取 5mL上述土壤悬浮液置于装有45mL灭菌生理盐水(0.85%)的150mL三角瓶中,摇匀后吸取上述土壤浸提液加入到 Biolog ECO 板,每孔 100μL,分别于 0, 24, 48, 72, 96, 120和144h在Biolog微孔板读数仪上测定其吸光值,测定波长为 590nm.土壤微生物代谢剖面的表达采用每孔的平均吸光度值(AWCD),计算式为AWCD值=[∑(C-R)]/95,其中C是所测定的95孔中每孔反应的吸光值,R是对照孔的吸光值.
土壤中 PAEs的预处理采用索氏抽提方法,参考美国环保署(US EPA) 3540方法,气相色谱-质谱联用(GC-MS)检测方法参考US EPA 8270C方法.具体步骤见文献[8-9].定量分析用 PAEs(包括DBP和DEHP等6种化合物)、内标化合物(菲-d10、芘-d10)和代用品标准物质(苯胺-d5)均为色谱纯标准物,购自美国 ULTRA Scientific,Inc., North Kingstown; RI..采用标样的0, 2.5, 5.0,7.5, 10.0μg/mL为工作曲线,内标法定量.苯胺-d5的回收率为82.69%~90.25%,DBP和DEHP的检测限为0.582μg/L和0.491μg/L,空白试验中未检测出试验化合物.
1.4 数据处理方法
实验数据采用 Duncan进行多重比较,所用软件为SAS 9.1.
2 结果与分析
2.1 2种基因型菜心不同生长期根际微生物生物量碳的动态变化特征
Bootstrap 是由 Twitter 的 Mark Otto 和 Jacob Thornton开发的。Bootstrap 是 2011 年8月在 GitHub 上发布的开源产品。Bootstrap基于 HTML、CSS、JAVASCRIPT, 用于快速开发 Web 应用程序和网站的前端框架。Bootstrap的响应式 CSS 能够自适应于台式机、平板电脑和手机。
未种植物处理、油青60处理和特青60处理根际土壤 Cmic分别介于 47.49~90.82mg/kg、84.45~213.30mg/kg 和 98.33~229.85mg/kg.各PAEs污染水平下2种基因型菜心不同生长期根际土壤Cmic的动态变化特征见图1.
各PAEs污染水平下, 2种基因型菜心不同生长期根际土壤Cmic均显著高于未种植物处理,其中油青 60根际土壤 Cmic是未种植物处理的1.2~2.6倍,特青 60根际土壤 Cmic是未种植物处理的1.5~3.4倍. 2种基因型菜心相比,除T3处理的开花期以外,特青60根际土壤Cmic均大于油青60,前者是后者的 1.1~1.7倍,绝大部分处理间差异显著.随着盆栽时间的增加,未种植物处理土壤Cmic值呈现增加的趋势,其中第 52d显著高于第15d和第25d; 2种基因型菜心根际土壤Cmic整体上呈先增加后下降(特青60天菜心T1处理除外)的趋势,且部分处理间差异显著.

对于未种植物处理, PAEs对 Cmic的影响与土壤PAEs添加浓度和取样时间有关.第15d时,试验所设置的3个PAEs添加浓度对土壤Cmic均具有一定的促进作用,50mg/kg(T2)处理比对照处理(T0)提高了46%,差异显著;第25d时,试验所设置的3个PAEs添加浓度处理土壤Cmic值接近;第39d和52d时,土壤Cmic随着土壤中PAEs添加浓度的增加而升高, T1,T2,T3比 T0分别提高了15%, 27%, 44% (第 39d)和 18%, 34%, 39% (第52d),差异显著(第39d的T1除外).
对于油青60,当土壤PAEs添加浓度不超过50mg/kg (T2)时,幼苗期和叶片生长期根际土壤Cmic随着 PAEs添加浓度的增加而增加,T2比T0分别提高了14% (幼苗期)和42% (叶片生长期),其中叶片生长期差异显著;当土壤 PAEs添加浓度达到100mg/kg (T3)时,根际土壤Cmic则受到抑制,与T0相比,幼苗期和叶片生期分别下降9%和12%,但差异不显著.随着盆栽时间的增加,菜苔形成期根际土壤Cmic则随着PAEs添加浓度的增加而显著增加, T1, T2, T3比 T0分别提高了39%,75%和99%.开花期根际土壤Cmic变化复杂,T1和 T3处理比 T0分别提高了 14%和 30%,差异显著,而T2则与T0接近.
对于特青60,与油青 60不同,本试验所设置的3个PAEs添加浓度在菜心生长幼苗期和叶片生长期并未对根际土壤 Cmic产生抑制作用,其中T2比T0分别提高了47% (幼苗期)和49% (叶片生长期),差异显著.菜苔形成期根际土壤 Cmic变化与油青60相似,随着PAEs添加浓度的增加而增加, T2, T3比T0分别提高了25%和39%.开花期根际土壤Cmic与前 3个生长期和油青 60不同,呈现 T3<T0<T2<T1,差异显著,这可能与特青 60 根际土壤中微生物生长周期特性有关.
2.2 2种基因型菜心不同生长期根际微生物生物量氮的动态变化特征
未种植物处理、油青60处理和特青60处理根际土壤微生物生物量氮(Nmic)分别介于 3.67~7.39mg/kg、9.00~25.86mg/kg 和 12.69~34.36mg/kg.各PAEs污染水平下2种基因型菜心不同生长期根际土壤Nmic的动态变化特征见图2.

对于不种植物处理,与 Cmic相似, PAEs对Nmic的影响与土壤PAEs添加浓度和取样时间有关.当土壤中PAEs添加浓度不超过50mg/kg (T2)时, Nmic在第15d和25d受到一定程度的促进作用,其中第 15d T2显著高于 T0,提高了 53%.而当土壤中PAEs添加浓度达到100mg/kg时, Nmic在第15d和25d受到一定程度的抑制作用,但差异不显著.随着盆栽时间的增加, PAEs高污染水平对土壤中Nmic的抑制作用消失, Nmic在第39d和52d则随着土壤 PAEs添加浓度的增加而增加,但差异也不显著.
2种基因型菜心不同生长期根际土壤中Nmic的变化特征与 Cmic相似.对于幼苗期和叶片生长期,当土壤PAEs添加浓度不超过50mg/kg时,油青60根际土壤Nmic随PAEs添加浓度的增加而增加,T2显著高于T0,分别提高了24% (幼苗期)和56%(叶片生长期);而特青60根际土壤Nmic随PAEs浓度的增加而显著增加,幼苗期T1和T2比T0分别提高了17%和85%,叶片生长期分别提高了13% (T1)和86% (T2).当土壤PAEs添加浓度达到100mg/kg时,油青60根际土壤Nmic受到一定程度的抵制作用,其中幼苗期比T0下降了27%,差异显著;而特青60幼苗期和叶片生长期根际土壤Nmic分别比T0提高了 30%和 50%,差异显著.随着盆栽时间的增加,高浓度 PAEs (100mg/kg)对油青 60根际土壤Nmic的抑制作用消失,菜苔形成期2种基因型菜心根际土壤Nmic均随PAEs浓度的增加而增加,大部分处理间差异显著.开花期各PAEs污染2种基因型菜心根际土壤Nmic变化不大,除特青60T1显著高于T0外,其他处理间差异均不显著.
2.3 2种基因型菜心不同生长期根际土壤微生物功能多样性的动态变化特征
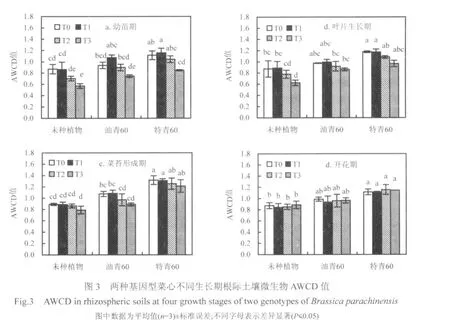
平均颜色变化率(AWCD)作为微生物总体活性的一个重要指标,其值的大小代表了不同处理土壤微生物对底物碳源利用能力上的差异.本实验采用Biolog微平板培养96h的数据来进行微生物总体代谢活性分析.未种植物处理、油青60处理和特青60处理根际土壤AWCD分别介于 0.571~0.891、0.744~1.075 和 0.847~1.318.各PAEs污染水平下2种基因型菜心不同生长期根际土壤AWCD的动态变化特征见图3.
与土壤 Cmic和 Nmic相似,各 PAEs污染水平下,不同生长期 2种基因型菜心根际土壤微生物总体代谢活性均大于未种植物处理,其中油青60根际土壤AWCD是未种植物处理的1.1~1.4倍,差异不显著;特青60根际土壤AWCD是未种植物处理的1.3~1.6倍,差异显著. 2种基因型菜心相比,特青60根际土壤微生物总体代谢活性高于油青60,前者AWCD值是后者的1.1~1.4倍,其中菜苔形成期差异显著.
对于未种植物处理,当 PAEs添加浓度为20mg/kg时,土壤AWCD值与T0接近;当PAEs添加浓度达到50mg/kg时,第15d和25d AWCD值下降,其中 T2(50mg/kg)比 T0分别下降 19%(第15d)和11%(第25d),但差异不显著; T3(100mg/kg)比T0分别下降34%(第15d)和29%(第25d),差异显著.随着培养时间的增加, PAEs对土壤微生物总体代谢活性的影响减小,第39d和52d各污染水平间 AWCD 值接近,无显著差异.这与前人研究结果相符[3,17],可能与土壤中PAEs对微生物的毒性随着时间的增加而减弱有关.
PAEs污染对2种基因型菜心根际土壤微生物总体代谢活性的影响与Cmic和Nmic不同.与T0相比, PAEs低污染浓度(20mg/kg,T1)在幼苗期对2种基因型菜心根际土壤AWCD均具有一定的促进作用,但差异不显著;而当 PAEs添加浓度达到50mg/kg时,幼苗期和叶片生长期对2种基因型菜心根际土壤AWCD均具有一定的抑制作用,其中特青60幼苗期T3比T0下降了24%,差异显著.随着盆栽时间的增加,叶片生长期 PAEs高污染浓度(≥50mg/kg,T1)对特青60根际土壤AWCD的影响逐渐减弱,各污染水平下的AWCD值接近;而对油青60根际土壤 AWCD仍具有一定的抑制,T2和T3比T0分别下降了10%和17%,但差异不显著.开花期 PAEs对根际土壤微生物总体代谢活性的抑制和促进作用减弱,各 PAEs污染水平下的AWCD值接近.
3 讨论
有关PAE污染对土壤微生物影响的研究表明,当土壤DBP浓度小于10mg/kg, DEHP浓度小于20mg/kg时,对土壤Cmic没有明显影响;而当浓度达到50mg/kg时,对土壤Cmic表现出抑制效应,且抑制作用随处理浓度的增加而加强[3].当DEHP浓度达100mg/kg时,显著降低了土壤微生物代谢活性[16].本文中,PAEs对 Cmic和当土壤PAEs添加浓度不超过 50mg/kg时,对土壤 Cmic和 Nmic具有一定的刺激作用.当土壤 PAEs添加浓度达到100mg/kg时,对土壤Cmic和Nmic的刺激和抑制作用在幼苗期和叶片生长期并不明显,但在菜苔形成期和开花期表现为显著的刺激作用.这与文献[3]的报道不同.究其原因可能有:其一,试验条件不同.与文献[3]的研究所用室内恒温箱相比,本试验是在通风的网室中进行,可能更有利于PAEs的非生物性损失和生物降解,从而导致2个试验中土壤PAEs的实际生物有效性可能不同;其二,取样时间不同.本研究的取样时间为添加污染物后的第20, 30, 44, 57d,较文献[3]的取样时间要迟.而一般来说,培育时间越长,污染物在土壤中的浓度和生物有效性越低,从而对土壤微生物的生态毒性作用越弱.如,百菌清处理前 3d对土壤微生物活性产生显著的抑制作用,但7d后逐渐恢复[17].恩诺沙星对土壤微生物功能多样性的抑制作用也只维持9d[18].农药对土壤微生物量的影响往往在1个月之内就恢复到原来的水平[19].本实验中,当土壤PAEs添加浓度为20mg/kg时,土壤微生物代谢活性与T0接近,而当土壤PAEs添加浓度达到50mg/kg时,对土壤微生物代谢活性具有一定的抑制作用,抑制作用的强度随 PAEs浓度的增加而增加,当土壤 PAEs添加浓度达到100mg/kg时作用显著.这与文献[3]和[17]所报道的100mg/kg一致.
种植 2种基因型菜心根际土壤 Cmic、Nmic以及AWCD均高于未种植物处理,与PAEs污染下种植绿豆处理根际土壤中微生物代谢活性高于未种植绿豆处理结果一致[16],这可能与植物生长过程中根系向生长介质分泌有机酸、糖类物质和氨基酸等物质,为土壤微生物的生长提供有利条件[16,20].不同植物甚至是同一植物不同基因型间土壤微生物数量和群落结构也存在一定的差异.如,超积累生态型东南景天根际土壤的微生物生物量碳氮以及微生物对碳源的利用速率和强度等均显著高于非超积累型东南景天的根际土壤[13].本实验中,特青60根际土壤Cmic、Nmic以及AWCD均高于油青 60 (T3处理的开花期除外),前者对Cmic、Nmic以及AWCD的促进作用均达到显著程度,而后者仅显著提高了根际土壤中Cmic和Nmic,对AWCD的促进作用不显著.一般来说,土壤中微生物生物量越大,代谢功能多样性越强,越有利于有机污染物的降解,从而降低植物对有机污染物的吸收和有机污染物在土壤中的残留.2种基因型菜心茎叶和根系中 PAEs含量与Cmic、Nmic和AWCD之间的相关性均未达到显著程度,说明 2种基因型菜心茎叶和根系中 PAEs含量可能与其他因素,如, 2种基因型菜心根系形态和生理特性[21]、根系解剖结构[22]及根际土壤中 PAEs的生物有效性[23]相关.根系分泌物不仅为根际微生物生长提供所需的能源,影响根际微生物的数量和种群结构[24-25].根际分泌物还会直接影响根际土壤中有机污染物的生物有效性[26],所以 2种基因型菜心根系分泌物的差异,及其对根际微生物群落结构和功能的特异性影响还有待于深入研究.
4 结论
4.1 两种基因型菜心根际土壤 Cmic分别为84.45~213.30mg/kg 和 98.33~229.85mg/kg,Nmic分别为 9.00~25.86mg/kg 和 12.69~34.36mg/kg,AWCD 值分别为 0.744~1.075 和 0.847~1.318,均为特青60大于油青60,甚至达到显著程度.
4.2 两种基因型菜心根际土壤 PAEs残留量主要与AWCD有关,但茎叶和根系中PAEs含量与Cmic、Nmic和 AWCD相关性不显著,说明这些因素不是导致2种基因型菜心茎叶和根系中PAEs含量显著差异的主要原因.
[1] 蔡全英,莫测辉,李云辉,等.广州、深圳地区蔬菜生产基地土壤中邻苯二甲酸酯(PAEs)研究 [J]. 生态学报, 2005,25(2):283-288.
[2] 杨国义,张天彬,高淑涛,等.广东省典型区域农业土壤中邻苯二甲酸酯的分布特征 [J]. 应用生态学报, 2007,18(10):2308-2312.
[3] 高 军,陈伯清.酞酸酯污染土壤微生物效应与过氧化氢酶活性的变化特征 [J]. 水土保持学报, 2008,22(6):166-169.
[4] 蔡玉祺,汤国才,王珊龄,等.邻苯二甲酸酯对蔬菜幼苗的影响[J]. 农业环境保护, 1994,13(4):163-166.
[5] 吴志辉,杨宇峰,聂湘平.酞酸酯对龙须菜的生态毒理研究 [J].海洋科学, 2006,30(6):46-50.
[6] Yin R, Lin X G, Wang S G, et al. Effect of DNBP/DEHP in vegetable planted soil on the quality of capsicum fuit [J].Chemosphere, 2003,50:801-805.
[7] 蔡全英,莫测辉,曾巧云,等.邻苯二甲酸酯在不同品种通菜-土壤系统中的累积效应研究 [J]. 应用生态学报, 2004,15(8):1455-1458.
[8] 曾巧云,莫测辉,蔡全英,等.邻苯二甲酸二丁酯在不同品种菜心-土壤系统的累积 [J]. 中国环境科学, 2006,26(3):333-336.
[9] 曾巧云,莫测辉,蔡全英,等.不同基因型菜心-土壤系统中邻苯二甲酸二(2-乙基已基)酯的分布特研究 [J]. 农业环境科学学报,2007,26(6):2239-2244.
[10] Staples C A, Peterson D R, Parkerton T F, et al. The environmental fate of phthalate esters: A literature review [J].Chemosphere, 1997,35:667-749.
[11] Arab H, Vlich V, Sikora R A. The use of phospholipids fatty acids(PLFA) in the determination of rhizosphere specific microbial communities of two wheat cultivars [J]. Plant and Soil,2001,228:291-297.
[12] Kapoor R. Root exudation and its application on rhizosphere mycoflora [J]. Advances in microbial biotechnology, 1999,351-362.
[13] 龙新宪,刘洪彦,戴 军,等.两种生态型东南景天根际与非根际土壤微生物特征的差异性 [J]. 土壤学报, 2009,46(3):547-552.
[14] Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C [J]. Soil Biol. Biochem.,1987,19:703-707.
[15] Chen J, Zhuang X, Xie H, et al. Associated impact of inorganic fertilizers and pesticides on microbial communities in soils [J].World J Microbiol and Biotechnol, 2007,23:23-29.
[16] 秦 华,林先贵,陈瑞蕊,等. DEHP对土壤脱氢酶活性及微生物功能多样性的影响 [J]. 土壤学报, 2005,42(5):829-834.
[17] 郝乙杰,向月琴,方 华,等.百菌清在土壤中的降解及对土壤微生物多样性的影响 [J] 农业环境科学学报, 2007,26(5):1672-1676.
[18] 王加龙,刘坚真,陈杖榴,等.恩诺沙星残留对土壤微生物功能的影响 [J]. 生态学报, 2005,25(2):279-282.
[19] Martinez M V, Salmeron V, Gonzalez Z J. Effect of simazine of the biological activity of Azotobacter chroococcum [J]. Soil Science, 1996,151:459-467.
[20] 王 江,张崇邦,常 杰,等.五节芒对重金属污染土壤微生物生物量和呼吸的影响 [J]. 应用生态学报, 2008,19(8):1835-1840.
[21] 曾巧云,莫测辉,蔡全英,等.两种基因型菜心根系形态和生理特性差异对其吸收累积DEHP影响的初步研究 [J]. 环境科学学报, 2010,30(6):1280-1285.
[22] 曾巧云,莫测辉,蔡全英,等.2种基因型菜心根系解剖结构差异及其对DEHP吸收累积的影响 [J]. 环境科学, 2010,31(8):58-61.
[23] 曾巧云,莫测辉,蔡全英,等.2种基因型菜心根系分泌物对水稻土中 DEHP解吸效应的初步研究 [J]. 农业环境科学学报,2010,29(8):1466-1469.
[24] Bakken L R. Microbial growth and immobilization/mineralization of N in the Rhizosphere [J]. Symbiosis, 1990,9:37-411.
[25] 申建波,张福锁,毛达如.根际微生态系统中的碳循环 [J]. 植物营养与肥料学报, 2001,7(2):232-240.
The microbial characteristics and differences in rhizospheric soils contaminated with PAEs grown high/low PAEs accumulation genotypes of Brassica parachinensis.
ZENG Qiao-Yun1, MO Ce-Hui2*, CAI Quan-Ying1(1. Key Laboratory of Ecological Agriculture of Ministry of Agriculture of the People1’s Republic of China, College of Resources and Environment, South China Agricultural University, Guangzhou 510642, China;2.Department of Environmental Engineering, Jinan University, Guangzhou 510632, China). China Environmental Science, 2011,31(3):466~473
Two genotypic cultivars of Brassica parachinensis, Youqing-60 (high PAEs-accumulation) and Teqing-60 (low PAEs-accumulation), were cultivated in the soils spiked with three levels (20, 50 and 100mg/kg) of dibutyl phthalate and di(2-ethylhexyl) phthalate to investigate the microbial biomasses and metabolic activities in rhizospheric soils at four growth stages of Brassica parachinensis. The results showed that the microbial biomass C (Cmic) in rhizospheric soils of Youqing-60 and Teqing-60 ranged from 84.45 to 213.30mg/kg and 98.33 to 229.85mg/kg, and their microbial biomass N(Nmic) varied from 9.00to 25.86mg/kg and 12.69 to 34.36mg/kg, respectively, which were significantly higher than those in soils without plant. Their average well color development (AWCD) values ranged from 0.744 to 1.075 and 0.847 to 1.318, respectively, which of Teqing-60 were significantly higher than those in soils without plant. Moreover, Cmic, Nmicand AWCD in soils grown cultivar Teqing-60 were higher than those of Youqing-60 (except for treatment of T3at flowering stage), and significant differences were observed between some treatments. PAEs concentrations in shoots and roots of two genotypes of Brassica parachinensis were not significantly negatively correlated with Cmic, Nmicand AWCD values, indicating that PAEs concentrations in shoots and roots of two genotypes of Brassica parachinensis may be related to other factors.
phthalic acid esters;Brassica parachinensis;soil;microbial biomass;microbial functional diversity
X503:23
A
1000-6923(2011)03-0466-08
2010-07-19
国家自然科学基金资助项目(30600372,30471007);广东省科技计划项目(2010A020507001-73,2010B020311006);中央高校基本科研业务费专项资金(21610410,21609709);广东省高校高层次人才项目
* 责任作者, 教授, tchmo@jnu.edu.cn
曾巧云(1974-),女,湖南益阳人,讲师,硕士,主要从事环境有机污染与控制技术研究工作.发表论文30余篇.
猜你喜欢
源流(2022年11期)2023-01-29 11:26:20
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
中国蔬菜(2016年8期)2017-01-15 14:23:38
饮食与健康·下旬刊(2016年9期)2016-05-14 17:26:31
西湖(2016年9期)2016-05-14 08:44:03
现代检验医学杂志(2015年6期)2015-02-06 01:44:02
实验动物与比较医学(2014年5期)2014-02-28 14:53:10
中国糖料(2013年1期)2013-01-22 12:28:23
医学综述(2011年18期)2011-12-10 01:09:01
